










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.65 no.2 Medellín June/Dec. 2012
EFFECT OF NUMBER OF FUNCTIONAL LEAVES AT FLOWERING ON YIELD OF BANANA GRAND NAINE (Musa AAA Simmonds)
EFECTO DEL NÚMERO DE HOJAS FUNCIONALES A LA FLORACIÓN SOBRE LA PRODUCCIÓN DE BANANO GRAN ENANO (Musa AAA Simmonds)
Carolina Rodríguez González1; Daniel Gerardo Cayón Salinas2 y John Jairo Mira Castillo3
1 Agronomist Engineer. Universidad Nacional de Colombia - Sede Bogotá- Faculty of Agronomy. Carrera 45 No. 26-85, Bogotá, Colombia. <crodriguezg@unal.edu.co>
2 Associate Professor. Universidad Nacional de Colombia - Sede Bogotá - Faculty of Agronomy. Carrera 45 No. 26-85, Bogotá, Colombia. <dgcayons@unal.edu.co>
3 Head Centro de Investigaciones del Banano. Cenibanano-AUGURA. Conjunto Los almendros Km 4 vía Carepa - Apartadó. Carepa - Antioquia, Colombia. <jmira@augura.com.co>
Received: March 05, 2012; accepted: July 11, 2012.
Abstract. The effect of the selective removal of leaves at flowering on yield, growth, maturity and quality of Grand Nain banana fruits was evaluated. The present study was conducted in three regions of Urabá (Colombia) with precipitation low (experiment 1), medium (experiment 2) and high (experiment 3). The experimental design used randomized complete blocks, four treatments (6, 8, 10 and 12 leaves from the start of flowering) and three replications. Every 15 days, the thickness (cm) and length (cm) of central fruit from the second hand were measured and two fruits were taken to record fresh weight and dry weight. At harvest, leaf number, bunch weight (kg), and the thickness (cm) and length (cm) of central fruit from the second hand were measured. During fruit ripening (8 and 15 days), the contents of starch and sugars in the pulp were determined. The thickness and length of the fruit, bunch weight and the box-stem ratio were not affected (P<0.05) by selectively removing leaves. The percentages of total sugars and starch during fruit ripening were unchanged (P<0.05) as a result of the number of functional leaves. These results suggest that in banana cultivation, it is possible to leave less than 12 leaves at flowering without affecting fruit quality or postharvest ripening.
Key words: Leaf pruning, growth, postharvest, total sugars.
Resumen. Se evaluó el efecto de la eliminación selectiva de hojas a la floración sobre la producción, crecimiento, maduración y calidad de los frutos de banano Gran Enano. El estudio se realizó en tres zonas de Urabá (Colombia), con precipitación baja (experimento 1), media (experimento 2) y alta (experimento 3). Se usó un diseño de bloques completamente al azar, cuatro tratamientos (6, 8, 10 y 12 hojas desde el inicio de floración) y tres repeticiones. Cada 15 días se midieron el grosor (cm) y la longitud (cm) del fruto central de la segunda mano y se tomaron dos frutos más para registrar el peso fresco y el peso seco. A la cosecha se midieron el número de hojas, el peso del racimo (kg), el grosor (cm) y la longitud (cm) del fruto central de la segunda mano. Durante la maduración de los frutos (8 y 15 días) se determinaron los contenidos de almidón y azúcares en la pulpa. El grosor y la longitud del fruto, peso del racimo y la cantidad de cajas a partir de un racimo (ratio), no fueron afectados (P<0,05) por la eliminación selectiva de hojas. Los porcentajes de almidón y azúcares totales durante la maduración de los frutos no variaron (P<0,05) como consecuencia del número de hojas funcionales presentes. Estos resultados sugieren que en el cultivo de banano, es posible dejar las plantas con menos de 12 hojas desde la floración sin que se afecte la calidad de los frutos o su maduración en poscosecha.
Palabras clave: Defoliación, crecimiento, poscosecha, azúcares totales.
The growth and development of the banana bunch rely on the physiological activity of functional leaves that are present from the appearance of inflorescence at the apex of the pseudostem and during fruit development (Barrera et al., 2009). The maximum leaf area in the banana plant coincides with the emergence of inflorescence at the apex of the pseudostem since afterwards no new leaves are produced because the bunch is terminal and the area begins to decline as older leaves become senescent (Robinson and Guy, 2010; Turner et al., 2010). In the Urabá region, the banana plant emits 35 to 36 leaves during the vegetative phase at a frequency of 1 leaf/week during the rainy season and 0.4 to 0.6 leaves/week in drought conditions (Mira et al., 2004).
Studies have shown that in Musa, defoliation, depending on the timing and intensity, reduces yield and fruit quality (Ostmark, 1974; Stover, 1980; Satyanarayana, 1986). In this regard, Daniells et al. (1994) suggest the retention of 9-10 leaves from flowering to harvest, while Satyanarayana (1986) found that 12 leaves is the minimum required to achieve maximum yield in bananas. The banana plant can retain eight leaves from flowering without significantly affecting the weight of the bunch, but vigorous plants can obtain acceptable fruit even with only 2 and 4 leaves during the filling period (Robinson et al., 1992). In the plantain plant, selective removal of leaves affects leaf area and fruit growth development, the effect was more dramatic with more intense defoliation, and plants with 6 to 9 leaves from flowering produced clusters with greater weight compared with plants that only retained three functional leaves (Cayón, et al., 2000, 2002; Barrera et al., 2009). The 'Williams' banana showed a 35% increase in CO2 uptake in plants with eight leaves; this compensation is produced by the photosynthetic activity of leaves 2 to 7, which contribute most to the total plant photosynthesis during fruit development (Robinson et al., 1992). When there is loss of leaf area near filling organs, the plant regulates translocation and increases the demand of photoassimilates in the remaining leaves and induces increased photosynthetic activity in these leaves; and so, it is the type of leaf eliminated and not the intensity of defoliation that determines the effect of defoliation on bunch weight (Robinson et al., 1992; Arcila et al., 1995; Belalcázar et al., 1995).
There is some disagreement over the practice of selective pruning in banana leaves. Soto (1992) ensures that the constant emission of leaves during the vegetative phase, allows for defoliation to control pests or diseases. In subtropical areas, without the presence of Black Sigatoka, Robinson et al. (1992) recommend no defoliation of banana plants because it causes yield reduction, while Daniells et al. (1994) do recommend it because contributes to greater control of pests that attack the foliage. The aim of this study was to evaluate the effect of the number of leaves at flowering on the components of production and concentration of carbohydrates during fruit ripening.
MATERIALS AND METHODS
The study consisted of three experiments that were performed in the banana region of Urabá, located at 30 m, with precipitation of 2,896 mm/year, relative humidity 87%, temperature 27 °C and sunshine of 1,700 h/year. The experiments were established in a rainy season (July to November) in 3 plots planted with the Cavendish banana cv. Grand Nain: low precipitation (experiment 1, "Paraíso" farm, 2,323 mm/year), medium rainfall (experiment 2, "Arizona" farm, 2,560 mm/year) and high rainfall (experiment 3, "Porvenir" farm, 2,945 mm/year). In each experimental area, four leaf removal treatments were evaluated (6, 8, 10 and 12 leaves/plant) in randomly selected plants that were in early stages of flowering (inflorescence apical emission) and had 14 functional leaves, with the control being 12 leaves plant. We used a randomized complete design, 4 treatments, 3 replications and 15 plants/plot.
The bunches were bagged between 1 and 2 weeks after flowering with polyethylene bags (12.7 mm thick and impregnated with 1% chlorpyrifos). Throughout the period of bunch growth, the thickness (cm) and length (cm) of central fruit of the outer row of the second hand were measured every 15 days and two central fruits of the second hand were taken to record the fresh weight; was the pulp dried in a forced ventilation oven at 80 °C until constant weight. At harvest, 82 days after flowering (DAF), leaf number, bunch weight, thickness and length of central fruit from the outer row of the second hand and the box-stem ratio were measured in each plant. To analyze the effect of leaf removal on postharvest fruit ripening, in each treatment, eight segments were taken from 5-7 fruits of the second hand of the bunch and were packed in boxes for proper identification. The boxes were stored under controlled conditions (24 °C temperature and 50% relative humidity); subsequently, 8 and 15 days later, five fruits per treatment were taken to determine the concentration of starch (enzymatic hydrolysis) and total sugar (anthrone method) of dry pulp, in the Laboratorio de Agroempresas Rurales del Centro Internacional de Agricultura Tropical (Laboratory of Rural Agribusiness, International Center for Tropical Agriculture) CIAT. The generated data were subjected to analysis of variance, and for the mean comparison test, Tukey's multiple range was employed (P<0.05) using SAS® statistical software, and fruit growth curves were made, adjusted using a polynomial function of time.
RESULTS AND DISCUSSION
Fruit growth. In the three experiments (Figure 1), curves for dry matter accumulation during fruit development in all four treatments were fitted to a polynomial growth model with an initial logarithmic phase of slow growth (15 to 30 DAF) followed by a exponential stage of accelerated, constant growth (30 to 75 DAF) evidenced by the slope of the curve, and the final stage of senescence initiation (80 DAF) (Figure 1). The leaf removal treatments did not change this pattern of dry matter accumulation in fruits. The increased fruit weight from the emergence of inflorescence to commercial harvest is exponential (Turner, 1994), and the accumulation of dry matter in the bunch starts after flowering due to the supply of photoassimilates in the functional leaves and the mobilization of the greater part of the previously accumulated reserves in the corm (Chaves et al., 2009). Bunch growth is influenced by genetic factors that are critical due to their high influence on growth rate, final size and shape of the fruit (Salisbury and Ross, 2000; Azcón-Bieto and Talón, 2001). Some studies on completely defoliated plants have shown that carbohydrate reserves in the rhizome could support fruit growth in the absence of leaves, suggesting that there must be a temporary reserve of carbohydrates in the stem and rhizome that can support the development and maturation of fruits (Lapointe, 1998). In some species, the stem has been shown to act as a reservoir of carbohydrates (photoassimilates), but usually contributes less than 50% to fruit development (Wardlaw, 1990; Schnyder, 1993).
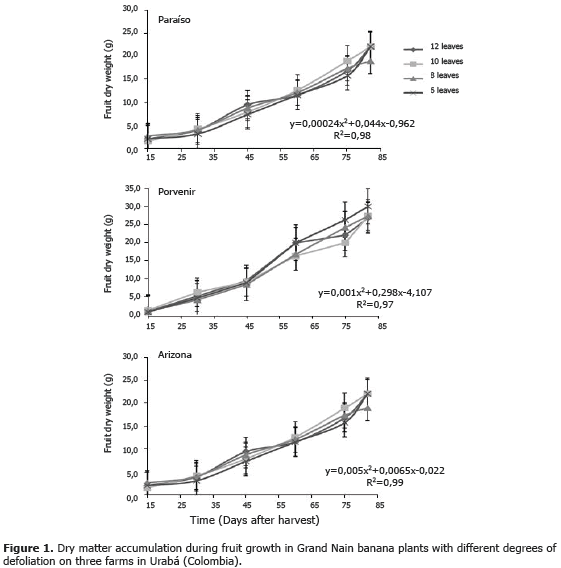
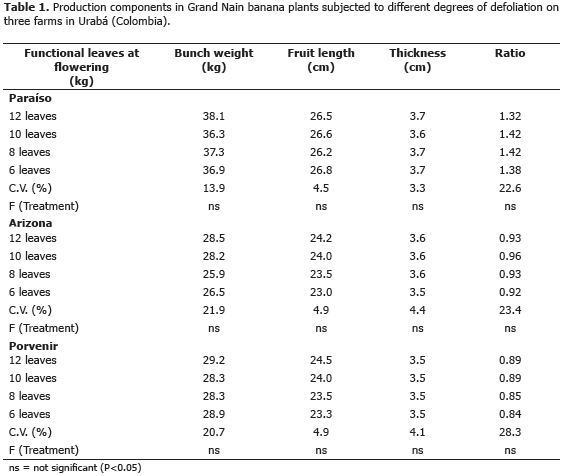
Bunch and fruit behavior. In all three experiments, yield components showed no statistical difference (P<0.05) between the leaf removal treatments (Table 1). Bunch weight, fruit thickness and length and box-stem ratio were not affected by the selective removal of leaves. Experiment 1 (Paraíso) yielded the highest bunch weight and Arizona the lowest; in Arizona, bunch weight was slightly lower in the six leaves treatment, but the length and size of the fruit and the ratio were not affected by the leaf removal. Dry matter accumulation in the fruit, and thus bunch weight, depends on photosynthesis of functional leaves present during the filling period, carbohydrates mobilized from the vegetative parts of the plant (Eckstein et al., 1995), and the limited photosynthesis in the peels of forming fruit (Jullien et al., 2001). Several authors have shown that banana leaf removal at flowering does not reduce bunch weight or fruit size (Vargas et al., 2008, 2009; Saborio and Granados, 2005; Segura and Valle, 2005) and does not induce changes in postharvest fruit maturation or quality (Ramirez et al., 2008), which suggests that this practice is recommended to improve production, fruit size and achieve greater control of pests of foliage. During the period from flowering to harvest, the banana plant can fill the fruit with a limited amount of leaves due to greater light penetration and diffusion of CO2 with air circulation and photosynthetic compensation, produced by young leaves (Robinson et al., 1992; Eckstein and Robinson, 1995; Delgado et al., 2003).
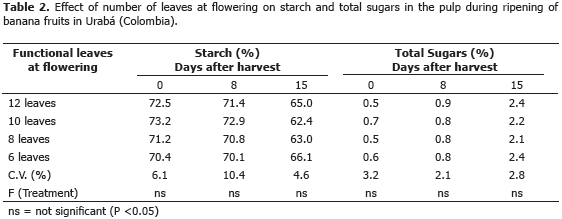
Carbohydrates. During fruit ripening under controlled postharvest conditions, the percentage of starch and total sugars in the pulp did not differ between defoliation treatments (Table 2). At harvest time, starch concentration was high (>70%) and decreased after 8 and 15 days, while total sugars had a marked increase over the previous days, but no significant differences between the defoliation treatments. Bananas and plantains are harvested at physiological ripeness (green) and during subsequent maturation starch decreases and total sugars increase, due to the biochemical activity during this metabolic process (Marriot et al., 1981; Hubbard et al., 1990; Arcila et al., 2002). The results of the present study are also consistent with those of Calvo and Bolaños (2001) in banana, and Barrera et al. (2009) in plantains, where the process of fruit ripening was not affected by the number of leaves at flowering, showing that the maturation of banana fruits, as indicated by Ramirez et al. (2008), is not affected by the selective removal of leaves at the time of flowering.
CONCLUSION
The results suggest that in cultured banana plants, less than 12 leaves may be left from flowering until harvest without affecting bunch weight, fruit quality or postharvest ripening.
ACKNOWLEDGEMENTS
Thanks to Centro de Investigaciones del Banano, Cenibanano - AUGURA for the technical and financial support provided for this project; and to the Arizona, Porvenir and Paraíso farms, for collaboration in the establishment and development of this study.
BIBLIOGRAPHY
Arcila, M.I., S. Belalcázar y G. Cayón, 1995. Influencia del número de hojas en postfloración sobre el llenado de los frutos del clon plátano Dominico-Hartón (Musa AAB Simmonds). pp. 90-93. En: Belalcázar, S., O. García, J.A. Valencia, M.I. Arcila, H. Mejía y H. García (eds.). Mejoramiento de la producción del cultivo del plátano. Produmedios, Bogotá. 256 p. [ Links ]
Arcila, M.I., L.D. Celis y H. Morales. 2002. Influencia de la edad de cosecha sobre la maduración del fruto de Dominico-Hartón en el departamento del Quindío. pp. 503-506. En: Memorias. XIV Reunión de la Asociación para la Cooperación en Investigaciones de Banano en el Caribe y América Tropical - Acorbat. Augura, Cartagena, Colombia. [ Links ]
Azcón-Bieto, J. y M. Talón. 2001. Fundamentos de Fisiología Vegetal. McGraw-Hill Interamericana, Barcelona, España. 522 p. [ Links ]
Barrera, J.L., G. Cayón, y J. Robles. 2009. Influencia de la exposición de las hojas y el epicarpio de frutos sobre el desarrollo y calidad del racimo de plátano Hartón (Musa AAB Simmonds). Agronomía Colombiana 27(1): 73-79. [ Links ]
Belalcázar, S., J.A. Valencia, M.I. Arcila y G. Cayón. 1995. Efecto de la defoliación selectiva durante la floración sobre el llenado de los frutos del clon de plátano Dominico-Hartón, Musa AAB Simmonds. pp. 104-111. En: Belalcázar, S., O. García, J.A. Valencia, M.I. Arcila, H. Mejía y H. García (eds.). Mejoramiento de la producción del cultivo del plátano. Produmedios, Bogotá. 256 p. [ Links ]
Calvo, C. y E. Bolaños. 2001. Comparación de tres métodos de deshoja en banano (Musa AAA): Su efecto sobre el combate de la Sigatoka Negra (Mycosphaerella fijiensis, Morelet) y sobre la calidad de la fruta. Corbana 27(54): 1-12. [ Links ]
Cayón, G., H. Morales y L.D. Celis. 2000. Respuestas del plátano Dominico-Hartón (Musa AAB Simmonds) a la defoliación selectiva en la zona cafetera central. Acta Agronómica 50(1,2): 20-25. [ Links ]
Cayón, G., H. Morales y L.D. Celis. 2002. Efecto de la defoliación sobre el comportamiento postcosecha del fruto de plátano Dominico-Hartón (Musa AAB Simmonds). Acta Agronómica 51 (1,2): 25-30. [ Links ]
Chaves, B., G. Cayón and J.W. Jones. 2009. Modeling potential plantain (Musa AAB Simmonds) potential yield. Agronomía Colombiana 27(3): 359-366. [ Links ]
Daniells, J.W., A.T. Lisle and N.J. Bryde. 1994. Effect of bunch trimming and leaf removal at flowering on maturity bronzing, yield and other aspects of fruit quality of bananas in North Queesland. Australian Journal of Experimental Agriculture 34(2): 259-265. [ Links ]
Delgado, E., O. González y N. Moreno. 2003. Efecto del desmane sobre el peso del racimo y las dimensiones del fruto del plátano FHIA 21. Bioagro 15(1): 17-22. [ Links ]
Eckstein, K. and J.C. Robinson. 1995. Physiological responses of banana (Musa AAA, Cavendish sub-group) in the subtropics. I. Influence of internal plant factors on gas exchange of banana leaves. Journal of Horticultural Science 70(1): 147-156. [ Links ]
Eckstein, K., J.C. Robinson and J. Davie, J. 1995. Physiological responses of banana (Musa AAA, Cavendish sub-group) in the subtropics. III. Gas exchanges, growth analysis and source-sink interaction over a complete crop cycle. Journal of Horticultural Science 70(1): 169-180. [ Links ]
Hubbard, N.L., D.M. Pharr and S. Huber. 1990. Role of sucrose phosphate synthase in sucrose biosynthesis in ripening bananas and its relationship to the respiratory climacteric. Plant Physiology 94: 201-208. [ Links ]
Jullien, A., N. Munier-Jolain, E. Malézieux, M. Chillet and B. Ney. 2001. Effect of pulp cell number and assimilate availability on dry matter accumulation rate in banana fruit [Musa sp. AAA group 'Grande Naine' (Cavendish subgroup)]. Annals of Botany 88: 321-330. [ Links ]
Lapointe, L. 1998. Fruit development in Trillium. Dependence on stem carbohydrate reserves. Plant Physiology 117: 183 -188. [ Links ]
Marriot, J., M. Robinson and K. Karikory. 1981. Starch and sugar transformation during the ripening of plantains and bananas. Journal of the Science of Food and Agriculture 32(10): 1021-1026. [ Links ]
Mira, J.J., A. Díaz y M Hernández. 2004. Influencia del régimen de lluvias sobre la productividad bananera de Urabá. p. 72. En: Memorias. XXXIV Congreso Sociedad Colombiana de Control de Malezas y Fisiología Vegetal. Comalfi, Bogotá [ Links ].
Ostmark, H.E. 1974. Economic insect pest of bananas. Annual Review of Entomology 19: 161-176. [ Links ]
Ramírez, M., M.V. Sáenz, A. Vargas and M. Araya. 2008. Leaf pruning intensities at flowering of banana (Musa AAA, cv. Grande Naine) did not influence fruit green and yellow life and quality. Scientia Horticulturae 115: 319-322. [ Links ]
Robinson, J.C., T. Anderson and K. Eckstein. 1992. The influence of functional leaf removal at flower emergence on components of yield and photosynthetic compensation in banana. Journal of Horticultural Science 67(3): 403-410. [ Links ]
Robinson, J.C., and V. Galán. 2010. Bananas and Plantains. Second edition. CABI, London, U.K. 311 p. [ Links ]
Saborío, G. y F. Granados. 2005. Defoliación controlada de plantas florecidas de banano (Musa AAA) como parte de una estrategia de manejo integrado de la Sigatoka Negra. p. 58. En: Resúmenes. Primer Congreso Científico Bananero Nacional. Dirección de Investigaciones, Corporación Bananera Nacional - Corbana. Guápiles, Costa Rica. [ Links ]
Salisbury, F.B. y C.W. Ross. 2000. Fisiología de las plantas. Editorial Paraninfo, Madrid. 988 p. [ Links ]
Satyanarayana, M. 1986. Effect of number of functional leaves on growth and yield of 'Dwarf Cavendish' banana (AAA). Banana Newsletter 9: 34-36. [ Links ]
Schnyder, H. 1993. The role of carbohydrate storage and redistribution in the source-sink relations of wheat and barley during grain filling: a review. New Phytologist 123(2): 233-245. [ Links ]
Segura, A. y H. Valle. 2005. Niveles de defoliación controlada a la floración y su efecto sobre el racimo y el hijo de sucesión de los cvs. Grande Naine, Williams y Valery. p. 56. En: Resúmenes. Primer Congreso Científico Bananero Nacional. Dirección de Investigaciones, Corporación Bananera Nacional - Corbana. Guápiles, Costa Rica. [ Links ]
Soto, M. 1992. Bananos: cultivo y comercialización. Segunda edición. LIL, San José, Costa Rica. 649 p. [ Links ]
Stover, R.H. 1980. Sigatoka leaf spots of banana and plantains. Plant Disease 64(8): 750-756. [ Links ]
Turner, D.W. 1994. Bananas and plantains. pp. 37-64. En: Schaffer, B. and P. Andersen (eds.). Handbook of enviromental physiology of fruit crops. Volume II: Sub-tropical and tropical crops. Taylor and Francis, Abingdon, U.K. 320 p. [ Links ]
Turner, D.W., J.A. Fortescue and D.S. Thomas. 2010. Chapter 2: Bananas and crop physiology. pp. 7-36. In: Damatta, F. (ed.). Ecophysiology of tropical tree crops. Nova Science Publishers, New York. 389 p. [ Links ]
Vargas, A., M. Guzmán, M. Araya, G. Murillo y F. Blanco. 2008. Efecto de la defoliación a la floración sobre el rendimiento del banano (Musa AAA) y la severidad de la sigatoka negra en condiciones semicomerciales. CORBANA 34(61): 39-54. [ Links ]
Vargas, A., M. Araya, M. Guzmán and G. Murillo. 2009. Effect of leaf pruning at flower emergence of banana plants (Musa AAA) on fruit yield and black Sigatoka (Mycosphaerella fijiensis) disease. International Journal of Pest Management 55(1): 19-25. [ Links ]
Wardlaw, I.F. 1990. The control of carbon partitioning in plants. New Phytologist 116: 341-381. [ Links ]
