










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.65 no.1 Medellín Jan./June 2012
Evaluación in vitro de Inductores de Resistencia sobre Mycosphaerella fijiensis Morelet
In vitro Evaluation of Inductors of Resistance on Mycosphaerella fijiensis Morelet
Ángela María Mogollón Ortiz1 y Jairo Castaño Zapata2
1 Magíster en Fitopatología. Universidad de Caldas, Facultad de Ciencias Agropecuarias. Calle 65 No. 26-10. Manizales, Colombia. <ammogollon@hotmail.com>
2 Profesor Titular. Universidad de Caldas, Facultad de Ciencias Agropecuarias. Calle 65 No. 26-10. Manizales, Colombia. <jairo.castano_z@ucaldas.edu.co>
Recibido: Mayo 16 de 2011; aceptado: Febrero 17 de 2012.
Resumen. La sigatoka negra (Mycosphaerella fijiensis M.), es la enfermedad foliar más importante del plátano y banano, ocasiona pérdidas entre el 30 y 40%. Su manejo basado en agroquímicos, representa el 40% de los costos de producción. Con el fin de encontrar nuevas alternativas de manejo de la enfermedad en plátano, se evaluó el efecto in vitro sobre el hongo de los inductores de resistencia ácido salicílico (AS), acibenzolar-S-metil (ASM), ácido 2,6- dicloroisonicotinico (INA), ácido 3- aminobutanoico (BABA) y fosfito de potasio, y los fungicidas propiconazol y clorotalonil. Cada producto fue considerado como un experimento independiente con un diseño completamente aleatorio a una vía, constituido por cuatro tratamientos y 15 repeticiones. Se evaluó: número de colonias cm2-1, tamaño de colonias (mm), esporulación y germinación (%) de M. fijiensis. El efecto de los inductores no mostró inhibición total sobre el crecimiento del hongo, pero, todas las variables de respuesta, a excepción de la germinación con AS, INA y BABA, presentaron diferencias significativas entre las concentraciones evaluadas. El número promedio de colonias cm2-1 con los inductores osciló entre 1,5 y 3,5 mientras el testigo presentó 2,1 y 3,8; el tamaño de las colonias varió entre 1 y 4,6 mm y en el testigo 3,8 y 4,6 mm; el número de conidias mL-1 fluctuó entre 1.142 y 4.700 y en el testigo 4.386 y 5.783; y la germinación de conidias con ASM y fosfito de potasio alternó entre 27 y 72,6%, mientras el testigo presentó 74 y 79%. Propiconazol y clorotalonil inhibieron completamente al hongo.
Palabras clave: Paracercospora fijiensis, sigatoka negra, hongo, agar V8.
Abstract. Black sigatoka (Mycosphaerella fijiensis M.), is the most important foliar disease of both banana and plantain, causing yield losses between 30 and 40%. Its management based on fungicides, represents 40% of the production costs. In order to find new alternatives to manage the disease in plantain, it was evaluated in vitro the effect on the fungus of the inductors of resistance salicylic acid (SA), acibenzolar-S-methyl (ASM), 2,6-dicloroisonicotinic acid (INA), 3-butanoic acid (BABA) and potassium phosphite, as well the fungicides propoconazole and clorothalonil. Each product was considered as an independent experiment in a completely randomized design in one way and conformed by four treatments and fifteen repetitions per treatment. It was evaluated: number of colonies cm2-1, size of colonies (mm), sporulation and germination (%) of M. fijiensis. The effect of the inductors of resistance did not show total inhibition on the fungus growth; however, all the variables, except the germination with SA, INA and BABA, showed significant differences between the concentrations evaluated. The average number of colonies cm2-1 with the inductors of resistance ranged between 1.58 and 3.5, while the control had 2.1 and 3.8; the colony size ranged between 1 and 4.6 mm and the control 3.8 to 4.6 mm; the number of conidia mL-1 ranged between 1,142 and 4,700 and in the control 4,386 and 5,783, and the germination of conidia with ASM and potassium phosphite ranged between 27 and 72.6% while in the control was between 74 and 79%. Both fungicides propiconazole and chlorothalonil totally inhibited the fungus growth.
Key words: Paracercospora fijiensis, black sigatoka, fungus, agar V8.
La sigatoka negra causada por el hongo Mycosphaerella fijiensis Morelet, anamorfo Paracercospora fijiensis (Morelet) Deighton, es considerada la enfermedad más importante que afecta la producción comercial de banano y plátano (Musa spp.) en la mayoría de las regiones productoras del mundo (Marín et al., 2003), debido a que causa altas pérdidas económicas como resultado de la disminución en el rendimiento del cultivo y la calidad del fruto (Arzanlou et al., 2007).
En la actualidad existen métodos químicos, culturales y de variedades resistentes para el manejo de la enfermedad, sin embargo, el problema persiste y se hace necesaria la búsqueda de nuevas alternativas. El tratamiento de plantas con inductores de resistencia es un excelente método utilizado contra hongos, bacterias, virus y nemátodos, mecanismo que se ha denominado resistencia sistémica adquirida (RSA) (Potlakayala et al., 2007; Vallad y Goodman, 2004; Walters et al., 2005). La RSA consiste en la expresión de un conjunto de respuestas de las plantas después de la infección por patógenos y luego del tratamiento químico con productos como ácido salicílico, acibenzolar-S-metil, ácido 2,6-dicloroisonicotinico, ácido 3-aminobutanoico, y fosfito de potasio (Bostock, 2005; Durrant y Dong, 2004; Hammerschmidt, 2007; Vallad y Goodman, 2004).
Resultados del efecto in vitro de los inductores de resistencia sobre patógenos han demostrado que no afectan directamente el desarrollo de estos. En lo referente al ácido salicílico (AS), este inductor no afecta la germinación ni el crecimiento de Phytophthora infestans (Mont.) De Bary; sin embargo, el tizón tardío en plantas de papa se redujo debido a reacciones de la pared celular y a la expresión de genes que codifican proteínas relacionadas con patogénesis (PR) luego del tratamiento con este producto (Halim et al., 2007), estas respuestas también fueron generadas en diversas plantas dicotiledóneas tratadas con ácido 2,6-dicloroisonicotinico, análogo funcional del AS, frente a virus, bacterias y hongos fitopatógenos (Chaturvedi y Shah, 2007). Plantas de café (Coffea arabica L.) sometidas a tratamiento con acibenzolar-S-metil, mostraron incremento de la actividad de las enzimas líticas b-1,3 glucanasa y quitinasa, enzimas asociadas a la RSA, las cuales afectaron a Hemileia vastratix Berk y Br. (Guzzo, 2004). Así mismo, Márquez y Castaño (2007), demostraron el efecto de la inducción de resistencia en plántulas de plátano Dominico-Hartón, después de tratar los cormos con este producto. Por su parte Querino et al. (2005) documentaron el efecto indirecto de ácido 3-aminobutanoico sobre Fusarium oxysporum F. sp. cubense (Sm.) Snyder & Hansen, tras demostrar la reducción de la severidad del mal de Panamá atribuida a la RSA en plantas de banano tratadas con el inductor. Se ha comprobado que el fosfito de potasio actúa indirectamente sobre patógenos como Mycosphaerella spp. en plátano Dominico-Hartón (Mogollón y Castaño, 2011).
Según Orober et al. (2002), el efecto indirecto del fosfito de potasio sobre Colletotrichum lagenarium (Pass.) Ell. & Halst se atribuye a la respuesta de hipersensibilidad y al aumento local y sistémico de AS. En plantas de pepino tratadas, el AS actúa como molécula mensajera activando la síntesis de proteínas relacionadas con patogénesis (PR). En contraste con este reporte, propiconazol y clorotalonil actúan directamente sobre los patógenos, el primero, interfiriendo en la síntesis del ergosterol de la membrana celular del patógeno, actuando en el momento de la penetración y principalmente en la formación de haustorios secundarios del hongo dentro de los tejidos de la planta (Köller, 1992) y clorotalonil por la unión de las moléculas a los grupos sulfhídrico de los aminoácidos, inactivando las enzimas que intervienen en el ciclo de Krebs y así impidiendo la producción de ATP, dando como resultado la muerte de las células (Washington et al., 1998).
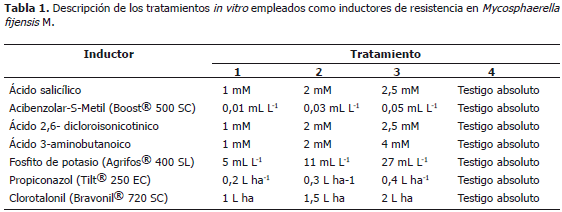
Debido al alto riesgo de desarrollo de resistencia M. fijiensis hacia los fungicidas sistémicos utilizados para el control de la sigatoka negra y al desconocimiento del efecto in vitro de los inductores de resistencia sobre el hongo, en este estudio se evaluaron los inductores de resistencia acido salicílico, acibenzolar-S-metil (Boost® 500 SC), ácido 2,6 dicloroisonicotinico, ácido 3-aminobutanoico y fosfito de potasio (Agrifos® 400 SL), y los fungicidas propiconazol (Tilt® 250 EC) y clorotalonil (Bravonil® 720 SC).
MATERIALES Y MÉTODOS
El experimento se llevó a cabo en el Laboratorio de Fitopatología, Departamento de Fitotecnia de la Universidad de Caldas (Manizales, Colombia). El aislamiento de M. fijiensis, fue obtenido utilizando el método de siembra directa propuesto por Castaño (1997) con algunas modificaciones en los desinfectantes y en el tiempo de exposición a éstos.
La identificación de M. fijiensis, se realizó tomando conidias del cultivo puro y colocándolas sobre un porta objetos con azul de lactofenol (ácido láctico 25 mL, glicerina 50 mL, agua destilada 25 mL, azul de algodón 0,05 g). Las características típicas de las conidias es la cicatriz (hilio) presente tanto en la base de los conidióforos como en las conidias (Castaño y del Rio, 1994). Las conidias se caracterizan por ser alargadas, septadas, hialinas y aciculares (Fullerton, 1994).
El diseño estadístico fue completamente aleatorio a una vía con 15 repeticiones por tratamiento. En total se realizaron de manera independiente siete experimentos que correspondieron a cinco inductores de resistencia, dos fungicidas y cuatro tratamientos (concentraciones) (Tabla 1). Por cada producto se evaluaron 60 cajas, es decir cada una de las concentraciones estuvo constituida por 15 cajas Petri. La evaluación de los tratamientos contra el hongo se efectuó en agar V8 (200 mL de jugo V8 Campbell®, 3 g de CaCO3, 20 g agar-bacteriológico y 800 mL L-1 de agua) modificado con cada inductor o fungicida, los productos fueron aplicados directamente al agar una vez fue esterilizado. Se utilizó agar V8 debido a que no se logró esporulación del hongo en PDA (Papa Dextrosa Agar). A partir de una solución madre de conidias de M. fijiensis, se tomaron alícuotas de 50 mL para ser sembradas en el medio de cultivo modificado con cada tratamiento. El hongo fue mantenido a una temperatura de 26 °C y luz continua durante 15 días en una incubadora Precision Scientific® (Modelo Standar 815, Referencia 1021).
Todas las variables fueron evaluadas 15 días después de la siembra del hongo.
Número de colonias cm2-1. Se cuantificó el número de colonias para cada caja Petri evaluada. Se utilizó una porción de hoja milimetrada en la base de la caja Petri como guía en el conteo.
Tamaño de colonias (mm). Se midió colocando una regla en la base de cada caja Petri y ubicando las colonias a medir.
Esporulación (conidias mL-1). Se definió el número de conidias mL-1 utilizando la cámara de Neubauer Boeco®; para ello se tomaron alícuotas de 20 mL a partir de una solución de conidias de M. fijiensis obtenida mediante la adición de 10 mL de agua destilada estéril a cada caja Petri y posterior raspado de las colonias con un asa redonda. Para establecer el número de conidias se utilizó la siguiente fórmula (Castaño, 1998):
X= E * 2000
X= No. de esporas mL-1; E= Esporas en los cinco cuadrados de 1 mm2.
Germinación (%). Se tomaron alícuotas de 20 mL a partir de una suspensión que resultó de la adición de 10 mL de agua y posterior raspado de las colonias de cada caja Petri a evaluar. Las conidias fueron sembradas en cajas Petri servidas con Agar-Agar y 32 h después de la siembra se contó el número de esporas germinadas de un total de 100, con ayuda de un microscopio compuesto de luz LW Scientific® (Modelo Revelation III) utilizando el objetivo de 40 X.
Una conidia se consideró germinada cuando el tubo germinativo alcanzó el doble de la longitud de la conidia.
Análisis estadístico. Se usó el programa estadístico Statgraphics Centurión XV para realizar el análisis de varianza, la prueba de comparación de medias de Tukey al 5% y la prueba de Kruskal Wallis al 5%. Debido a que no se cumplieron los supuestos del análisis de varianza en los tratamientos con propiconazol y clorotalonil, fue necesario utilizar pruebas no paramétricas, por tal razón se empleó la prueba Kruskal Wallis, la cual evalúa la hipótesis de que las medianas dentro de cada uno de los niveles de tratamiento son iguales. Cuando se presentaron diferencias significativas entre tratamientos se usó el gráfico de cajas y bigotes para determinar cuáles medianas eran significativamente diferentes de otras.
RESULTADOS
Efecto del ácido salicílico (AS). Los análisis de varianza del número y tamaño de colonias y esporulación indicaron diferencias altamente significativas entre tratamientos (P= 0,0000), a excepción de la variable germinación, en la cual no se presentaron diferencias (P= 0,0594).
La prueba de Tukey al 5% no indicó diferencias significativas entre los valores promedio del número y tamaño de las colonias cuando se evaluaron las concentraciones de 1, 2 y 2,5 mM, sin embargo, se presentaron diferencias de estos tratamientos con respecto al testigo (Tabla 2).
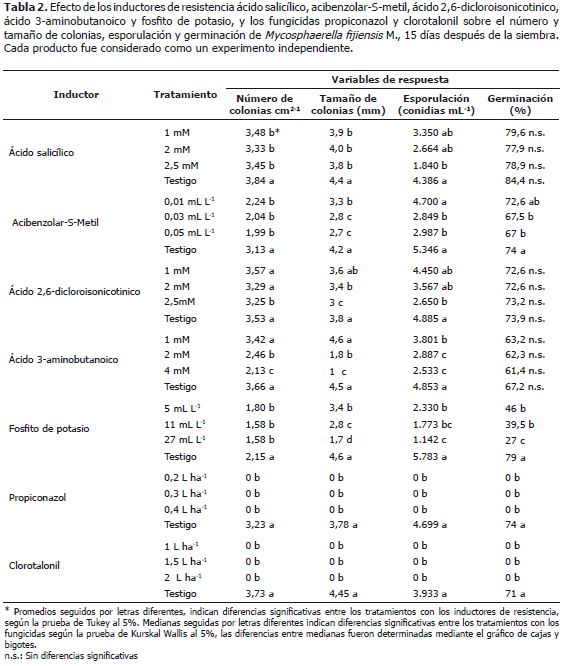
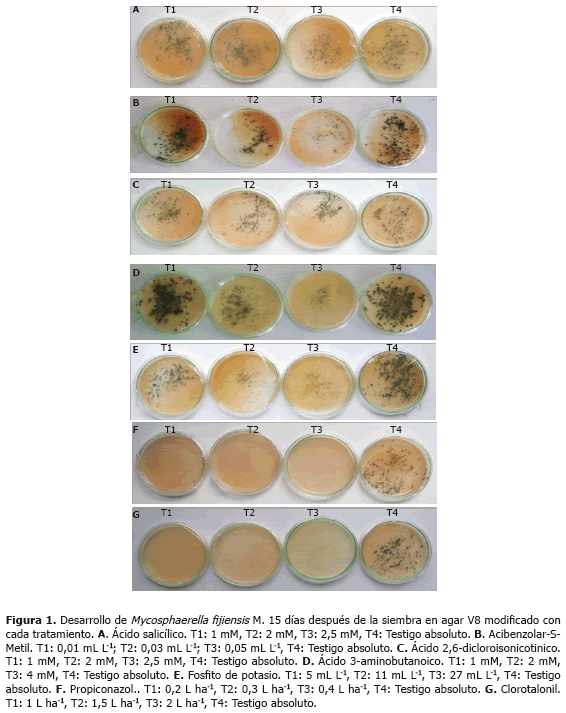
Los valores promedio del número de colonias para los tratamientos que incluyeron AS oscilaron entre 3,33 a 3,48 colonias cm2-1, en comparación con el testigo que registró 3,84 colonias cm2-1 (Figura 1 A); para el tamaño de las colonias los valores promedio variaron entre 3,8 y 4 mm, con respecto al testigo que presentó un valor promedio de 4,4 mm; mientras que para la variable esporulación la mayor reducción se logró con la concentración de 2,5 mM cuyo valor promedio de conidias mL-1 fue de 1.840, el cual fue significativamente menor que los valores registrados para las demás concentraciones que alternaron entre 2.664 y 3.350 conidias mL-1 y el testigo absoluto con 4.386 conidias mL-1 (Tabla 2).
Los resultados demostraron que las diferentes concentraciones de AS evaluadas, redujeron los valores promedio del número y tamaño de colonias y esporulación con respecto al testigo, sin embargo, la germinación de las conidias de M. fijiensis no fue inhibida totalmente. Estos resultados coinciden con los de Zhang et al. (2002), quienes demostraron que concentraciones entre 0,01 y 1.000 mg mL-1 de AS no inhiben la germinación de esporangios de Peronospora tabacina Adam, ni tienen efecto sobre la longitud de los tubos germinativos, no obstante, Chaturvedi y Shah (2007) atribuyen al AS la función de molécula activadora de la inducción de resistencia en plantas, al desencadenar la respuesta de hipersensibilidad, el refuerzo de la pared celular y la activación de genes que codifican para las proteínas relacionas con patogénesis (PR), de manera similar a cuando ocurre la infección por patógenos.
Efecto de acibenzolar-S-metil (ASM). Los análisis de varianza de las variables número y tamaño de colonias, esporulación y germinación de M. fijiensis indicaron diferencias altamente significativas (P= 0,0000), presentándose un menor desarrollo del patógeno en los tratamientos con ASM en relación con el testigo.
El valor promedio del número de colonias cm2-1 fue de dos a diferencia del testigo que fue de tres (Figura 1 B); el tamaño promedio de las colonias osciló entre 2,7 y 3,3 mm, con respecto a 4,2 mm del testigo, la mayor reducción del crecimiento del hongo, se obtuvo con la concentración más alta de 0,05 mL L-1, la cual no presentó diferencias significativas con respecto a 0,03 mL L-1, pero si las mostró con la menor concentración (Tabla 2); el número promedio de conidias mL-1 para los tratamientos con el inductor varió entre 2.987 y 4.700, con respecto a 5.346 conidias mL-1 del testigo. Los porcentajes promedio de germinación fluctuaron entre 67 y 72,6% en comparación con 74% del testigo (Tabla 2).
La reducción en los promedios del número y tamaño de colonias, esporulación y germinación con los tratamientos que incluyeron ASM no llegó a ser absoluta, comprobando que la aplicación de este producto a concentraciones de 0,01; 0,03; y 0,05 mL L-1, reduce, mas no evita, el desarrollo de M. fijiensis. Estos resultados coinciden con reportes previos que indican que la reducción en el desarrollo de los patógenos luego de la aplicación de ASM en plantas, está relacionada con la activación de los mecanismos de defensa de estas, más que con su efecto directo sobre los patógenos. Al respecto, Friedrich et al. (1996) concluyen de experimentos in vitro con ASM a una de concentración de 1,4 mM, que éste no tiene actividad fungicida, incluso con concentraciones más altas sobre hongos como: Alternaria brassicae (Berk.) Sacc., Botrytis cinerea (de Bary) Whetzel, Ceratocystis ulmi (Buis.) Moreau, Cladosporium cucumerinum Ellis & Arthur, Helminthosporium oryzae (Brenda de Hann) Schoemarker, Penicillium digitatum (Pers.) Sacc, Phytophthora infestans (Mont.) de Bary, Pyricularia oryzae Cavara, Rhizoctonia solani Kühn, Ustilago maydis (DC.) Corda y Verticillium dahliae Kleb; sin embargo, en estudios a nivel in vivo, Jiang et al. (2008) demostraron que plántulas de pera tratadas con ASM presentaron respuestas asociadas a la inducción de resistencia frente a la roña causada por Venturia nashicola Tanaka y Yamamoto, dicha resistencia incluyó la activación de la síntesis de proteínas relacionadas con patogénesis, generación de especies oxigeno reactivas y lignificación de la pared celular. También in vivo, Márquez y Castaño (2007), demostraron que plántulas de plátano provenientes de cormos tratados con ASM en concentración de 0,005 mL L-1 de agua, presentaban los niveles más bajos de grado de evolución de síntomas y severidad de las sigatokas amarilla (M. musicola Leach) y negra (M. fijiensis), demostrando el efecto de inducción de resistencia a estas enfermedades en estado de plántula.
Efecto de ácido 2,6-dicloroisonicotinico (INA). Los análisis de varianza indicaron diferencias estadísticas altamente significativas sobre el número y tamaño de colonias y esporulación (P= 0,0000), más no sobre la germinación del hongo (P= 0,9315).
La concentración más alta evaluada (2,5 mM), presentó los valores promedios más bajos para las variables de respuesta número (Figura 1 C) y tamaño de colonias, y esporulación con respecto a las demás concentraciones (1 y 2 mM) y el testigo absoluto. Los valores promedio del número de colonias cm2-1 para los tratamientos que incluyeron INA oscilaron entre 3,25 a 3,57, mientras que en el testigo se registraron 3,53 colonias cm2-1; el tamaño promedio de las colonias varió entre 3 y 3,6 mm, mientras que para el testigo fue de 3,8 mm; el número de conidias mL-1 fluctuó entre 2.650 y 4.450, con respecto a 4.885 conidias mL-1 del testigo (Tabla 2).
En resumen, el INA no inhibió el desarrollo del hongo a pesar que se observaron reducciones en los valores promedio de las variables de respuesta número y tamaño de colonias, y esporulación con respecto al testigo. Un estudio in vitro realizado por Dann et al. (1998), indicó que el efecto de INA sobre Sclerotinia spp. Lib., aplicado a una concentración de 400 ppm no inhibe el crecimiento del hongo; no obstante señalan que reduce la severidad de la podredumbre blanda del tallo de la soya lo que sugiere una estimulación de los mecanismos de defensa de la planta, luego del tratamiento con el producto.
Efecto de ácido 3-aminobutanoico (BABA). Con respecto a la acción del BABA sobre las variables de respuesta número y tamaño de colonias, y esporulación, los análisis de varianza indicaron diferencias altamente significativas (P=0,0000), pero éstas no se presentaron para la variable germinación de las esporas (P=0,0956).
Los valores más bajos en los promedios de las variables de respuesta se alcanzaron con la dosis más alta evaluada (4 mM). El número de colonias cuando se estimó el efecto de BABA varió entre 2,13 a 3,42 colonias cm2-1, en comparación con el testigo que registró 3,66 colonias cm2-1 (Figura 1 D); para el tamaño de colonias, el valor promedio fluctuó entre 1 y 4,6 mm, en comparación el testigo que presentó un promedio de 4,5 mm; el número de conidias osciló entre 2.533 y 3.801 mL-1 en comparación con 4.853 conidias mL-1 del testigo (Tabla 2).
El efecto de BABA sobre M. fijiensis mostró reducciones en el número y tamaño de colonias, y esporulación en comparación con el testigo, aunque, el desarrollo del patógeno continuó a lo largo del experimento. Estos resultados coinciden con los de Sunwoo et al. (1996), Tosi y Zazzerini (1999), Reuveni et al. (2003) quienes demostraron que BABA no afecta la germinación ni la elongación de tubos germinativos de Peronospora tabacina Adam, Plasmopara helianthi Novot y Alternaria alternata (Fries) Keissler, sin embargo, se conoce la capacidad de BABA para proteger los cultivos como la alcachofa frente Sclerotinia sclerotiorum (Lib.) de Bary, al inducir la rápida acumulación de enzimas b-1,3-glucanasa y quitinasa luego del tratamiento con el inductor (Marcucci et al., 2010).
Efecto de fosfito de potasio. Los análisis de varianza del número y tamaño de colonias, esporulación y germinación, indicaron diferencias altamente significativas entre tratamientos (P= 0,0000).
La mayor reducción se observó utilizando la dosis más alta (27 mL L-1) (Figura 1 E). Para el número de colonias no se encontraron diferencias significativas entre las concentraciones evaluadas (5, 11 y 27 mL L-1) pero si de éstas con respecto al testigo. Los valores promedio del número de colonias cm2-1 en los tratamientos que incluyeron fosfito de potasio oscilaron entre 1,58 a 1,80 a diferencia del testigo que registró 2,15 colonias cm2-1; el valor promedio del tamaño de las colonias varió entre 1,7 y 3,4 mm en comparación con el testigo que fue de 4,6 mm; los promedios del número de conidias mL-1 fluctuaron entre 1.142 y 2.330, con respecto al el testigo que registró 5.783 conidias mL-1 y el porcentaje de germinación alternó entre 27 y 46% en comparación con 79% del testigo (Tabla 2).
En este estudio, el desarrollo M. fijiensis no demostró una inhibición absoluta, sólo algunas reducciones en los valores promedio del número y tamaño de colonias, producción de conidias y germinación, a pesar de la alta concentración evaluada (27 mL L-1). Estos resultados coinciden con informes que mencionan que este producto interviene en la activación del sistema de defensas de las plantas, sin actuar directamente sobre los patógenos. Al respecto, Orober et al. (2002), atribuyen el control de la Antracnosis [Colletotrichum lagenarium (Pass.) Ell. & Halst] del pepino al aumento local y sistémico de AS y la respuesta de hipersensibilidad luego del tratamiento con el inductor, de igual manera, Eshraghi et al. (2011) documentaron que plantas de arabidopsis [Arabidopsis thaliana (L.) Heynh] tratadas con fosfito de potasio e infectadas con Phytophthora cinnamomi Rands, mostraron más tempranamente la inducción de respuestas de defensa a diferencia de las plantas no tratadas, las respuestas generadas incluyeron la transcripción de genes de defensa, lignificación de la pared celular y producción de peróxido de hidrógeno (H2O2). Recientemente, Mogollón y Castaño (2011), demostraron que el efecto de inductores de resistencia sobre los valores bajos de índice severidad (%), grado de evolución de los síntomas y tasa de desarrollo (r) de las sigatokas amarilla y negra en plántulas de Dominico-Hartón, fue producto de la activación de los mecanismos de defensa de las plantas, en donde el Fosfito de potasio, fue el mejor inductor, sin presentar diferencia significativa con respecto a propiconazol.
Efecto de propiconazol y clorotalonil. Los tratamientos que incluyeron propiconazol y clorotalonil inhibieron completamente el crecimiento de M. fijiensis (Figuras 1 F, G).
La prueba de Kruskal-Wallis indicó diferencias significativas entre las medianas de los tratamientos para las variables número de colonias (P= 2,08422E-12), tamaño de colonias (P= 0,00034576), esporulación (P= 2,03393E-12) y germinación (P= 2,07856E-12) cuando se evaluó el efecto de propiconazol y de igual manera para las variables número de colonias (P= 2,09555E-12), tamaño de colonias (P= 0,00034576), esporulación (P= 1,91558E-12) y germinación (P = 2,05336E-12) cuando se evaluó el efecto de clorotalonil.
Los gráficos de cajas y bigotes no indicaron diferencias significativas entre las dosis evaluadas de propiconazol (0,2; 0,3 y 0,4 L ha-1) y clorotalonil (1; 1,5; 2 L ha-1), sin embargo, se presentaron diferencias altamente significativas de las tres concentraciones evaluadas de cada fungicida con respecto al testigo absoluto (Tabla 2). Los resultados demuestran que estos fungicidas son igual de efectivos en todas las dosis empleadas.
Koné et al. (2009), afirman que en evaluaciones in vitro de los fungicidas pertenecientes a los triazoles se inhibe totalmente el crecimiento del hongo. Por su parte el efecto de Clorotalonil sobre M. fijiensis evaluado por Washington et al. (1998), demuestra que la elongación de los tubos germinativos es detenida luego del tratamiento con el fungicida por unión de las moléculas del producto a los grupos sulfidrilos de las enzimas involucradas en el ciclo de Krebs.
CONCLUSIONES
El tratamiento con los inductores de resistencia ácido salicílico (AS), acibenzolar-S-metil (ASM), ácido 2,6-dicloroisonicotinico (INA), ácido 3-aminobutanoico (BABA) y fosfito de potasio disminuyeron el número y tamaño de colonias, esporulación y la germinación de conidias de M. fijiensis, presentándose mayor reducción con las concentraciones más altas evaluadas.
El efecto in vitro de los inductores de resistencia no demostró inhibición absoluta en el desarrollo de M. fijiensis, estos resultados coinciden con estudios que evidencian que los inductores de resistencia no actúan directamente sobre los patógenos, sin embargo, activan las respuestas de defensas en las plantas tales como: síntesis de proteínas relacionadas con patogénesis, aumento de lignificación de pared celular, respuesta de hipersensibilidad y síntesis de fitoalexinas.
Propiconazol y clorotalonil en las diferentes concentraciones evaluadas, inhibieron de manera absoluta el desarrollo de M. fijiensis, demostrando su efecto directo frente al patógeno. Ninguno de los inductores de resistencia evaluados se asemejó al comportamiento inhibitorio de los fungicidas.
BIBLIOGRAFÍA
Arzanlou, M., E. Abeln, G. Kema, C. Waalwijk, J. Carlier, I. de Vries, M. Guzmán and P. Crous. 2007. Molecular diagnostics for the sigatoka disease complex of banana. Phytopathology 97(9): 1112 -1118. [ Links ]
Bostock, R. 2005. Signal crosstalk and induced resistance: straddling the line between cost and benefit. Annual Review of Phytopathology 43(1): 545-580. [ Links ]
Castaño, J. y L. del Río. 1994. Sigatokas del banano. pp. 217-218. En: Guía para el Diagnóstico y Control de Enfermedades en Cultivos de Importancia Económica. Tercera edición. Zamorano Academic Press, Honduras. 290 p. [ Links ]
Castaño, J. 1997. Manejo de muestras y secuencia de diagnóstico. pp. 1-11. En: Castaño, J. and L. del Rio. (eds.). Manual para el diagnóstico de hongos, bacterias, virus y nemátodos fitopatógenos. Universidad de Caldas, Zamorano Academic Press, Manizales, Colombia. 210 p. [ Links ]
Castaño, J. 1998. Prácticas de laboratorio de Fitopatología. Segunda edición. Centro Editorial Universidad de Caldas. Zamorano Academic Press, Escuela Agrícola Panamericana, Zamorano, Honduras. 103 p. [ Links ]
Chaturvedi, R. and J. Shah. 2007. Salicylic acid in plant disease resistance. pp. 335-370. In: Hayat, S., A. Ahmad (eds.). Salicylic acid a plant hormone. Springer, The Netherlands. 401 p. [ Links ]
Dann, E., B. Diers, J. Byrum and R. Hammerschmidt. 1998. Effect of treating soybean with 2,6- dichloroisonicotinic acid (INA) and benzothiadiazole (BTH) on seed yields and the level of disease caused by Sclerotinia sclerotiorum in field and greenhouse studies. European Journal of Plant Pathology 104(3): 271-278. [ Links ]
Durrant, W. and X. Dong. 2004. Systemic acquired resistance. Annual Review of Phytopathology 42(1): 185-209. [ Links ]
Eshraghi, L., J. Anderson, N. Aryamanesh, B. Shearer, J. McComb, G. Hardy and P. O'Brien. 2011. Phosphite primed defense responses and enhanced expression of defense genes in Arabidopsis thaliana infected with Phytophthora cinnamomi. Plant Pathology 60(6): 1086-1095. [ Links ]
Friedrich, L., K. Lawton, W. Ruess, P. Masner, N. Specker, M. Gut, B. Mejer, S. Dincher, T. Staub, S. Uknes, J. Métraux, H. Kessmann and J. Ryals. 1996. A benzothidiazole derivate induces systemic acquired resistance in tobacco. The Plant Journal 10(1): 61-70. [ Links ]
Fullerton, R. 1994. Sigatoka leaf diseases. pp. 12-14. In: Ploetz, R.; G. Zentmeyer, W. Nishijima, K. Rohrbach and H. Ohr. (eds.). Compendium of tropical fruit diseases. APS Press, St. Paul, Minnesota, USA. 111 p. [ Links ]
Guzzo, S.D. 2004. Aspectos bioquímicos e moleculares da resistência sistêmica adquirida em cafeeiro contra Hemileia vastatrix. Tese de Doutorado, Centro de Energia Nuclear na Agricultura, Universidade de São Paulo, Piracicaba. 236 p. [ Links ]
Halim, V., L. Eschen, S. Altmann, M. Birschwilks, D. Scheel and S. Rosahl. 2007. Salicylic acid is important for basal defense of Solanum tuberosum against Phytophthora infestans. Molecular Plant-Microbe Interactions 20(11): 1346-1352. [ Links ]
Hammerschmidt, R. 2007. Definition and some history. pp. 1-8. In: Walters, D., A. Newton, G. Lyon. (eds.). Induced resistance for plant defense. A sustainable approach to crop protection. Blackwell Publishing, Oxford, UK. 272 p. [ Links ]
Jiang, S., P. Park and H. Ishii. 2008. Ultrastructural study on acibenzolar-S-methyl-induced scab resistance in epidermal pectin layers of Japanese pear leaves. Phytopathology 98(5):585-591. [ Links ]
Köller, W. 1992. Antifungal agents with target sites in sterol functions and biosynthesis. pp. 119-206. In: Köller, W. (ed). Target sites of fungicide action. CRC Press, Boca Raton, Florida. 328 p. [ Links ]
Koné, D, O. Badou, E. Bromisso, B. Camara and S. Ake. 2009. In vitro activity of different fungicides on the growth in Mycosphaerella fijiensis var. difformis Stover and Dickson, Cladosporium musae Morelet and Deightoniella torulosa (Syd.) Ellis, isolated parasites of the banana phyllosphere in the Ivory Coast. Comptes Rendus de l'Académie des Sciences 332(5): 448-55. [ Links ]
Marcucci, E., M. Aleandri, C. Gabriele and P. Magro. 2010. Induced resistance by b-aminobutyric acid in artichoke against white mould caused by Sclerotinia sclerotiorum. Journal of Phytopathology 158(10): 659-667. [ Links ]
Marín, D., R. Romero, M. Guzmán and T. Sutton. 2003. Black sigatoka: an increasing threat to banana cultivation. Plant Disease 87(3): 208-222. [ Links ]
Márquez, L.J. y J. Castaño. 2007. Inducción de resistencia a sigatokas en plántulas de plátano Dominico-Hartón. Agronomía 15(2): 49-57. [ Links ]
Mogollón, A.M. y J. Castaño. 2011. Efecto de inductores de resistencia en plántulas de plátano Dominico-Hartón (Musa balbisina AAB) contra Mycosphaerella spp. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 35(137): 463-471. [ Links ]
Orober, M., J. Siegrist and H. Buchenauer. 2002. Mechanisms of phosphate-induced disease resistance in cucumber. European Journal of Plant Pathology 108(4): 345-353. [ Links ]
Potlakayala, S., D. Reed, P. Covello and P. Fobert. 2007. Systemic acquired resistance in canola is linked with pathogenesis-related gene expression and requires salicylic acid. Phytopathology 97(7): 794-802. [ Links ]
Querino, C., D. Laranjeira, R. Coelho e A. Matos. 2005. Efeito de dois indutores de resistência sobre a severidade do mal-do-Panamá. Fitopatologia Brasileira 30(3): 239-243. [ Links ]
Reuveni, M., D. Sheglov and Y. Cohen. 2003. Control of moldy-core decay in apple fruits by b- aminobutyric acids and potassium phosphites. Plant Disease 87(8): 933-936. [ Links ]
Sunwoo, J.Y., Y.K. Lee and B.K. Hwang. 1996. Induced resistance against Phytophthora capsici in pepper plants in response to DL-b-amino-n-butyric acid. The European Journal of Plant Pathology 102(7): 663-670. [ Links ]
Tosi, L. and A. Zazzerini. 1999. Induced resistance against Plasmopara helianthi in sunflower plants by DL-b-amino-n-butyric acid. Journal of Phytopathology 146(5-6): 295-299. [ Links ]
Vallad, G. and R. Goodman. 2004. Systemic acquired resistance and induced systemic resistance in conventional agriculture. Crop Science 44(6): 1920-1934. [ Links ]
Walters, D., D. Walsh, A. Newton and G. Lyon. 2005. Induced resistance for plant disease control: maximizing the efficacy of resistance elicitors. Phytopathology 95(12): 1368-1373. [ Links ]
Washington, J.R., J. Cruz, F. López and M. Fajardo. 1998. Infection studies of Mycosphaerella fijiensis on banana and the control of black sigatoka with chlorothalonil. Plant Disease 82(11): 1185-1190. [ Links ]
Zhang, S., A. Moyne, M. Reddy and J. Kloepper. 2002. The role of salicylic acid in induced systemic resistance elicited by plant growth-promoting rhizobacteria against blue mold of tobacco. Biological Control 25(3): 288-296. [ Links ]

