










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.64 no.2 Medellín July/Dec. 2011
Selección de Cepas de Hongos Entomopatógenos para el Manejo de Anastrepha obliqua (Macquart, 1835) (Diptera: Tephritidae) en Colombia
Selection of Strains of Entomopathogenic Fungi for Management of Anastrepha obliqua (Macquart, 1835) (Diptera: Tephritidae) in Colombia
Armando Osorio-Fajardo1 y Nelson A. Canal2
1 Investigador Asociado. Universidad del Tolima - Grupo de Investigación en Moscas de las Frutas. A.A. 546, Ibagué, Colombia. <osorio.fajardo@hotmail.com>
2 Profesor Asociado. Universidad del Tolima - Facultad de Ingeniería Agronómica. Barrio Altos de Santa Elena, A.A. 546, Ibagué, Colombia. <nacanal@ut.edu.co>
Recibido: Marzo 08 de 2011; aceptado: Octubre 31 de 2011.
Resumen: Se evaluaron 15 cepas de los hongos entomopatógenos Beauveria bassiana y Metarhizium anisopliae sobre adultos de un día de edad de la mosca de la fruta Anastrepha obliqua. El trabajo se realizó con el fin de seleccionar las cepas más virulentas al insecto y estudiar el efecto sobre los adultos jóvenes cuando el hongo era aplicado antes de la emergencia. Mediante un screening con una concentración de 1x107 conidias/mL se seleccionaron las tres cepas más virulentas, siendo dos de ellas de Beauveria y una de Metarhizium, las cuales causaron mortalidades del 77%, 71% y 66%. Valores de CL50 de 2,38x106, 1,81x106 y 9,94x106 conidias/mL, respectivamente, fueron determinados para cada una de estas cepas y un TL50 respectivo de 48,12; 56 y 42,75 horas. No se encontraron diferencias significativas en la mortalidad entre hembras y machos. La aspersión de la CL90 de las cepas seleccionadas sobre el medio de pupación de la mosca de la fruta produjo 34-48% de mortalidad durante las 120 horas de evaluación. Los hongos entomopatógenos pueden ser utilizados fácilmente para el control biológico de A. obliqua aplicándolos de manera dirigida a los adultos jóvenes bajo la copa de los árboles, en programas de manejo integrado de plagas.
Palabras clave: Beauveria bassiana, control biológico, moscas de la fruta, Metarhizium anisopliae.
Abstract: Fifteen strains from entomopathogenic Beauveria bassiana and Metarhizium anisopliae fungi were evaluated on one day-old adults of Anastrepha obliqua fruit fly. Tested were carried out for selecting the most virulent strains and the effectiveness of their use on young adult when the entomopathogen were applied before emergence were studied too. A screening with a 1x107 conidia/mL concentration was used for selecting the three most pathogenic isolates, two from Beauveria and one from Metarhizium, having 77, 71 and 66% mortality. The LC50 for these isolates were 2.38x106, 1.81x106 and 9.94x106 conidia/mL, respectively, and a respective LT50 were 48.12, 56 and 42.75 hours. No significant differences were found between females and males mortality. LC90 spraying of selected strains on fly pupation medium led to 34-48% mortality at 120 hours. Entomopathogenic fungi could be used for A. obliqua biological control and also it can be used easily spraying towards young adults under tree canopies in the integrated pest management programs.
Key words: Beauveria bassiana, biological control, fruit flies, Metarhizium anisopliae.
Las verdaderas moscas de las frutas pertenecen a la familia Tephritidae, cuyos ataques a las especies frutales ocasiona un importante impacto económico, es así como en Colombia las moscas del género Anastrepha son las causantes del mayor daño a las frutas y la mosca Anastrepha obliqua (Macquart) es el principal problema del mango en muchas áreas del neotrópico; sin embargo, también se ha reportado en un gran número de plantas hospederas, principalmente especies de Anacardiaceae (White y Elson, 1992).
Tradicionalmente el manejo de las moscas de la fruta se ha realizado utilizando aspersiones de cebos tóxicos y medidas culturales; estas labores han sido poco efectivas y han acarreado problemas de contaminación (Aluja, 1994). Existen otras alternativas de manejo más amigables con el medio ambiente, una de ellas es la Técnica del Insecto Estéril (TIE), cuyos resultados han sido muy exitosos (Enkerlin, 2005). También se han referido programas de control biológico exitosos, bien sea utilizando solo enemigos naturales (Montoya y Liedo, 2000; Rendon et al., 2006) o en programas integrados a la TIE (Montoya y Cancino, 2004), y recientemente se ha planteado la posibilidad de utilizar control microbiológico (Roessler, 1989; Quesada et al., 2006; Almeida et al., 2007).
Existe referencia de la susceptibilidad de varias especies de mosca de las frutas a nemátodos entomopatógenos (Toledo et al., 2006; Almeida et al., 2007), Bacillus thuringensis Berliner (Robacker et al., 1996; Toledo et al., 1999), y a hongos entomopatógenos. Estos últimos se han evaluado sobre estados inmaduros de la mosca (Ekesi et al., 2002; Destéfano et al., 2005; Almeida et al., 2007) y sobre adultos, en dispositivos de impregnación, mediante aplicación tópica o por inmersión (Dimbi et al., 2003; Quesada et al., 2006; Ali et al., 2008) en pruebas de laboratorio o campo. Dimbi et al. (2003), evaluaron, en laboratorio, 14 aislamientos de hongos entomopatógenos sobre tres especies de Ceratitis usando la metodología del residuo y encontraron mortalidades entre 7 y 100%. Destéfano et al. (2005) evaluaron el efecto de un aislado de Metharizium anisopliae (Metschnikoff) incorporado al suelo sobre larvas, pupas y adultos de Anastrepha fraterculus (Wiedemann), alcanzando mortalidades del 86% referida a la emergencia de adultos en los tratamientos. Ekesi et al. (2002) evaluaron 15 aislamientos de hongos entomopatógenos, incorporados en arena, sobre larvas de tercer estadío de dos especies de Ceratitis, encontrando que todos causaban mortalidad significativa. Quesada et al. (2006), en laboratorio, probaron 10 cepas de hongos sobre pupas y adultos de Ceratitis capitata (Wiedemann), encontrando mortalidades entre 70 y 100% para adultos y entre 52,5 y 70% para puparios. Almeida et al. (2007) evaluaron diez aislamientos de Beauveria bassiana (Balsamo) Vuillemin y M. anisopliae sobre prepupas de C. capitata, en pruebas de laboratorio e invernadero, encontrando reducciones en la mortalidad de adultos de hasta el 80% en condiciones de laboratorio y 66,6% en invernadero. Ali et al. (2008) evaluaron en laboratorio, utilizando la metodología de inmersión, la mortalidad causada por Beauveria y Paecilomyces sobre adultos de C. capitata, encontrando mortalidades hasta del 100% con concentraciones de 3x107 conidias/mL de Beauveria y hasta 70% con la misma concentración de Paecilomyces.
En Colombia el manejo de las moscas de la fruta se limita al uso de cebos tóxicos, aplicación de insecticidas químicos, medidas culturales y trampeo, además, a pesar de la importancia de A. obliqua para Latinoamérica, no existen trabajos referentes a su susceptibilidad a los hongos entomopatógenos. Considerando que la eficiencia de las cepas de hongos entomopatógenos pueden variar en ambientes diferentes, en este trabajo se evaluó la patogenicidad de 15 cepas nativas de los hongos entomopatógenos M. anisopliae y B. bassiana sobre adultos de A. obliqua, con el fin de contribuir con herramientas biológicas para programas de manejo integrado de esta especie en Colombia y toda la región de incidencia de la plaga.
MATERIALES Y MÉTODOS
Insectos. Puparios del insecto fueron obtenidos del pie de cría del Laboratorio de Diagnóstico Vegetal del Instituto Colombiano Agropecuario ICA (Ibagué -; Tolima, Colombia), los cuales han sido criados desde el año 1993 en dieta artificial. Las pruebas se realizaron en el laboratorio de Entomología de la Universidad del Tolima. Luego que los adultos emergieron se mantuvieron en jaulas de 30x30x30 cm, a temperatura ambiente. Estos adultos fueron alimentados a voluntad con la dieta artificial descrita por Núñez y Guzmán (1999).
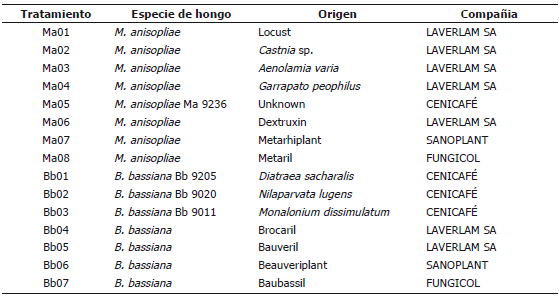
Hongos entomopatógenos. Los hongos entomopatógenos fueron obtenidos de ceparios de los laboratorios LAVERLAM S.A. (Cali - Valle del Cauca, Colombia), del Centro Nacional de Investigaciones de Café CENICAFE (Chinchiná - Caldas, Colombia) y de productos comerciales. Todas las cepas se repicaron en medio Papa-Destroxa-Agar, PDA, y se mantuvieron a temperatura ambiente para la recolección del inóculo necesario. Se obtuvieron 15 aislamientos, siete de B. bassiana y ocho de M. anisopliae, siete de ellos provenientes de productos comerciales y ocho de cepas almacenadas, pero no existía disponibilidad de cepas provenientes de tefritídeos o al menos de dípteros (Tabla 1).
Tabla 1. Origen de las cepas de Metharizium anisopliae y Beauveria bassiana evaluadas en adultos de un día de edad de Anastrepha obliqua.
Preparación del inóculo. Para la producción del inóculo en cada una de las etapas del experimento, se realizaron subcultivos de las cepas en cajas de Petri con PDA selladas con Parafilm y mantenidas a temperatura ambiente por siete días. Las concentraciones del hongo fueron preparadas por raspado de los conidios de las cajas de Petri en una suspensión de agua destilada estéril + Tween 20 al 0,1%.
Se realizaron tres grupos de experimentos, los cuales serán denominados etapa I, etapa II y etapa III. La etapa I fue un relevamiento para escoger las tres cepas más virulentas; la etapa II consistió en la determinación de la CL50, CL90 y TL50 de las cepas seleccionadas anteriormente y la etapa III fue un ensayo de mortalidad por aspersión de la CL90 sobre el sustrato de pupación. La concentración del inóculo para la etapa I fue ajustada por dilución a 1x107 conidias/mL. En la etapa II se usaron las concentraciones de 2x107, 1x107, 5x106 y 1x105 conidias/mL y en la etapa III se utilizaron las concentraciones equivalentes a las CL90 de las cepas seleccionadas, que fueron 7,4x109, 1,4x109 y 7,1x109 conidias/mL. En todos los ensayos se utilizaron tres tratamientos como testigo: tratamiento con agua destilada + Tween 20 (polisorbato de sodio) al 0,1%, tratamiento con agua destilada y testigo absoluto; sin embargo, como no se encontraron diferencias entre los tratamientos testigo, se muestran los resultados usando solo el testigo absoluto, el cual fue considerado como un tratamiento más.
Para los experimentos de las etapas I y II se utilizaron adultos de la mosca de un día de edad. Cada tratamiento se replicó cinco veces y cada réplica consistía de 20 adultos de la mosca (10 Hembras y 10 Machos) en una jaula tipo EUGO (González et al., 1997a) con agua y alimento. En las pruebas de selección de cepas era necesario garantizar un buen contacto de las moscas con los entomopatógenos, por lo tanto, cada individuo fue sumergido en el tratamiento respectivo por cinco segundos, secado ligeramente en papel toalla y transferidos a las jaulas, como fue realizado también por Ali et al. (2008). Se realizó conteo de mortalidad cada 24 horas por cinco días. Para la etapa III se utilizó un aerógrafo para asperjar 5 mL de la CL90 de los tratamientos seleccionados, o de los tratamientos testigos (agua+Tween 20) y agua, sobre el medio de pupación (vermiculita), conteniendo 100 puparios de A. obliqua próximos a la emergencia de los adultos. Luego de la aspersión de los tratamientos, se dejaron emerger los adultos en jaulas metálicas de 30x30x30 cm. Una vez emergidas las moscas adultas se trasladaron a las jaulas tipo EUGO, hasta completar 20 individuos por réplica. Cada tratamiento fue repetido cinco veces, realizando el registro de mortalidad de las moscas de la misma forma que en las etapas anteriores.
En cada una de las pruebas, los adultos muertos eran retirados diariamente de las jaulas y trasladados a cámara húmeda, a temperatura de 25 ºC para la revisión de la esporulación.
Análisis estadístico. Se analizaron los datos de mortalidad usando el análisis de varianza (ANAVA) y el test DMS para determinar las diferencias de patogenicidad por comparación de las medias. La concentración letal media CL50, la CL90 y el tiempo letal medio TL50, fueron estimados por medio del análisis de Probit con los porcentajes de mortalidad acumulada. El método estadístico fue totalmente al azar. La unidad experimental del bioensayo fue la jaula tipo EUGO con 20 individuos de A. obliqua de un día de edad. Las lecturas de mortalidad fueron transformadas inicialmente con el porcentaje de esporulación del tratamiento y luego como
RESULTADOS
Etapa I -; Selección de cepas patogénicas. En esta etapa todas las cepas evaluadas mostraron patogenicidad promisoria sobre los adultos de un día de edad de A. obliqua. El total de la mortalidad estuvo entre 18 y 92% a las 120 h de evaluación, siendo que dos tratamientos de B. bassiana (Bb05, Bb03) y uno de M. anisopliae (Ma06) causaron una mortalidad superior al 80% a las 96 h y los tratamientos Bb05 y Ma06 causaron más del 90% de mortalidad a las 120 h (Tabla 2). La mortalidad en el testigo fue de 3% a las 120 h. Las menores mortalidades se encontraron para los tratamientos Ma02, Ma04 y Bb04 (Tabla 2). Todos los tratamientos realizados con hongos presentaron esporulación del entomopatógeno, la cual estuvo entre 70 y 92% cinco días después de la muerte de los insectos; sin embargo, no se encontró diferencia estadística entre la esporulación de los diferentes tratamientos.
Tabla 2. Virulencia de cepas de Metharizium anisopliae y Beauveria bassiana en adultos de un día de edad de Anastrepha obliqua. Porcentajes de mortalidad en el screening durante 120 horas de evaluación.
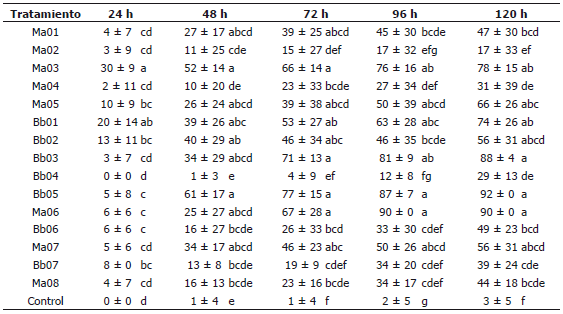
Los porcentajes de mortalidad de los adultos de A. obliqua de un día de edad, presentaron diferencias significativas entre tratamientos en cada uno de los tiempos de evaluación y los coeficientes de variación, disminuyeron a medida que avanzó el tiempo de lectura debido a la completa expresión de la infección resultante (24 h: F= 3,1152, P<0,00059, CV= 16,259%; 48 h: F= 3,2468, P<0,00040, CV= 20,646%; 72 h: F= 5,2246, P<0,00001, CV= 18,61%; 96 h: F= 6,0282, P<0,00001, CV= 16,989%; 120 h: F= 6,8030; P<0,00001, CV= 14,939%). Se observaron diferencias entre tratamientos de la misma especie de hongo (Tabla 2).
Se realizó un análisis de mortalidad diaria de la mosca con los tratamientos a base de M. anisopliae y B. bassiana, encontrándose diferencias significativas entre las evaluaciones realizadas hasta las 72 h, pero siendo estadísticamente igual las mortalidades entre las 72, 96 y 120 h (F=18,1079, P<0,00001, CV= 14,638%) (Figura 1). Con este análisis se decidió utilizar la mortalidad a las 72 h, como el parámetro de tiempo de selección para la siguiente prueba, pues es el menor tiempo con máxima mortalidad. Se seleccionaron para continuar el estudio los tratamientos Bb05, Bb03 y Ma03, los cuales presentaron los mayores porcentajes de mortalidad a las 72 h.
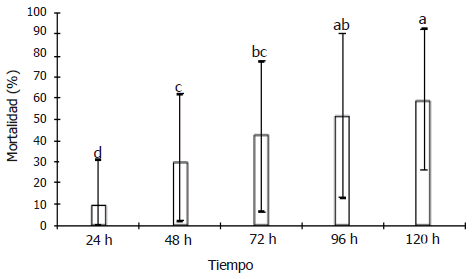
Figura 1. Mortalidad diaria de Anastrepha obliqua tratada con diferentes aislamientos de Metharizium anisopliae y Beauveria bassiana. Tratamientos con diferentes letras difieren estadísticamente.
La hembra de la mosca es la causante del daño directo a la fruta, la mayor susceptibilidad de éstas permitiría un control más eficiente de la plaga; sin embargo, son pocos los trabajos referidos al tema. Por lo tanto, para determinar alguna susceptibilidad ligada al sexo de A. obliqua a M. anisopliae y/o B. bassiana, se analizaron los porcentajes de mortalidad de los dos sexos a las 72 h, para cada uno de los tratamientos seleccionados. Este análisis no mostró diferencias significativas de mortalidad ligada al sexo (Bb05: F=0,0026, P<0,95957, CV=6,320%; Bb03: F=0,0006, P<0,97877, CV=7,057%; Ma03: F=0,0242, P<0,87450, CV=5,750%).
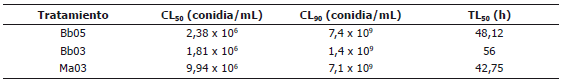
Etapa II - Determinación de la CL50, CL90 y TL50. La mortalidad de A. obliqua fue ascendente en el tiempo y mayor que en el testigo (Figura 2). Según los resultados del análisis de Probit, se requiere menor cantidad del inóculo con los tratamientos Bb03 y Bb05 de B. bassiana (1,81x106 conidias/mL y 2,38x106 conidias/mL, respectivamente); sin embargo, con el tratamiento Ma03 de M. anisopliae se requiere menor tiempo (42,75 h) para causar el 50% de mortalidad a las moscas (Tabla 3).
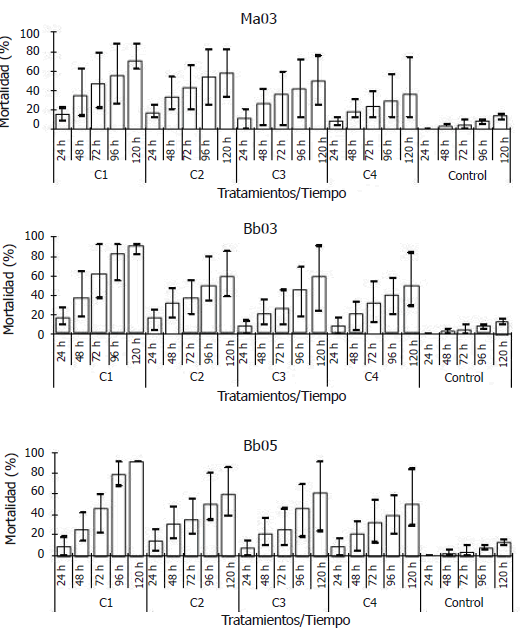
Figura 2. Porcentaje medio (barras), máximo y mínimo (lineas), de la mortalidad de adultos de un día de edad de Anastrepha obliqua, tratados con cuatro concentraciones de tres aislamientos de hongos entomopatógenos. Concentraciones: C1, 2x107 conidia/ml; C2, 1x107 conidia/ml; C3 5x106 conidia/ml; C4 1x105 conidia/ml.
Tabla 3. Valores de CL50, CL90 y TL50 calculados para tres cepas de los hongos entomopatógenos Metharizium anisopliae y Beauveria bassiana en adultos de un día de edad de Anastrepha obliqua.
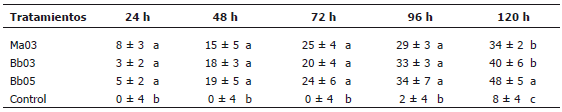
Etapa III -; Aspersión de la CL90. Este ensayo se hizo con el fin de conocer la posibilidad de aplicar hongos entomopatógenos sobre adultos poco tiempo antes de su emergencia. La mortalidad total de los tratamientos estuvo entre 47 y 63%. En cada uno de los tiempos de evaluación en esta fase se presentaron diferencias significativas entre los tratamientos a base de hongo con respecto a los tratamientos testigo (24 h: F=5,3765, P<0,00216, CV=8,997%; 48 h: F=16,2284, P<0,00001, CV=8,416%; 72 h: F=24,9536, P<0,00001, CV= 7,433%; 96 h: F=42,1433, P<0,00001, CV= 5,806%; 120 h: F=90,2464, P<0,00001, CV=3,911%) (Tabla 4). No se observó mortalidad de pupas en el experimento.
Tabla 4. Virulencia de tres cepas de Metharizium anisopliae y Beauveria bassiana en Anastrepha obliqua en aplicaciones sobre el medio de pupación. Porcentajes de mortalidad de adultos durante 120 horas de evaluación.
DISCUSIÓN
A. obliqua en Colombia es susceptible a los hongos entomopatógenos M. anisopliae y B. bassiana, de la misma forma que otros tefritídeos han mostrado susceptibilidad a los mismos (Lezama et al., 2000; Ekesi et al., 2002; Dimbi et al., 2003; Quesada et al., 2006; Almeida et al., 2007; Ali et al., 2008). La mortalidad de A. obliqua con M. anisopliae y B. bassiana se encuentran entre los rangos de mortalidad mencionados en ensayos con hongos entomopatógenos sobre moscas de la fruta. Lezama et al. (2000) refieren mortalidades superiores al 83,7% con M. anisopliae sobre Anastrepha ludens (Loew.), mientras con B. bassiana, De la Rosa et al. (2002) encontraron mortalidades del 98-100% sobre la misma especie de mosca. En adultos de C. capitata y C. rosa var. fasciventris (Karsch.) Dimbi et al. (2003) registraron mortalidades del 7 al 100% y 11,4 al 100%, con aislamientos de M. anisopliae y B. bassiana, respectivamente. Igualmente Quesada et al. (2006) encontraron mortalidades en adultos de C. capitata del 30-100% con cepas de M. anisopliae y B. bassiana después de la aplicación tópica sobre el abdomen. En los experimentos de Toledo et al. (2007) las mortalidades de A. ludens fueron superiores al 98% con dos cepas de B. bassiana y Toledo et al. (2008) refieren entre 75,2 y 100% de mortalidad de C. capitata y A. ludens con nueve cepas de B. bassiana y M. anisopliae. Quesada et al. (2008) registran entre 95 y 100% de mortalidad de poblaciones de laboratorio de C. capitata por transmisión horizontal de M. anisopliae y Ali et al. (2008) señalan mortalidades máximas de 100% de C. capitata con B. bassiana y 75% con Paecilomices. Almeida et al. (2007), luego de sumergir prepupas de C. capitata en suspensiones de conidios de B. bassiana y M. anisopliae, encontraron una reducción de la emergencia de adultos de hasta 80%.
Tres aislamientos de M. anisopliae y de B. bassiana fueron seleccionados en este trabajo como de alta virulencia sobre adultos de A. obliqua, sin embargo, los otros aislamientos evaluados muestran mortalidades que pueden ser consideradas importantes para posibles programas de manejo integrado de la mosca.
Las respuestas de mortalidad fueron diferentes entre las diferentes cepas de hongos estudiados. Diferencias entre cepas también son observadas en evaluaciones sobre C. capitata con hongos entomopatógenos (Ekesi et al., 2002; Dimbi et al., 2003; Quesada et al., 2006; Almeida et al., 2007), por este motivo se han realizado pruebas en diferentes regiones y con diferentes especies plagas, con el fin de encontrar las cepas más patogénicas que se puedan utilizar en programas locales de manejo integrado de plagas.
El criterio de selección utilizado por otros autores ha sido el de alta patogenicidad de las cepas, superiores al 80% o en rangos menores. Lezama et al. (2000) en la selección de tres aislamientos de M. anisopliae sobre larvas de último instar de A. ludens, manejaron mortalidades superiores al 83,7%. Ekesi et al. (2002) seleccionaron los aislamientos de M. anisopliae ICIPIE 18, 20, 60 y 69 con mortalidades superiores al 80%, bajo el criterio de alta patogenicidad a C. capitata y C. r. fasciventris. Igualmente Dimbi et al. (2003) seleccionaron los aislamientos que causaron más del 80% de mortalidad a C. capitata y C. r. fasciventris para ser usados contra C. cosyra (Walker.). Mientras Quesada et al. (2006) seleccionaron aislamientos que ocasionaron mortalidades entre un 62,5% a 95% sobre C. capitata.
La mortalidad ligada al sexo ha sido evaluada en respuestas de mortalidad a insecticidas químicos, donde los resultados han sido variables, González et al. (1997b) en aplicaciones tópicas y por consumo en cebo de Malation sobre A. striata Schiner, no encontraron ninguna susceptibilidad ligada al sexo; cuando los insectos fueron tratados con el insecticida y Cruz et al. (1997) en evaluaciones con el insecticida Fenthion sobre A. fraterculus, en aplicaciones tópicas y por consumo tampoco encontraron susceptibilidad ligada al sexo, sin embargo, Humeres et al. (1999) encontraron que los machos de A. fraterculus eran más tolerantes que las hembras a la exposición de Fenthion en todos los estados de desarrollo.
Los valores de CL50 para A. obliqua con los tratamientos a base de B. bassiana son menores a los referidos, mientras con M. anisopliae son mayores a los obtenidos en la mayoría de los estudios sobre moscas de la fruta. De la Rosa et al. (2002), encontraron valores de CL50 de 3,12x106 a 9,07x106 conidias/mL con B. bassiana al evaluar en larvas, pupas y adultos de A. ludens. Por otro lado, Quesada et al. (2006), indican valores de CL50 en adultos de C. capitata con B. bassiana entre 1x106 y 5,4x107 conidias/mL y con M. anisopliae 4,9x105 conidias/mL con el aislado EAMa 01/58-Su. Lezama et al. (2000), en larvas de tercer instar de A. ludens, citan concentraciones letales medias de 3,7-4,8x105 UFC/mL con los aislamientos Ma2, Ma8 y Ma16 de M. anisopliae. Por otro lado, Destéfano et al. (2005), reportan valores altos de CL50 de 8,44x109 y 12,23x109 conidias/g de M. anisopliae en estudios de aplicación a suelo esterilizado y no esterilizado, calculada de acuerdo con la reducción en la emergencia de adultos de A. fraterculus.
Los tiempos letales para A. obliqua con los tratamientos a base de B. bassiana son inferiores a los notados en ensayos sobre A. ludens y C. capitata,. De la Rosa et al. (2002) con los aislamientos Bb16, Bb26 y Bb24 obtuvieron tiempos de 2,8, 3,7 y 4,2 días, respectivamente sobre A. ludens, mientras Quesada et al. (2006) en adultos de C. capitata encontraron tiempos letales entre 4,8 y 5,3 días. Toledo et al. (2008) refieren un TL50 entre 2,8 y 3,94 para A. ludens y de 3,83 a 4,22 días para C. capitata. En estudios de transmisión horizontal de entomopatógenos en A. ludens, Toledo et al. (2007) encontraron un TL50 entre 4,20 y 4,04 días para LCCP y Bassiamil respectivamente, ambos productos a base de B. bassiana. Igualmente con M. anisopliae los tiempos letales son menores a los informados en la literatura, Lezama et al. (2000) en evaluaciones sobre A. ludens expresan un TL50 de 1,8 a 6,2 días, mientras Dimbi et al. (2003) especifican un TL50 sobre C. capitata y C. rosa var. fasciventris de 4,3 a 6,5 y de 4,7 a 5,7 días, respectivamente, y Quesada et al. (2006) reportan un TL50 de 5,1 días en adultos de C. capitata. Toledo et al. (2008) hallaron un TL50 entre 4,07 y 4,95 días para A. ludens tratada con aislamientos de M. anisopliae.
La tercera etapa de este trabajo se desarrolló con el objetivo de proponer alguna tecnología para el uso de hongos entomopatógenos sobre moscas de las frutas, que permita reducir las aplicaciones de insecticidas químicos. En el ensayo no se evidenció mortalidad a nivel de pupa. Al igual, De la Rosa et al. (2002), con aislamientos de B. bassiana, no registraron mortalidades en pupas de A. ludens, sin embargo, hay evidencias de mortalidad a nivel de pupas de A. ludens, C. capitata, C. corsyra y C. rosa var fasciventris con M. anisopliae y B. bassiana cuando se aplicó el entomopatógeno sobre larvas en prepupa (Lezama et al., 2000; Ekesi et al., 2002; Quesada et al., 2006; Almeida et al., 2007). La diferencia de la mortalidad de pupas en este ensayo con respecto a lo referido por los autores citados anteriormente, se explica por las diferencias de protocolo en las evaluaciones, pues en el presente trabajo los adultos estaban a horas de emergencia (pupas de aproximadamente 14 días). Según los resultados del ensayo de Ekesi et al. (2002) a mayor edad de la pupa el nivel de infección es más bajo, atribuyendo este resultado a la inhabilidad del tubo germinativo para penetrar el integumento de la pupa, debido a la alta esclerotización, igualmente, Quesada et al. (2006) argumentan que una mayor esclerotización de pupas y prepupas de tefritídeos hacen menos susceptible este estado al ataque de hongos entomopatógenos.
Los hongos entomopatógenos para el manejo de moscas de las frutas pueden ser utilizados de varias formas: 1) Dirigidas a la larva, bien sea incorporadas al suelo o en aspersiones superficiales dirigidas a la larva cuando se entierran para empupar; 2) Dirigidos a los adultos jóvenes; 3) Los entomopatógenos pueden ser dispersados liberando insectos estériles infectados. Recientemente Ortiz (2009) aislaron compuestos químicos a partir de hongos entomopatógenos, con los cuales se pueden elaborar bioinsecticidas para el manejo de moscas de las frutas.
Ekesi et al. (2005) encontraron que formulaciones de hongos entomopatógenos incorporados al suelo pueden causar mortalidad de larvas hasta por dos años. En grandes huertos de frutales es difícil incorporar hongos al suelo, la alternativa más viable son las aspersiones sobre la superficie, pero la duración de las conidias sería menor y podría existir mayor probabilidad de fuga de la larva al pasar de la fruta al suelo. Recientemente Toledo et al. (2007) sugieren el uso de insectos estériles infectados para la diseminación del inoculo del entomopatógeno con ventajas como la especificidad sobre la plaga blanco, menor efecto en organismos no blanco y mayor efectividad de la TIE. Sin embargo, esta tecnología no puede ser usada en áreas donde no existan programas específicos de liberación de insectos estériles, pero los trabajos de Toledo et al. (2007), Almeida et al. (2007) y Quesada et al. (2008) muestran un potencial interesante de la transmisión horizontal de hongos entomopatógenos en moscas de las frutas.
Los resultados sugieren que las aspersiones dirigidas a la superficie del suelo, con el objetivo de infectar adultos recién emergidos, es una alternativa viable y serían de mayor facilidad para los agricultores. Un buen conocimiento de los aspectos de biología y ecología de la mosca (conocimiento del comportamiento de las poblaciones, uso de hospederos alternativos, período de desarrollo, entre otros aspectos), permitirán planificar el uso de entomopatógenos de manera eficiente. Igualmente, la capacidad que tienen estos aislamientos de infectar las moscas al momento de la emergencia, sugieren que puede evaluarse la posibilidad de poner en contacto la mosca y el hongo por otros medios, como por ejemplo cebos alimenticios. Los trabajos realizados por Dimbi et al. (2003) sustentan esta posibilidad ya que ellos demostraron que las moscas pueden sufrir infección cuando caminan sobre superficies tratadas. La transmisión horizontal propuesta por Toledo et al. (2007), Almeida et al. (2007) y Quesada et al. (2008) permitiría alcanzar las moscas en hospederos alternativos o refugios.
La mayoría de los estudios de hongos entomopatógenos sobre moscas de las frutas realizados hasta el momento, han sido adelantados en laboratorio o invernadero, no hay trabajos que evalúen la eficiencia de los hongos entomopatógenos en condiciones de cultivos comerciales y, quizás para aumentar la eficiencia del control microbiológico de tefritídeos sea necesario hacer uso de aplicaciones al suelo y en cebos tóxicos, dirigidos a la larva en prepupa y a los adultos, todo dentro de programas de manejo integrado que consideren otras opciones de control.
El empleo de insecticidas sistémicos en aspersiones o en cebos tóxicos trae problemas de contaminación y efectos negativos sobre organismos no blancos (Purcell y Schroeder, 1996). Los hongos entomopatógenos son más seguros y han mostrado selectividad a los enemigos naturales (Ekesi et al. 2005). Los productos biológicos como los hongos entomopatógenos ganan mayor relevancia cuando se trata de manejar moscas de las frutas en países tropicales, caracterizados por su mayor diversidad biológica y donde, por la gran cantidad de hospederos secundarios de la plaga, se hace necesario replantear los sistemas de control con los principios de manejo de plagas en áreas amplias, buscando llegar a los repositorios naturales del insecto.
BIBLIOGRAFÍA
Ali, A., H. Sermann and C. Büttner. 2008. Susceptibility of Ceratitis capitata Wiedemann (Diptera: Tephritidae) to entomopathogenic fungi. Communications in Agricultural and Applied Biological Sciences 73(3): 589-596. [ Links ]
Almeida, J.E.M., A. Batista, F.C. Oliveira and A. Raga. 2007. Pathogenicity of the entomopathogenic fungi and nematode on medfly Ceratitis capitata (Wied.) (Diptera: Tephritidae). In: BioAssay 2:7, http://www.bioassay.org.br/articles/2.7/BA2.7.pdf. 7 p.; consulta: octubre 2009. [ Links ]
Aluja, M. 1994. Bionomics and management of Anastrepha. Annual Review of Entomology 39: 155-178. [ Links ]
Cruz, I.B.M. da, E. Humeres and A.K. Oliveira. 1997. Toxicity of fenthion to Anastrepha fraterculus (Wied.) (Diptera: Tephritideae): dose response analysis. Anais da Sociedade Entomologica do Brasil 26(3): 471-479. [ Links ]
De La Rosa, W., F.L. López and P. Liedo. 2002. Beauveria bassiana as a pathogen of the Mexican fruit fly (Diptera: Tephritideae) under laboratory conditions. Journal of Economic Entomology 95(1): 36-43. [ Links ]
Destéfano, R.R.H., I.J. Bechara, C.L. Messias and A.E. Piedrabuena. 2005. Effectiveness of Metarhizium anisopliae against immature stages of Anastrepha fraterculus fruit fly (Diptera: Tephritideae). Brazilian Journal of Microbiology 36(1): 94-99. [ Links ]
Dimbi, S., N.K. Maniania, A.S. Lux, S. Ekesi and J.K. Mueke. 2003. Pathogenicity of Metarhizium anisopliae (Metsch.) Sorokin and Beauveria bassiana (Balsamo) Vuillemin, to three adult fruit fly species: Ceratitis capitata (Weidemann), C. rosa var. fasciventris Karsch and C. cosyra (Walker) (Diptera: Tephritideae). Mycopathologia 156(4): 375-382. [ Links ]
Ekesi, S., N.K. Maniania and S.A. Lux. 2002. Mortality in three African tephritid fruit fly puparia and adult caused by the entomopathogenic fungi, Metarhizium anisopliae and Beauveria bassiana. Biocontrol Science and Technology 12(1): 7-17. [ Links ]
Ekesi, S., N.K. Maniania, S.A. Mohamed and S.A. Lux. 2005. Effect of soil application of different formulations of Metarhizium anisopliae on African tephritide fruit flies and their associated endoparasitoids. Biological Control 35(1): 83-91. [ Links ]
Enkerlin, W.R. 2005. Impact of fruit fly control programmes using the sterile insect technique. pp. 651-676. In: Dyck, V.A., J. Hendrichs and A.S. Robinson (eds.) Sterile insect technique, principles and practice in area-wide integrated pest management. Springer, Viena. 787 p. [ Links ]
González, E., R. Cásares y R. González. 1997a. Una jaula para la experimentación con Anastrepha spp. (Diptera: Tephritideae). Boletín de Entomología Venezolana 12: 115-117. [ Links ]
González, E., J. Deibis y R. Casares. 1997b. Susceptibilidad de poblaciones adultas, machos y hembras de Anastrepha striata Schiner al insecticida Malatión usando técnicas de aplicaciones tópicas y consumo de cebos tóxicos. Boletín de Entomología Venezolana 12(1): 51-57. [ Links ]
Humeres, E., I.B.M. da Cruz and A.K. Oliveira. 1999. Age and time exposure-related toxicity of fenthion to male and female Anastrepha fraterculus (Wied.) (Diptera: Tephritidae). Anais da Sociedade Entomologica do Brasil 28(2): 285-292. [ Links ]
Lezama, R., A. Trujillo, J. Molina, O. Rebolledo, A.R. Pescador, M. López and M. Aluja. 2000. Virulence of Metarhizium anisopliae (Deuteromycotina: Hyphomycetos) on Anastrepha ludens (Diptera: Tephritideae) laboratory and field trials. Journal of Economic Entomology 93(4): 1080-1084. [ Links ]
Montoya, P. y J. Cancino. 2004. Control biológico por aumento en moscas de las frutas (Diptera: Tephritideae). Folia Entomológica Mexicana 43(3): 257-270. [ Links ]
Montoya, P. and P. Liedo. 2000. Biological control of fruit fly (Diptera: Tephritideae) through parasitoid augmentative releases: current status. pp. 719-732. In: T. Ken-Hong. (ed.). Area-wide control of fruit flies and other insect pests. Penerbit Universiti Sains Malaysia, Penang. 780 p. [ Links ]
Nuñez, L. y R. Guzmán. 1999. Avances sobre la cría artificial de Anastrepha fraterculus (Wied.) (Diptera: Tephritideae) en Colombia. pp. 85-94. In: International Atomic Energy Agency (IAEA) (ed.). The South American fruit fly, Anastrepha fraterculus (Wied.); advances in artificial rearing, taxonomic status and biological studies. IAEA-TECDOC-1064. Vienna, Austria. 202 p. [ Links ]
Ortiz, A., I. Garrido, C. Santiago and E. Quesada. 2009. Purification and characterisation of proteins secreted by the entomopathogenic fungus Metarhizium anisopliae with insecticidal activity against adults of the Mediterranean fruit fly, Ceratitis capitata (Diptera: Tephritidae). Pest Management Science 65(10): 1130-1139. [ Links ]
Purcell, M.F. and W.J. Schroeder. 1996. Effect of silwet L-77 and diazinon on three tephritide fruit flies (Dipetra: Tephritideae) and associated endoparasitoids. Journal of Economic Entomology 89(6): 1566-1570. [ Links ]
Quesada, E., I. Martin, I. Garrido and C. Santiago. 2008. Horizontal transmission of Metarhizium anisopliae among laboratory populations of Ceratitis capitata (Wiedemann) (Diptera: Tephritidae). Biological Control 47(1): 115-124. [ Links ]
Quesada, E, A. Ruiz and C. Santiago. 2006. Laboratory evaluation of entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae against puparia and adults of Ceratitis capitata (Diptera: Tephritideae). Journal of Economic Entomology 99(6): 1955-1966. [ Links ]
Rendon, P., J. Sivinski, T. Holler, K. Bloem, M. López, A. Martínez and M. Aluja. 2006. The effects of sterile males and two braconid parasitioids, Fopius arisanus (Sonan) and Diachasmimorpha krausii (Fullaway) (Hymenoptera), on caged populations of Mediterranean fruit flies, Ceratitis capitata (Wied.) (Diptera: Tephritideae) at various sites in Guatemala. Biological Control 36(2): 224-231. [ Links ]
Robacker, D.C., A.J. Martínez, J.A. García, M. Dayz and C. Romero. 1996. Toxicity of Bacillus thuringiensis to Mexican fruit fly (Diptera: Tephritideae). Journal of Economic Entomology 89(1): 104-110. [ Links ]
Roessler, Y. 1989. Insecticidal bait and cover sprays. pp. 329-336. In: A.S. Robinson and G. Hooper (eds). World crop pests - fruit flies: Their biology, natural enemies and control. Elsevier, Amsterdam. 447 p. [ Links ]
Toledo, J., P. Liedo, S. Flores, S.E. Campos, A. Villaseñor and P. Montoya. 2008. Use of Beauveria bassiana and Metarhizium anisopliae for fruit fly control: a novel approach. pp. 127-132. In: R.L. Sugayama, R.A, Zucchi, S.M. Ovruski and J. Sivinski. (eds.). Fruit Flies of economic importance: From basic to applied knowledge. Press Color Gráficos Especializados Ltda., Salvador, Brazil. 351 p. [ Links ]
Toledo, J., P. Liedo, T. Williams and J. Ibarra. 1999. Toxicity of Bacillus thuringiensis ß-exotoxin to three species of fruit flies (Diptera: Tephritideae). Journal of Economic Entomology 92(5): 1052-1056. [ Links ]
Toledo, J., R. Romero and I.E. Jorge. 2006. Efficiency of Heterorhabditis bacteriophora (Nematodo: Heterorhabditidae) on Anastrepha serpentina (Diptera: Tephritideae) larvae under laboratory conditions. Florida Entomologist 89(4): 524-526. [ Links ]
Toledo, J., S.E. Campos, S. Flores, P. Liedo, J.F. Barrera, A. Villaseñor and P. Montoya. 2007 Horizontal transmisión of Beauveria bassiana in Anastrepha ludens (Diptera: Tephritidae) under laboratory and field cage conditions. Journal of Economic Entomology 100(2): 291-297. [ Links ]
White, I.M. and M.M. Elson. 1992. Fruit flies of economic significance: Their identification and bionomics. CAB International, London. 601 p. [ Links ]

