










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.65 no.1 Medellín Jan./June 2012
Caracterización Citogenética de Cinco Genotipos de Papa Criolla, Solanum phureja (Juz. et Buk.)
Cytogenetic Characterization of Five "Criolla" Potato Genotypes, Solanum phureja (Juz. et Buk.)
Tiani Melissa Gómez Pulgarín1; Juan Bautista López Ortiz2; Rosana Pineda Tuirán3; Leonardo Fabio Galindo López4; Rafael Arango Isaza5 y Juan Gonzalo Morales Osorio6
1 Ingeniera Biológica. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias - Escuela de Biociencias. A.A. 3840, Medellín, Colombia. <tianisar@gmail.com>
2 Profesor Asociado. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias - Escuela de Biociencias. A.A. 3840, Medellín, Colombia. <jblopez@unal.edu.co>
3 Ingeniera Agrónoma. Investigadora. Universidad Nacional de Colombia - Sede Medellín - Unidad de Biotecnología Vegetal - Corporación para Investigaciones Biológicas - CIB-UNALMED. A.A. 7378, Medellín, Colombia.<rosanapaola@gmail.com>
4 Ingeniero Agrónomo. Investigador. Universidad Nacional de Colombia - Sede Medellín - Unidad de Biotecnología Vegetal - Corporación para Investigaciones Biológicas - CIB-UNALMED. A.A. 7378, Medellín, Colombia. <lefagalo@gmail.com>
5 Profesor Asociado, Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias - Escuela de Biociencias. A.A. 3840, Medellín, Colombia. <rearango@unal.edu.co>
6 Profesor Asociado. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias Agrarias. A.A. 1779, Medellín, Colombia. <jgmoraleso@unal.edu.co>
Recibido: Febrero 29 de 2012; aceptado: Mayo 07 de 2012.
Resumen. El cultivo de la papa ocupa el cuarto lugar en el mundo en importancia después del arroz, trigo y maíz. La papa criolla (Solanum phureja) posee características valiosas desde el punto de vista culinario y nutricional, por su alto contenido de vitaminas, minerales, fibra y calidad de proteína. Colombia es un centro de diversidad y origen de papas criollas, lo que representa un enorme potencial de recurso genético para el fitomejoramiento. En la Universidad Nacional de Colombia se conserva el banco de germoplasma de papas criollas de Colombia. El estudio de los cariotipos es importante para entender las bases genéticas que apoyan los cruces que se realizan en los programas de mejoramiento. A pesar de la importancia de este recurso, la caracterización de los cariotipos de las accesiones no ha sido realizada completamente. En este estudio se caracterizaron dos materiales de la colección de S. phureja y tres provenientes de la F1 del cruce de un genotipo resistente y otro susceptible a la enfermedad conocida como gota de la papa. El análisis comparativo permitió identificar diferencias en la morfología celular, dos niveles de ploidía (diploide y triploide), con dos variaciones aneugénicas (hipodiploide y trisomía doble) y cuatro números cromosómicos (18, 24, 26 y 36) para los cinco genotipos evaluados.
Palabras clave: Biodiversidad, cariotipos, idiogramas, tubérculos andinos.
Abstract. Potato crop is forth in importance worldwide after rice, wheat, and corn. Diploid potato cv. Criolla (Solanum phureja) shows highly desired culinary and nutritional characteristics, such as vitamins, minerals, fiber and protein quality. Colombia is a diversity and origin center of diploid potatoes cv. Criolla, which represents an enormous genetic resource potential for plant breeding. At Universidad Nacional de Colombia the potato "criollas" germplasm bank is mantained. Karyotype analysis is important for understanding genetic basis of plant breeding. Despite the importance of this resource karyotype analysis of this collection has not been completed. In this study two accesions of the S. phureja collection and three genotypes from a F1 obtained from a cross between a late blight disease resistant and a susceptible genotype were characterized. Comparative analysis allowed the identification of differences in the cellular morphology, two ploidy levels (diploid and triploid) with aneugenic variations (hypodiploidy and double trisomy), and four chromosome numbers (18, 24, 26 y 36) for the five genotypes evaluated.
Key words: Biodiversity, potato karyotype, idiograms, andean tubers.
El cultivo de la papa ocupa el tercer lugar en importancia en el mundo como alimento para consumo humano después del arroz y el trigo (International Potato Center, 2011). Las papas cultivadas están clasificadas en el género Solanum sección Petota. Estudios recientes proponen cuatro especies cultivadas y cerca de 100 especies silvestres relacionadas (Spooner et al., 2009, 2007). Algunas especies silvestres de papa poseen características genéticas agronómicamente deseables, como resistencia a enfermedades y a estrés abiótico entre otras, sin embargo, la transferencia de los caracteres deseables a las variedades cultivadas es limitada desde algunas especies silvestres. Los materiales haploides se han utilizado para poder transferir estas características deseadas desde el germoplasma silvestre hacia los materiales cultivados de papa. El conocimiento y el uso del número de balance de endospermo y gametos 2n y haploides se han usado en forma efectiva y eficiente para transferir la diversidad presente en las accesiones silvestres a las variedades cultivadas (Peloquin et al., 1989). El grupo de papas criollas diploides (2n=2x=24) Solanum tuberosum Grupo Phureja, denominada previamente S. phureja, posee excelentes propiedades agronómicas y cruzabilidad con otras especies silvestres y cultivadas de papa (Burgos et al., 2009). En Colombia se ha identificado un centro de diversidad de papas criollas, lo que representa un recurso genético invaluable para el mejoramiento de la papa cultivada, el cual no ha sido completamente caracterizado a nivel citogenético (Estrada, 1996). Para poder usar el recurso en todo su potencial es fundamental su caracterización citogenética.
El análisis citogenético de genotipos de papa se ha empleado para estudiar la producción de haploides y para usar haploides en genética y mejoramiento, para monitorear el número de cromosomas de los materiales híbridos y para investigar la inestabilidad cromosómica (Wilkinson, 1994; revisado por Gavrilenko, 2007). Todas las especies de la sección Petota presentan el mismo número básico de cromosomas (x=12). Rybin (1929, 1933) fue el primer autor que describió la serie completa de poliploides en papas silvestres (2x-3x-4x-5x-6x) y estableció una serie completa de poliploides en especies cultivadas (2x-3x-4x-5x). La taxonomía de las especies de papa cultivada ha presentado una enorme controversia; Rybin (1929), propuso usar las diferencias en los niveles de ploidía para la clasificación taxonómica de las papas cultivadas. El nivel de ploidía ha sido uno de los caracteres más importantes para reconocer las especies de papa cultivadas, indicando la necesidad de realizar este tipo de estudios en los recursos genéticos que poseen los bancos de germoplasma (Spooner et al., 2010; Gravilenko, 2007; Lee et al., 1972; Rybin 1929).
La posibilidad de obtener cruzamientos exitosos entre especies que difieren ampliamente en el nivel de ploidía, depende en gran medida del comportamiento meiótico de los híbridos resultantes. Una meiosis regular es esencial si la pérdida de material de cromatina o la fertilidad pobre, no es un obstáculo para la transferencia de material (Magoon et al., 1958). En cruces de materiales tetraploides X diploides que involucran S. phureja, se ha observado una frecuencia de alrededor del 90% de progenie tetraploide (Oppenheimer, 1933). Los estudios de meiosis y mitosis en polen de S. phureja, indican que esta alta frecuencia de descendencia tetraploide, posiblemente es causada por fertilización selectiva con granos de polen diploides no reducidos (Höglund, 1970).
Mohanty et al. (2004), examinaron el cariotipo de 30 genotipos de papa (S. tuberosum), encontrando características específicas de los cromosomas de los genotipos y alteraciones estructurales en los cromosomas con un raro hallazgo de condición aneusomática (2n=4xC2=50) en la variedad denominada K. Chandramukhi, indicando la importancia de estas indagaciones. Carputo (2003) estudió la citología y el comportamiento de los híbridos derivados del cruce 3x X 4x de S. commersonii - S. tuberosum. En esta investigación se encontraron híbridos con alta variación en el número cromosómico desde hipo-pentaploide (2n=5x-8=52), hasta hiper-pentaploide (2n=5x+7=67), con predominio de la clase de pentaploides euploides (2n=5x=60). Las observaciones de la megaesporogénesis y microesporogénesis en estos cruces generaron evidencia indirecta que indica que los híbridos pentaploides pueden producir gametos aneuploides viables con un número cromosómico variable entre 24 y 36. En este trabajo no se encontró un efecto del número cromosómico sobre los parámetros de fertilidad en los cruces de varios genotipos pentaploides (aneuploides) (Carputo, 2003). Estas caracterizaciones citogenéticas han sido particularmente importantes en el cultivo de la papa ya que, entre las plantas cultivadas, es una en la cual el nivel de ploidía, ha sido manipulado durante la reproducción sexual, como estrategia de mejoramiento. En este estudio se determinó la dotación cromosómica, el nivel de ploidía y la clasificación estructural de los cromosomas, de cinco genotipos de papa criolla, como apoyo al conocimiento y caracterización de este valioso recurso genético.
MATERIALES Y MÉTODOS
Para la caracterización citogenética se seleccionaron cinco genotipos de S. phureja, dos de los cuales (C17, C23) son variedades nativas (Figura 1); los otros tres (367, 385, 403), son clones F1, producto del cruce de los genotipos Col2 X Col3. Los materiales fueron mantenidos en el Centro Agropecuario Paysandú de la Universidad Nacional de Colombia, Sede Medellín (Universidad Nacional de Colombia, 1996).

Para establecer el número cromosómico, es decir el número total de cromosomas de cada genotipo, se realizaron extendidos de células de raíz en estado de mitosis y para determinar el grado de ploidía en la dotación cromosómica, o sea el número de veces que se repite cada tipo de cromosoma en el genoma, se realizaron extendidos celulares de anteras. Para verificar los resultados obtenidos se realizó un análisis comparativo de los cromosomas encontrados en las células en estado de mitosis y meiosis I, específicamente entre las fases de paquitene y metafase I (Gavrilenko, 2007; Howard, 1970; Höglund, 1970; Swaminathan y Howard, 1953; Estrada, 1996).
Preparación de los extendidos cromosómicos a partir de raíces y botones florales. El material vegetal se colectó en horas de la mañana, debido a que en este momento se observó el mayor número de células metafásicas (Talledo et al., 1993).
Botones florales. Una vez iniciada la floración, se colectaron botones florales de diferentes tamaños con el fin de evaluar el estado óptimo, para encontrar la mayor cantidad de células en división meiótica. Se determinó que cuando las anteras tenían una longitud desde su base de 0,4 cm, presentaron las mejores características para el análisis y se escogió este parámetro para el resto del trabajo. Los botones florales colectados se fijaron completos, inmediatamente en solución Carnoy (3:1, etanol absoluto: ácido ácetico glacial) y se conservaron a 4 °C hasta su uso. Los botones florales se abrieron con ayuda de estereomicroscopio y se separaron las anteras. Los microesporocitos contenidos en las anteras se extrajeron con estilete y se depositaron en una gota de colorante orceína-acética (1 g de orceína en 30 mL de ácido acético glacial, más 20 mL de agua destilada), en un portaobjetos. La preparación se flameó ligeramente y luego se aplastó con el cubreobjetos para realizar el extendido. La placa se observó en un microscopio de luz Nikon® E200.
Raíces. Se seleccionaron raíces debido a que son tejidos meristemáticos en constante división y presentan núcleos grandes lo que facilita la identificación de los cromosomas. Se evaluaron raíces de diferentes longitudes para determinar cuál presentaba el mayor número de células en estado de mitosis. Se seleccionaron raíces de 1,2 cm, como la longitud óptima para la observación de los diferentes genotipos. Las raíces colectadas se sometieron a pre-tratamiento con 8-hidroxiquinolina (0,002M), compuesto antimitótico que permite continuar la división celular hasta el estado de metafase. Luego se fijaron en solución Carnoy (3:1, etanol absoluto:ácido ácetico glacial), hasta su uso. Para preparar el extendido celular, se sumergieron las raíces fijadas en solución de coloración orceína-acética, se flameó ligeramente y se aplastó con cubreobjetos para la observación en un microscopio de luz, como se describió anteriormente. Los extendidos celulares de anteras y raíces se sellaron por los bordes del cubreobjetos con la resina sintética Entellán (Merck®), para su conservación (Estrada, 1996).
Cálculo de índice centromérico e índice braquial. Para clasificar los cromosomas se procedió a calcular los índices centromérico y braquial usando fórmulas existentes (Shaffer et al., 2009; Ferrer et al., 2009):
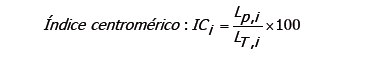
En donde Lp,i es la longitud del brazo corto del cromosoma i, y LT,i es la longitud total del cromosoma i.
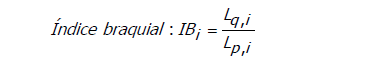
En donde Lq,i es la longitud del brazo largo del cromosoma i, y Lp,i es la longitud del brazo corto del cromosoma i.
Usando como parámetros los índices centroméricos y braquial, los cromosomas se clasificaron según el método propuesto por López y Márquez (2002) (Tabla 1).

El valor de la longitud total del genoma (TCL), equivale a la sumatoria de la longitud de los valores individuales, de todos los cromosomas de un genotipo. El área de cada célula se calculó a partir del diámetro, asumiendo una forma aproximadamente circular. Todas las mediciones se realizaron por triplicado y se calculó la desviación estándar.
Análisis comparativo de células entre los estados de mitosis y meiosis. Se estableció la relación del número de estructuras observadas en mitosis, respecto al número observado en la profase de la meiosis, para determinar el número de ploidía.
RESULTADOS Y DISCUSIÓN
Existen pocos trabajos para caracterizar citogenéticamente las papas silvestres y cultivadas, principalmente porque los cromosomas son pequeños y numerosos (Chen y Li, 2005; Wilkinson, 1994). Las especies incluidas en el género Solanum sect. Petota son difíciles de clasificar taxonómicamente en parte por la presencia de hibridación inter-específica tanto a nivel diploide como poliploide (Ovchinnikova et al., 2011; Ochoa, 1999; Hawkes 1990). En este estudio se evaluaron cinco genotipos de S. phureja, en los cuales se identificaron diversos niveles de ploidía.
Los ensayos preliminares para determinar el mejor momento de colecta de raíces y botones florales, mostraron que las raíces jóvenes de 1,2 cm presentaron el mayor número de células en mitosis. Para las anteras se determinó que cuando medían 0,4 cm, tenían el mayor número de células en meiosis. Tanto para anteras como para células de raíz, se observó un mejor comportamiento cuando se colectaron en horas de la mañana. También se determinó que un pre-tratamiento superior a cuatro horas no mejoró significativamente los resultados. Estos parámetros se utilizaron para todos los análisis realizados.
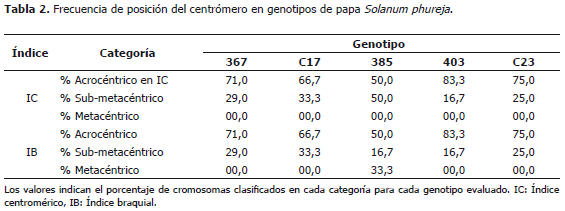
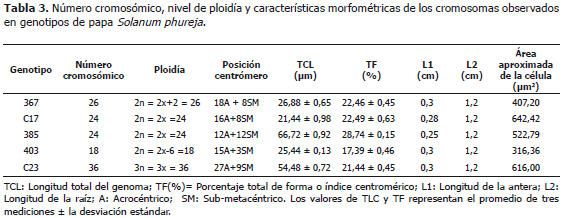
El grupo de genotipos de papas criollas S. tuberosum Grupo Phureja, se ha considerado con un nivel de ploidía diploide (2n=2x=24) (Hawkes, 1990). El análisis comparativo realizado en este estudio permitió identificar diferencias en la morfología celular, dos niveles de ploidía (diploide y triploide), con dos variaciones aneugénicas (hipodiploide y trisomía doble); cuatro números cromosómicos (18, 24, 26 y 36), en los cinco genotipos evaluados (Tablas 2 y 3; Figura 2). Los cruzamientos generados de forma natural o inducida y la segregación irregular de variedades como la triploide, posibilitan variaciones estructurales y numéricas que favorecen y/o desfavorecen la compatibilidad con otras especies, dando origen a genotipos triploides, originados por cruces entre materiales diploide (2x=24) y tetraploide (4x=48) y poliploidía impar por cruces entre diploides (2x=24) y triploides (3x=36). Algunos trabajos indican que es posible encontrar genotipos de papas que presentan raras aneuploidías. Mohanty et al. (2004), identificaron alteraciones estructurales en cromosomas, identificando un fenómeno raro de aneugénesis (2n=4xC2=50). Esta variación la atribuyeron a la no disyunción mitótica de los tallos, dando origen a raíces aneusomáticas.
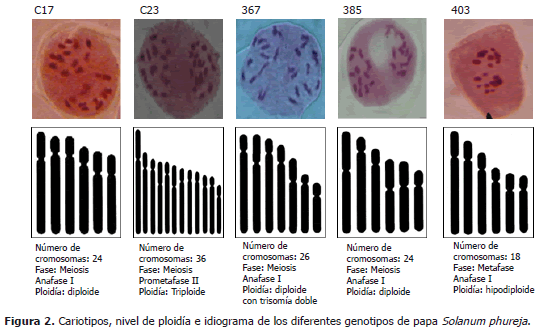
En este estudio se identificaron dos variaciones similares para el cruce mencionado (Col2 x Col3), en donde se observó una dotación cromosómica igual para todas las células; en el clon 403, se identificó una hipodiploidía y en el clon 367, se identificó una trisomía doble. La dotación cromosómica del clon 403 con 18 cromosomas, podría originarse por la unión de un gameto proveniente de un organismo diploide 2n = 24 y un gameto desbalanceado proveniente de una segregación dispar de un organismo hibrido segmental, este genera gametos irregulares que en este caso se puede suponer entre 6 hasta 18 cromosomas. La presencia de la trisomía doble para el genotipo 367, puede ser el resultado de una no disyunción somática de un organismo diploide, o la unión de dos gametos de genotipos diploides, que hayan sufrido una no disyunción doble en uno de sus parentales. La triploidía encontrada en la accesión C23, podría ser el resultado del cruce entre genotipos diploide (2n=24) y tetraploide (4n=48), unificando un material genético n con uno 2n para dar origen al triploide (3n=36).
Los resultados encontrados sugieren que la hibridación pudo haber contribuido en la formación de estos genotipos tanto a nivel diploide como poliploide. La introgresión e hibridación inter-específica natural, es común para muchas especies de la sección Petota y podría explicar estos resultados. Recientemente se ha encontrado evidencia que soporta el origen poliploide de los miembros de la sección Petota; estos trabajos se han realizado en especies silvestres y los resultados apoyan el origen auto y alopoliploide de diferentes especies de la sección Petota (Rodríguez et al., 2010). Respecto a las papas cultivadas, el origen híbrido de S. tuberosum se ha propuesto dentro de especies muy estrechamente relacionadas, excepto por dos de las tres accesiones examinadas de S. tuberosum grupo Chilotanum, las cuales aparentemente han sido hibridadas con la especie silvestre S. maglia (Rodríguez et al., 2010).
La generación de materiales aneuploides ha sido mencionada en varios trabajos. Lee et al. (1972), encontraron que cerca del 30% de la progenie de un cruce interespecífico 3x2x, resultaron aneuploides múltiples (2n=26-29). Seis de siete clones presentaron fertilidad femenina y la mayoría crecieron vigorosamente en el campo. Carputo (2003) mostró que los parámetros de fertilidad de los cruces que involucraron varios genotipos pentaploides (aneuploides), no fueron influidos por el número cromosómico. Este hallazgo sugiere un efecto de amortiguamiento de la poliploidía sobre la aneuploidía. En ese trabajo se indicó la posibilidad de usar exitosamente genotipos pentaploides (aneuploides) para el mejoramiento de la papa. Una situación similar se podría presentar en la colección de papas criollas evaluadas en este estudio, en donde la presencia de aneuploidías en la generación F1 del cruce Col2 x Col3, se podría usar en los programas de mejoramiento.
El índice centromérico entre los genotipos evaluados varió entre 17,39% y 28,74%, con una mediana de 26,88%; este índice varía entre 0% y 50% y en la medida que los valores se acercan al valor máximo mencionado, los cariotipos tienden a ser más simétricos (Ferrer et al., 2009). Para este caso el índice sugiere que los cromosomas tienden a tener su centrómero más cercano a uno de los extremos, en una proporción de 1:3 aproximadamente, indicando una tendencia a la asimetría con una mayor frecuencia de los cromosomas acrocéntricos, en la mayoría de los genotipos evaluados (Tabla 2). Basados en los cálculos realizados con los índices braquiales y centroméricos y la longitud relativa (TCL), se identificaron posibles homologías entre la mayoría de los cromosomas presentes en las distintas variedades, lo cual sugiere y confirma la hipótesis de un ancestro común entre ellas.
La longitud total del genoma combinado con índice centromérico o índice de asimetría, indica la cercanía existente entre los biotipos evaluados, mostrando las semejanzas entre los arreglos lineales de los genes de los cromosomas y permitiendo identificar homologías entre los cromosomas de una misma especie (García et al., 1996). El área celular hace parte de las características diferenciales entre los niveles de ploidía presentados por los genotipos de una especie, es un indicador que al ser correlacionado con otros parámetros citomorfológicos, facilitan los estudios de los genotipos como contribución a los trabajos de mejoramiento. El mejoramiento del cultivo de la papa se encuentra seriamente limitado por las barreras precigóticas y postcigóticas (Estrada, 1996). Los métodos usados para manipular los niveles de ploidía, hacen que la mayoría de especies de papa que poseen diferentes número de balance de endospermo, se puedan cruzar en forma compatible con S. tuberosum (Hougas y Peloquin, 1958). Para poder aplicar esos métodos es indispensable conocer el cariotipo de los materiales evaluados. Características como la fertilidad del polen y los niveles de ploidía y somías, son cruciales para determinar cuándo, un material se puede cruzar con otro con fines de mejoramiento. La fertilidad del polen es superior en la especie S. phureja, que en la especie S. tuberosum (Estrada, 1996). A pesar de los problemas de cruzabilidad que presentan las diferentes especies de papas silvestres y cultivadas, S. tuberosum se ha podido cruzar con numerosas especies silvestres generando un gran número de variedades cultivadas. La caracterización citogenética permite identificar algunos de los problemas de cruzabilidad, generados por variaciones en el número de cromosomas o el nivel de ploidía, contribuyendo de esta forma a diseñar las estrategias adecuadas para incorporar las características deseadas de las especies silvestres o genotipos nativos, en variedades comerciales. Este estudio contribuye a identificar estas posibles situaciones, para poder aprovechar este rico recurso genético que posee Colombia.
CONCLUSIONES
Para la colección de trabajo de papas criollas establecida en la Universidad Nacional de Colombia, se ha asumido incorrectamente que presenta un nivel diploide. Los resultados obtenidos muestran que la colección no tiene homogeneidad genotípica, identificándose materiales triploides, diploides y aneuploides, por lo que se requiere seguir con las valoraciones citogenéticas en el resto de genotipos mantenidos en la colección, haciendo énfasis en los parentales de los genotipos evaluados. La estandarización de la técnica de preparación de extendidos cromosómicos indica una longitud óptima de botón floral de 0,4 cm y raíz de 1,2 cm, para todos los genotipos evaluados. Los cruces entre genotipos de papa criolla pueden generar aneuploidías, que podrían ser materiales valiosos para los programas de mejoramiento del cultivo de la papa. La evaluación citogenética completa y complementada con evaluación molecular de todos los genotipos que se tienen en la colección, es importante para apoyar los programas de pre-mejoramiento y el secuenciamiento de genomas. Se muestra la gran importancia que tienen los estudios citogenéticos para la caracterización de toda variedad de plantas que entren en los diferentes procesos de producción agropecuaria.
AGRADECIMIENTOS
Este trabajo fue financiado por el Ministerio de Agricultura y Desarrollo Rural de Colombia, La Corporación para Investigaciones Biológicas y la Universidad Nacional de Colombia, Sede Medellín, Departamento de Ciencias Agronómicas y Escuela de Biociencias, con fondos del proyecto número-contrato 2008S71014-3625 "Estudio de la diversidad de papas silvestres (Solanum Sect. petota) asociadas a papas cultivadas (Solanum tuberosum), en los departamentos de Antioquia, Boyacá y Cundinamarca y análisis de su flujo de genes en condiciones de campo".
BIBLIOGRAFÍA
Burgos, G., E. Salas, W. Amoros, M. Auqui, L. Muñoa, M. Kimura and M. Bonierbale. 2009. Total and individual carotenoid profiles in Solanum phureja of cultivated potatoes: I. Concentrations and relationships as determined by spectrophotometry and HPLC. Journal of Food Composition and Analysis 22(6): 503-508. [ Links ]
Carputo, D. 2003. Cytological and breeding behavior of pentaploids derived from 3x × 4x crosses in potato. Theoretical and Applied Genetics 106(5): 883-888. [ Links ]
Chen, Q. and H. Li. 2005. An improved technique for high resolution mitotic chromosome studies in Solanum. HortScience 40(1): 54-56. [ Links ]
Estrada, N. 1996. Los recursos genéticos en el mejoramiento de la papa en los países andinos. pp. 1-14. En: Papas colombianas con el mejor entorno ambiental. UNIPAPA-ICA-CORPOICA. Comunicaciones y Asociados Ltda, Bogotá. 272 p. [ Links ]
Ferrer, H., N. Alcorcés y J. Méndez. 2009. Asimetrías cariotípicas observadas en dos especies de Gossypium L. Acta Biologica Paranaense, Curitiba 38(3-4):179-186. [ Links ]
García, D., M. González y O. Sam. 1996. Algunas características asociadas con la ploidía en papa. Cultivos Tropicales 17(1): 65-66. [ Links ]
Gavrilenko, T. 2007. Chapter 10: Potato cytogenetics. pp. 203-216. In: Vreugdenhil, D., J. Bradshaw, C. Gebhardt, F. Govers, D. Mackerron, M. Taylor and H. Ross. (eds.). Potato biology and biotechnology: advances and perspectives. First edition. Elsevier, UK. 823 p. [ Links ]
Hawkes, J.G., 1990, The potato: evolution, biodiversity and genetic resources. Belhaven Press, Oxford, UK. 259 p. [ Links ]
Höglund, M. 1970. Meiosis in Solanum phureja. Hereditas 66(2): 183-188. [ Links ]
Hougas, R. and S. Peloquin. 1958. The potential of potato haploids in breeding and genetic research. American Journal of Potato Research 35(10): 701-707. [ Links ]
Howard, H.M. 1970. Genetic of the potato Solanum tuberosum. Logos Press Limited, London. 126 p. [ Links ]
International Potato Center, 2011. http://www.cipotato.org/potato; consulta: mayo 2011. [ Links ]
Lee, H., R. Kessel and P. Rowe. 1972. Multiple aneuploids from interspecific crosses in Solanum: fertility and cytology. Canadian Journal of Genetics and Cytology 14(3): 553-543. [ Links ]
López, J.B. and M.E. Márquez. 2002. Modelo experimental para el estudio cromosómico en células de mamíferos. Universidad Nacional de Colombia, Medellín. 170 p. [ Links ]
Magoon, M., R. Hougas and D.C. Cooper. 1958. Cytogenetic studies of tetraploid hybrids is Solanum. Journal of Heredity 49(4): 170-178. [ Links ]
Mohanty, I.C., D. Mahapatra, S. Mohanty and A.B. Das. 2004. Karyotype analyses and studies on the nuclear DNA content in 30 genotypes of potato (Solanum tuberosum L.). Cell Biology International 28: 625-633. [ Links ]
Ochoa, C. 1999. The potatoes of South America: Perú. The wild species, Part 1. Allen Press, Lawrence, Kansas, USA. 1036 p. [ Links ]
Oppenheimer, C. 1933. Cytogenetische untersuchungen an bastarden knollentragender Solanum-Arten. I. Solanum chacoence Bitt. X Solanum tuberosum. L. S. str. - Z. Vererbungsl 65:72-98. [ Links ]
Ovchinnikova, A., E. Krylova, T. Gavrilenko, T. Smekalova, M. Zhuk, S. Knapp and D. Spooner. 2011. Taxonomy of cultivated potatoes (Solanum section Petota: Solanaceae). Botanical Journal of the Linnean Society 165: 107-155. [ Links ]
Peloquin, S., S. Jansky and G. Yerk. 1989. Potato cytogenetics and germplasm utilization. American Journal of Potato Research 66(10): 629-638. [ Links ]
Rodríguez, F., M. Ghislain, A. Clausen, S. Jansky and D. Spooner. 2010. Hybrid origins of cultivated potatoes. Theoretical and Applied Genetics 121(6): 1187-1198. [ Links ]
Rybin, V. 1929. Karyological investigation on some wild growing and indigenous cultivated potatoes of America. Trudy po Prikladnoj Botanike, Genetike i Selekcii (Bulletin of Applied Botany, Genetics, and Plant Breeding) 20: 655-720. [ Links ]
Rybin, V. 1933. Cytological investigation of the South American cultivated and wild potatoes, and its significance for plant breeding. Trudy po Prikladnoj Botanike, Genetike i Selekcii (Bulletin of Applied Botany, Genetics, and Plant Breeding) 2(2): 3-100. [ Links ]
Shaffer, L., M. Slovak and L. Campbell. 2009. ISCN 2009: An international system for human cytogenetic nomenclature: recommendations of the International Standing Committee on Human Cytogenetic Nomenc. First edition. Karger Publishing, Switzerland. 138 p. [ Links ]
Spooner, D., T. Gavrilenko, S. Jansky, A. Ovchinnikova, E. Krylova, S. Knapp and R. Simon. 2010. Ecogeography of ploidy variation in cultivated potato (Solanum sect. Petota). American Journal of Botany 97(12): 2049-60. [ Links ]
Spooner, D. M., M. Ames, D. Fajardo, and F. Rodríguez. 2009. Species boundaries and interrelationships of Solanum sect. Petota (wild and cultivated potatoes) are drastically altered as a result of PBI-funded In: Abstract, Botany and Mycology 2009 Annual Meeting. http://2009.botanyconference.org/engine/search/index.php. Snowbird, Utah, USA. [ Links ]
Swaminathan, M. and H. Howard. 1953. The cytology and genetics of the potato (Solanum tuberosum) and related species. Bibliographia Genetica 16:1-192. [ Links ]
Talledo, D., C. Escobar, and V. Alleman. 1993. Introducción al análisis cromosómico en vegetales. Universidad Ricardo Palma, Lima-Perú. 141 p. [ Links ]
Universidad Nacional de Colombia. 1996. Plan de Manejo Ambiental: centro de producción, docencia, investigación y proyección a la comunidad "Paysandú". Facultad de Ciencias Agropecuarias, Medellín. 52 p. [ Links ]
Wilkinson, M. 1994. Genome evolution in potatoes. pp. 43-70. In: Bradshaw, J.E and G.R. Mackay (eds.). Potato genetics. CAB International. Wallingford. UK. 576 p. [ Links ]

