










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.65 no.2 Medellín June/Dec. 2012
RELATIONSHIP OF Monalonion velezangeli CARVALHO & COSTA (HEMIPTERA: MIRIDAE) WITH THE PHENOLOGY OF AVOCADO (Persea americana MILL., CV. HASS)
RELACIÓN DE Monalonion velezangeli CARVALHO & COSTA (HEMIPTERA: MIRIDAE) CON LA FENOLOGÍA DEL AGUACATE (Persea americana MILL., CV. HASS)
Luisa Fernanda Torres Jaimes1; Guillermo Antonio Correa Londoño2; José Régulo Cartagena Valenzuela3; Danilo Augusto Monsalve García4 y Martha Eugenia Londoño Zuluaga5
1 Agronomic Engineer. Agrarian Science Master's student - Universidad Nacional de Colombia - Sede Medellin - Faculty of Agrarian Sciences - Department of Agronomic Sciences. P.O. Box 1779. Medellin, Colombia. <lftorresj@unal.edu.co>
2 Associate Professor. Universidad Nacional de Colombia - Sede Medellin - Faculty of Agrarian Sciences - Department of Agronomic Sciences. P.O. Box 1779. Medellin, Colombia. <gcorrea@unal.edu.co>
3 Associate Professor. Universidad Nacional de Colombia - Sede Medellin - Faculty of Agrarian Sciences - Department of Agronomic Sciences. P.O. Box 1779. Medellin, Colombia. <jrcartag@unal.edu.co>
4 Professional Researcher. CORPOICA C.I. El Nus. San José del Nus, San Roque, Antioquia, Colombia.<dmonsalveg@corpoica.org.co>
5 Principal Researcher. CORPOICA C. I. La Selva. P.O. Box 100. Rionegro, Antioquia, Colombia. <mlondono@corpoica.org.co>
Received: January 19, 2012; accepted: September 19, 2012.
Abstract. Monalonion velezangeli is considered one of the most harmful pests of avocado cv. Hass, as far as it attacks vegetative buds, flowers and fruits during all the productive stages of the crop. This situation is aggravated by the lack of knowledge on insect preferences and their relationship with crop phenology. As a contribution to the management of this insect, we studied the relationship between plant phenology and the presence of M. velezangeli, as well as its preferences across tree strata and structures. Data were obtained from six orchards located in the Colombian departments of Antioquia, Caldas and Quindío. After randomly selecting 20 trees at each orchard, 12 branches were marked on each tree, covering the combinations of the four cardinal points with three plant strata (low, medium and high). During a period of 12 months, each branch was monitored by recording its phenological stage (vegetative, flowering and fruiting), the number of individuals of M. velezangeli and the presence of fresh damages on it. An analysis of variance determined that the presence of the insect and its damage were more abundant on the flowering and fruiting branches than on the vegetative ones, as well as in the medium and upper strata of the trees. The damage caused by M. velezangeli was observed to be significantly higher in the reproductive structures (flowers and fruits) as compared to the vegetative structures.
Key words: Tropical fruits, tree canopy, avocado bug, phenological stage.
Resumen. Monalonion velezangeli es considerada una de las principales plagas del aguacate cv. Hass en Colombia, debido a que ataca brotes vegetativos, flores y frutos, durante todas las etapas productivas del cultivo. Esta situación se ve agravada por el desconocimiento de las preferencias del insecto, así como de su relación con la fenología de la planta. Como un aporte al manejo de este insecto, se estudió la relación entre la fenología de la planta y la presencia de Monalonion, así como sus preferencias por estructura y estrato. Las observaciones se hicieron en seis huertos ubicados en los departamentos de Antioquia, Caldas y Quindío (Colombia). Se eligieron 20 árboles por finca; en cada uno, se marcaron 12 ramas que correspondían a las combinaciones de los cuatro puntos cardinales en tres estratos (bajo, medio y alto). En la rama seleccionada se registró, durante 12 meses, su estado fenológico (vegetativo, floración o fructificación), el número de individuos de M. velezangeli y los daños frescos presentes. Mediante análisis de varianza, se determinó que la presencia del insecto, así como su daño fueron mayores sobre las ramas en floración y fructificación y en los estratos alto y medio de los árboles. El daño causado por M. velezangeli fue significativamente mayor en las estructuras reproductivas (flores y frutos) en relación con las estructuras vegetativas.
Palabras clave: Frutas tropicales, dosel del árbol, chinche del aguacate, estado fenológico.
In Colombia, avocado (Persea americana Mill.) cv. Hass planted area is approximately 5,500 hectares, distributed in the departments of Antioquia, Tolima, Cauca, Quindío, Caldas, Valle del Cauca and Santander, which yielded 13,000 fruit t in 2009 (Mejía, 2010). Avocado orchards can be found between 1,200 and 2,600 masl, thus exhibiting a wide adaption range and correspondingly variable developmental behaviors (Vásquez et al., 2010).
Understanding of the crop phenology not only contributes to the development of strategies for pest and disease prevention and management, but also allows knowing the production limiting factors as well (Whiley and Saranah, 1995). In Colombia, there are different insects affecting the production of this crop among which we can count the complex of fruit and stem suckers and borers (Londoño, 2008).
One of the main pests affecting avocado cv. Hass in Colombia is the avocado bug, Monalonion velezangeli Carvalho & Costa, which affects the internal and external quality of the fruit and is capable of determining losses that range from 50 to 80% of production (Arango and Arroyave, 1991). M. velezangeli belongs to the order Hemiptera, family Myridae. In Colombia, the record of crops affected by this pest dates from 1984. Adults and nymphae harm the tree by feeding on it with their sucker-like apparatus, mainly attacking young shoots and small fruits (Arango and Arroyave, 1991).
Fresh injuries caused by the insect on leaves, stems and inflorescences are featured by brown reaction areas with red, humid and cumbersome exudates (Vargas and Londoño, 2009); whereas old injuries look dry, dark and furrowed. The fruits manifest irregular brown stains of oily appearance. Under the attack of the plague, young shoots crack and wither, whereas branches might dry out and break. Injured fruits present white stains formed by plant exudates. Small fruits just stop growing and dry out (Londoño, 2008).
Neither the preferences of M. velezangeli over the plant nor the relationship between the insect and the phenology of the crop are well known in Colombia to date. Thus, the objectives of the current research work were to determine the preferences of the insect in terms of structure and strata over the avocado cv. Hass plant, as well the relationship between the phenology of the crop and the presence of the plague or the injuries it causes.
MATERIALS AND METHODS
Space and time location. Research was conducted from September 2010 to August 2011, in six avocado cv. Hass orchards where the plague had been reported (see Table 1 for site location and description).
Sampling. The active population of M. velezangeli, as well as the recent damages it produced (brown reaction areas with red, bulky exudates of humid texture), were recorded in the six studied orchards approximately every 20 days. Each of the orchards located in East Antioquia-Colombia (El Cebadero, El Guarango, La Miranda and Persea) were the object of 11 samplings; while those found in the Coffee Growing Zone (El Jordán and Chile) were assessed 14 times each, for a great total of 72 samplings.
Twenty trees ranging from four to eight years old were randomly selected at each orchard. Twelve terminal branches were marked in each tree, so as to cover the four cardinal points across three strata: low, medium and high. At each sampling, the phenological stage of the branch was recorded (vegetative, flowering or fruiting) together with the number of nymphs and adults and the number of recent injuries found in the last 30 cm of the branch. The response variable was the presence of the plague, expressed as the amount of M. velezangeli individuals and recent injuries found at each sampling. The structure on which the injuries were found was also recorded, as well as its available area, which was used as covariate in order to estimate possible preferences of the plague. In this case, the response variable corresponded to recent injuries. The phenological condition of the branches was also used to observe variations in the biological processes of avocado cv. Hass along the year.
Average surface estimation in the evaluated plant structures. The surfaces of the different available food sources were calculated through the cylinder's formula, in the case of branches and inflorescences, and through that of the prolate spheroid for floral buds (as parts of the inflorescence) and fruits, in which the equatorial diameter is shorter than the polar one.
Diameters were measured with an IPS 65® digital caliper. Structure lengths were estimated with a L525CME Lufkin® flexometer, and foliar area with a LI-3000A LI-COR® meter.
Statistical analysis. Data were worked out through ANOVA on a SAS® software package licensed to Universidad Nacional de Colombia, Sede Medellin. Mean comparisons were carried out through a Tukey test for a=0.05. Each farm at a given time was included as a blocking factor. Phenological stage, tree stratum and attacked tree structure were taken as the fixed effects of the model, in which the available surface of each structure was taken as covariate. The transformation was applied for the response variable (the sum of insects and/or recent injuries).
RESULTS AND DISCUSSION
Relationship between the presence of M. velezangeli and the crop's phenological stage. The statistical analysis revealed that the insect was significantly more abundant in fruiting and flowering branches than in vegetative ones. No significant differences were found between the two former branch types (Table 2).

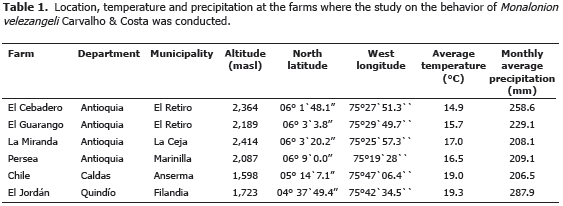
Presence of the insect across tree strata. Statistically significant differences were found between the three strata, the upper and medium ones registering a more abundant presence of the insect (Figure 1).
Insect preferences for tree structures. The damage caused to fruits and inflorescences was greater than that found on vegetative structures (P = 0.0006). No significant difference was observed between fruit and inflorescence damage (P = 0.8886; Table 3).

In a similar study with greenhouse grown, one year old avocado trees, Vargas and Londoño (2009) found the vegetative structures (76.8% of feeding sites) to be the preferred ones by M. velezangeli, followed by inflorescences (13.1%). These results contrast with those of the current work, in which the insect showed preference for reproductive structures (inflorescences and fruits). Ramírez et al. (2008) mention that the adults of this plague feed on the flower buds of the coffee plant, as also observed in the present study with avocado trees, and documented by Vargas and Londoño in 2009.
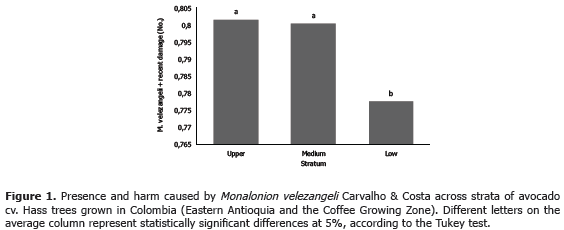
In the two regions sampled in the current work, the avocado trees showed heterogeneous phenology, i.e., they presented vegetative, flowering and fruiting branches all along the studied period. Branches carrying fruits at different developmental stages were relatively constant over time, while vegetative and flowering branches were inversely correlated (Figures 2 and 3). Under these conditions, the insect has a multiple offer of all available structures, which allows it a more independent choice on feeding sites. Recurrent vegetative shooting in avocado cv. Hass is interpreted by Cossio et al. (2008) as a means to abundant flowering, which in turn increases the probability of fruiting, thus minimizing the alternation produced by the lack of fruiting sites. Although this cultivar originated in Guatemala, it incorporates some Mexican genes (Newett et al., 2002), which determine the terminal buds to develop at anthesis (Rodríguez, 1982), and accounts for the heterogeneity implied in the simultaneous presence of vegetative and flowering branches.
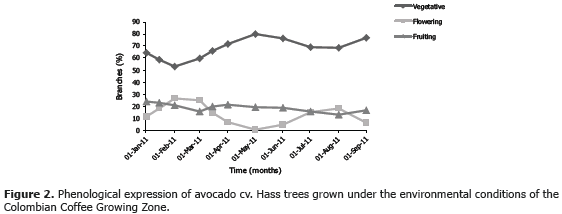
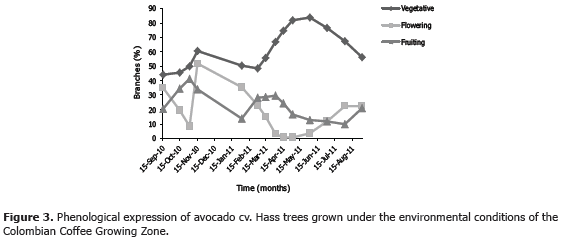
On the other hand, the time elapsed from floral initiation to anthesis is variable and depends on major climatic conditions during floral development (Salazar et al., 1998). In this respect, Gaillard (1987) affirms that the flowering period is considerably variable, depending on cultivars and climate conditions, and normally taking place during the dry season, which is when the temperature is low. Chaikiattiyos et al. (1994) have confirmed this, stating indeed that floral induction or initiation takes place approximately two months before flowering and requires temperatures below 25 °C because the transition from the vegetative to the flowering phase generally takes place between 15 and 25 °C. Tropical zones have particular climate conditions that contrast with those of temperate regions. The former are featured by sharp contrasts between day and night, in turn affected by solar radiation. In the Andean regions temperature ranges sometimes reach up to 20 °C (Jaramillo, 2005). So, it is not surprise that Avilan et al. (2007) have found three to five vegetative shootings in Venezuelan avocado trees in one year, which certainly favors phenological heterogeneity. The irregular phenology of avocado in tropical regions allows the presence of M. velezangeli all along the year, since this plague is prone to attack any branch type.
The results indicate that this plague causes more harm on the flowering and fruiting branches of avocado cv. Hass, specifically attacking reproductive structures. Thus, by affecting fruit quality, it takes a toll directly on productivity, and by impacting inflorescences it hampers future production.
Other research works coincide in reporting that certain bed bugs attacking avocado trees mainly feed on fruits and inflorescences, just as M. velezangeli. Alberts (2010) points out that the mirid complex of the genus Lygus spp., which affects avocado plantations in the district of Soutpansberg (North of South Africa), is more active when there are more open flowers. Similarly, Yarita (2005) indicates that the harm produced by the mirid Dagbertus sp. on avocado crops in Virú, La Libertad (Peru), takes place mainly during the feeding stage, having floral buds and recently set fruits fall down and significantly reducing productivity. Glenn and Baranowski (unpublished data cited by Peña et al. 2003) have observed that the mirid species Dagbertus fasciatus (Reuter) and D. (Reuter) feed on buds, leaves, flowers and small fruits in avocado crops of in Florida (USA), mainly affecting recently formed fruits and flowers and causing them to fall down. Peña, et al. (2003) confirm that the mirids D. fasciatus (Reuter), Rhinacloa sp. and D. olivacea (Reuter) feed on avocado flowers, thus contributing to their falling down and to significant harvest reductions in the Florida orchards (USA).
The dominant presence of M. velezangeli in the medium and upper strata can be considered to be an antecedent of specifically oriented monitoring and management techniques. It is worthwhile noting that, for this reason, high trees are likely to allow the plague in distant places, out of the reach of management practices. Hence, it might be advisable to keep relatively low trees at the moment of pruning.
CONCLUSION
The results indicate that the presence of M. velezangeli and the harm it causes on avocado cv. Hass have a greater impact on flowering and fruiting branches at the medium and upper strata of the trees, the most affected structures being flowers and fruits. The heterogeneous phenology of avocado favors the presence of the plague all along the year.
ACKNOWLEDGEMENTS
The authors want to thank the Ministry of Agriculture and Rural Development of Colombia (Ministerio de Agricultura and Desarrollo Rural de Colombia - MADR), the Colombian Corporation for Agricultural Research (Corporación Colombiana de Investigación Agropecuaria - CORPOICA) and its associates, who supported the current study through the project "Development of strategies for the prevention and management of Monalonion velezangeli Carvalho & Costa" (Desarrollo de estrategias para la prevención and manejo de Monalonion velezangeli Carvalho & Costa). Just as well, we are thankful to the avocado growers who allowed the conduction of research activity in their farms.
BIBLIOGRAPHY
Alberts, A.J. 2010. Defining economical action levels for avocado bugs (Miridae) and the use of area wide IPMK to warn growers of possible outbreaks. South African Avocado Growers Association Yearbook 33: 63-69. [ Links ]
Arango, R. y H. Arroyave. 1991. Ciclo de vida y hábitos de la chinche del aguacate Monalonion velezangeli Carvalho y Costa (Hemiptera: Miridae), en Antioquia. Tesis de grado Ingeniero Agrónomo. Universidad Nacional de Colombia, Sede Medellín, Medellín. 52 p. [ Links ]
Avilán, L., E. Soto, M. Pérez, M. Rodríguez y J. Ruiz. 2007. Fenología de cultivares e híbridos de aguacate de la raza mexicana en la región centro-norte costera de Venezuela. Agronomía Tropical 57(2): 89-98. [ Links ]
Cossio, L.E., S. Salazar, I.J. Durán y R. Medina. 2008. Fenología del aguacate Hass en el clima semicálido de Nayarit, México. Revista Chapingo Serie Horticultura 14(3): 319-324. [ Links ]
Chaikiattiyos, S., C. Menzel and T. Rasmussen. 1994. Floral induction in tropical fruit trees: Effects of temperature and water supply. Journal of Horticultural Science 69(3): 397-415. [ Links ]
Gaillard. J. 1987. Lavocatier sa culture, ses produits. (Techniques agricoles et productions tropicales). France. G.P. Maisonneuve et Larose et A.C.C.T. 419 p. [ Links ]
Jaramillo, R. 2005. Clima andino y café en Colombia. Chinchiná, Cenicafé. 196 p. [ Links ]
Londoño, M. 2008. Capítulo IV: Insectos. pp. 135-136. En: Tecnología para el cultivo del aguacate. Corporación Colombiana de Investigación Agropecuaria. CORPOICA, C.I. "La Selva", Rionegro, Antioquia, Colombia. Manual Técnico No. 5. 241 p. [ Links ]
Mejía, A. 2010. Contexto productivo actual del aguacate en Colombia. p. 32-35. En: Memorias. VII Seminario de Frutas Tropicales. Secretaría Técnica Consejo Nacional del Aguacate. Ministerio de Agricultura y Desarrollo Rural. Medellín. [ Links ]
Newett, S.D., J.H. Crane and C.F. Balerdi. 2002. Cultivars and rootstocks. 161-187. In: Whiley, A.W., B. Schaffer and B.N. Wolstenholme (ed.). The avocado botany, production and uses. CABI Publishing. New York. 416 p. [ Links ]
Peña, J., H. Glenn and M. Baranowski. 2003. Dynamics and sampling of mirids (Hemiptera: Miridae) in avocado in Florida. p. 495-501. In: Proceedings. V World Avocado Congress. Granada - Malaga, España. [ Links ]
Ramírez, H., Z. Gil, P. Benavides y A. Bustillo. 2008. Monalonion velezangeli la chinche de la chamusquina del café. Avances Técnicos Cenicafé No 367. Chinchiná, Caldas, Colombia. [ Links ]
Rodríguez, F. 1982. El Aguacate. AGT (ed.). México. 167 p. [ Links ]
Salazar, G., E.M. Lord and C.J. Lovatt. 1998. Inflorescence and flower development of the "Hass" avocado (Persea americana Mill) during "on" and "off" crop years. Journal of the American Society for Horticultural Science 123(4): 537-544. [ Links ]
Vargas, H. y M. Londoño. 2009. Descripción del daño ocasionado por Monalonion velezangeli Carvalho y Costa (Hemiptera: Miridae) sobre Persea americana Miller. En: Resúmenes XXXVI Congreso de la Sociedad Colombiana de Entomología - SOCOLEN, Medellín. 94 p. [ Links ]
Vázquez, L., G. Gallego, M. Londoño y M. Torres. 2010. Aspectos biofísicos y socioeconómicos del sistema de producción de aguacate Hass en Antioquia y Eje Cafetero. En: Memorias. VII Seminario de Frutas Tropicales. Secretaría Técnica Consejo Nacional del Aguacate. Ministerio de Agricultura y Desarrollo Rural, Medellin. 67 p. [ Links ]
Whiley, A. and B. Saranah. 1995. Pheno-physiological modelling in avocado - an aid in research planning. pp. 71-75. In: Proceedings. III World Avocado Congress, Tel Aviv, Israel. [ Links ]
Yarita, Y. 2005. Ciclo biológico y algunos aspectos del comportamiento de Dagbertus sp. (Hem.: Miridae) en el cultivo de palto en el fundo Frusol, Virú La Libertad, agosto, 2004 a marzo, 2005. In: http://www.avocadosource.com/international/peru_papers/YaritaYulissa2005.pdf [ Links ]
