










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.65 no.2 Medellín June/Dec. 2012
POPULATION STRUCTURE AND DEMOGRAPHY OF THE PALM Wettinia kalbreyeri FROM AN ANDEAN MONTANE FOREST OF COLOMBIA
ESTRUCTURA POBLACIONAL Y DEMOGRAFÍA DE LA PALMA MACANA Wettinia kalbreyeri EN UN BOSQUE ALTOANDINO DE COLOMBIA
Carlos Esteban Lara Vásquez1; María Claudia Díez Gómez2 and Flavio Humberto Moreno Hurtado3
1 Forestry Engineer. Teacher Assistant Universidad Nacional de Colombia - Sede Medellín - Faculty of Agricultural Sciences - Deparment of Forestry Siences. P.O. Box. 579. Medellín, Colombia. <celarav@unal.edu.co>
2 Associate Professor. Universidad Nacional de Colombia - Sede Medellín - Faculty of Agricultural Sciences - Deparment of Forestry Siences P.O. Box. 1779, Medellín, Colombia <mcdiez@unal.edu.co>
3 Associate Professor. Universidad Nacional de Colombia - Sede Medellín - Faculty of Agricultural Sciences - Deparment of Forestry Siences P.O. Box. 1779. Medellín, Colombia. <fhmoreno@unal.edu.co>
Received: February 28, 2012; accepted: September 17, 2012.
Abstract. Wettinia kalbreyeri is a monoecious palm distributed on the Andean highland forests from Colombia and Ecuador with high ecological and economical value. For several decades, its populations have been intensively harvested, which caused a significant decrease of its natural stocks. However, no research has been done yet on the regeneration and dynamics of this important species. In order to get critical information for its management, we characterized the life cycle and surveyed for one year an undisturbed natural forest dominated by this palm in the Western Cordillera of Colombia. We established 10 permanent plots (0.1- ha each) and evaluated the structure and population dynamics with a matrix model structured by sizes. We found a high density of individuals up to 50 cm- height (129.520 ± 72.701 ha-1); the density of adult palms was also high (768 ± 263 ha-1), with an average basal area of 21.34 ± 8.84 m2 ha-1. Population growth was positive during the period evaluated (l = 1.079); results of the elasticity analysis suggest that changes of adult density could severely impact the population dynamics. Because of the high temporal variability of natural populations, a longer monitoring time is important to improve the reliability of estimates.
Key words: Elasticity, population dynamics, sustainable management, tropical Andes.
Resumen. Wettinia kalbreyeri es una palma monoica de los bosques altoandinos de Colombia y Ecuador, con un alto valor ecológico y económico. Durante varias décadas sus poblaciones han sido aprovechadas intensivamente, lo cual generó una notable disminución de sus existencias naturales. Sin embargo, hasta ahora no se ha realizado ningún trabajo sobre la regeneración y dinámica de esta importante especie. Con el fin de acopiar información fundamental para su manejo, se caracterizó su ciclo de vida y se monitoreó durante un año un bosque natural no intervenido dominado por esta palma en la cordillera occidental de Colombia. Para ello se establecieron 10 parcelas permanentes de crecimiento de 0,1 ha y se evaluó la estructura y dinámica poblacional por medio de un modelo matricial estructurado por categorías. Sobresale la alta densidad de individuos hasta 50 cm de altura (129,520 ± 72,701 ha-1). La densidad de individuos adultos también fue alta (768 ± 263), con un área basal promedia de 21,34 ± 8,84 m2 ha-1. El crecimiento poblacional positivo durante el período evaluado (l = 1,079); los resultados del análisis de elasticidad sugieren que cambios en la densidad de los individuos adultos podrían tener un alto impacto sobre la dinámica de la población. Debido a la alta variabilidad temporal de la dinámica de las poblaciones naturales, es importante su monitoreo durante un período más prolongado, lo cual permitiría mejorar la confiabilidad de las estimaciones.
Palabras clave: Dinámica poblacional, elasticidad, manejo sostenible, Andes tropicales.
Palms are an important component of neotropical forests, not only for their high abundance and wide altitudinal distribution (Durán and Franco, 1992; Henderson et al., 1995), but also for their ecological and economical functions. For instance, as a source of food for wild and pollinators; in addition, palms are often one of the most important sources of food for several species along dry seasons (Durán and Franco, 1992). On the other hand, people have used the Areaceae family in different ways; in Colombia, about 120 different applications of this family have been identified and 61% of 231 palm species have one or more alternative products (Galeano and Bernal, 2010).
Particularly, Wettinia kalbreyeri (Burret) R. Bernal is a palm species distributed in the highlands of Colombia and Ecuador (above 2000 m altitude). Despite its logging is forbidden, many people in Antioquia and in other states of Colombia have been exploiting this palm for several decades; basically, they cut the palm stems to be used in rural constructions and handicrafts (CORANTIOQUIA, 2007). Because of the high demand, the extraction of this palm is currently increasing in the Andean forests of this region of Colombia where it is abundant (in fact, several palm species of this region are endangered; Galeano and Bernal, 2010). This situation seems to be non-compatible with the long-term conservation of this species.
Despite its tremendous importance and the imminent danger of this species, the basic knowledge for its sustainable management is lacking. For example, there are no studies on the ecology and biology of W. kalbreyeri, including basic aspects of its population dynamics and demography. Fortunately, several researches of population dynamics have been done in the tropics using another palm species as a model of study; particularly, in South America several authors have studied other palm species (Durán and Franco, 1992; Olmsted and Álvarez, 1995; Zea, 1997; Bernal, 1998; Bernacci, 2001; Suárez, 2001; Waldrón, 2001; Bonesso et al., 2008; Holm et al., 2008; Thompson et al., 2009; Arango et al., 2010; De Cássia et al., 2010). These studies provide insights into the biological systems and present valuable information to determine levels of sustainable harvest (Svenning and Balslev, 1997). These studies were based on matrix models, which classify the population in different categories of sizes (Caswell, 2001) and have proven to be a solid approach to describe the complex life cycle of palms (Silvertown et al., 1996).
This study is the first step for understanding the ecology of W. kalbreyeri, and includes a description of its life cycle and habitat. Particularly, we focused on the (1) population structure and (2) population dynamics based on a matrix model of W. kalbreyeri structured by sizes (Caswell, 2001) along one year using permanent plots in an Andean forest from the north portion of the Western Cordillera of Colombia.
METHODS
Study area. This study was conducted in the regional reserve named "Cuchilla Jardín-Támesis", located in the north of the Western Cordillera of the Colombian Andes (5°37'45" - 5°39'18" N and 75°47'53" - 75°46'30" W). The total area of the reserve is about 32,100 ha, distributed in two municipalities of the department of Antioquia (Jardín and Támesis). Specifically, the survey was carried out in natural forests of two adjacent properties named "La Tribuna" and "La Glorieta", which have an approximate area of 850 ha, and belong to the Corporación Autónoma Regional del Centro de Antioquia (CORANTIOQUIA), which is the regional environmental authority. The altitude of the study site varies between 2,200 and 2,800 masl. Average rainfall is 3,000 mm (all months rainfall is above 100 mm), temperature is 17 ºC, relative humidity is 81%, and sunshine reaches 1987 h per year (CORANTIOQUIA, 2007). The main life zone in this area is Tropical Lower Montane Moist Forest (sensu Holdridge, 1978). The studied forests are dominated by W. kalbreyeri; although some tree species share the canopy of these forests, such as Billia rosea, Chrysochlamys colombiana, Dicksonia sellowiana, Hieronyma antioquensis, Ladenbergia macrocarpa, Palicourea andaluciana and Prestoea acuminata.
Species studied. In Colombia W. kalbreyeri is locally known as "gualte bola" or "macana". It is a monoecious palm with unisexual inflorescences, and up to 15 inflorescences per node. The infructescences contain from 7 to 20 pendulous branches with free fruits, which are globose to slightly ellipsoid, up to 3.5 cm long; seeds are ovoid to ellipsoid, 2 cm long each. This palm also has a solitary stem, reaching a maximum height of 20 m, supported by a cone of epigeic roots of about 1 m height; each root has stingers of 1 cm long. Its leaves are polystic, up to 5 leaves per palm, with an average length of 3.5 m (Henderson et al., 1995; Galeano and Bernal, 2010).
The natural distribution of W. kalbreyeri comprises part of the Andes of Colombia and Ecuador; particularly, in Colombia is more abundant on the western slopes of the Western Cordillera, and the northern part of the Central Cordillera; occasionally it may be found on the eastern slopes of the Cordillera Oriental. It is the most widely distributed species of its genus (Galeano and Bernal, 2010), and is common in primary forests (Galeano and Bernal, 1987; Henderson et al., 1995).
Survey units. In February 2009 we established and measured 10 permanent plots distributed systematically (at least separated by a minimum distance of 100 m). These plots were measured again one year later. We used different survey units to evaluate different sizes of palms as follows: (1) in plots of 20 x 50 m (1000 m2) were marked and measured all individuals of W. kalbreyeri with height greater than one meter. (2) Within each of these plots a subplot of 10 x 10 m (100 m2) was delimited to measure palms between 0.5 - 1.0 m heights. (3) Within each subplot we measured palms under 50 cm height in a quadrant of 2.5 x 2.5 m (6.25 m2). All individuals were marked with aluminum tags.
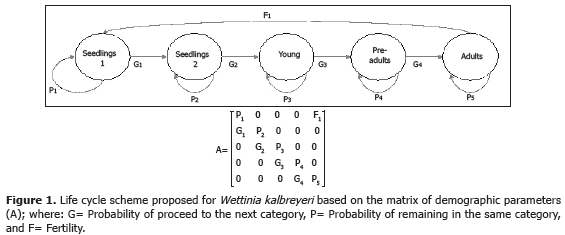
Population dynamics. To characterize the life cycle of Wettinia kalbreyeri we employed a matrix model structured by categories (Lefkovitch, 1965; Caswell, 2001) This model was based on morphological characteristics of individuals using five categories: (1) seedling 1, (2) seedling 2, (3) young, (4) pre-adult, and (5) adult (Table 1). The Lefkovitch transition matrix (denoted as A, Figure 1) was made with the proportion of palms remaining at the same category of development (Pij), the proportion of palms that advanced from one to the next category (Gij), and fertility (Fij) which was obtained as F = Nfp / g, where: Nfg is the number of fruits produced per palm, and g is the percentage of germination.
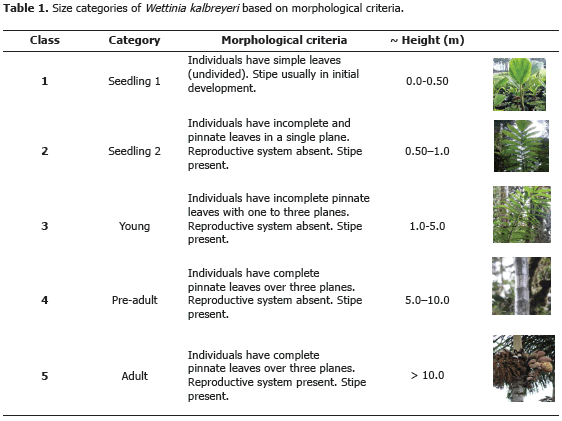
In order to evaluate the production of fruits (Nfg), 100 adult palms were selected. Observations of infructescence productivity were done monthly along a year. The number of fruits per inflorescence was estimated from the average number of fruits produced in a sample of 10 infructescences. Germination (g) was evaluated directly in the field as follows: we sowed 4 lots of 300 seeds in an area of 2 m2 each; seeds were sown in the forest floor no more than 5 cm depth; germination rate was measured one year later.
The projected population growth was obtained by multiplying the Leftkovitch transition matrix (A) by the column vector of density (Nt); the result describes the structure of the population in function of time. The basic matrix model is: Nt+1 = ANt. The finite rate of population growth (l) was calculated as the dominant eigenvalue of the transition matrix (A), and the intrinsic growth rate was calculated as: r = Ln l (Caswell 2001).
We also calculated the stable structure of population, which is a density vector that maintains constant the proportion along the growth projections of the population. That vector was denoted w, and corresponds to the dominant right vector on the matrix (A). Another demographic parameter evaluated was the reproductive value (v), which was calculated as: v = NtT x A and corresponds to the dominant left eigenvector of the matrix (A). Finally, we calculated the confidence interval for the finite rate of increase l, obtained from a Boopstrap resampling (Caswell, 2001) with 1,000 iterations.
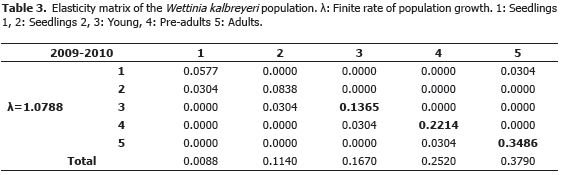
Prospective disturbance analysis. This analysis was based on the proposal by De Kroon et al. (1986) and De Kroon et al. (2000) to assess the impact on l of the relative changes in vital rates (growth, reproduction and survival); to do it, we calculated the elasticity matrix from the equation ei,j = [(pij/l)(viwj/{v,w})]. Subsequently, we used an important property of the elasticity matrix (Sijeij = 1), which means that each value of the matrix represents the proportional contribution of each element on population growth.
RESULTS
Population structure. The density of individuals was evaluated by categories of size (Figure 2). The highest density per ha occurred in seedlings 1 (X =129.520; DE=72.701; n=10), followed by seedlings 2, which decreased dramatically ( X=240; DE=187; n=10) and was similar to the density of young individuals (X=259; DE=214; n=10), and pre-adults (X=307; DE=192; n=10); while adults showed a significant rise (X=768; DE=263; n=10).
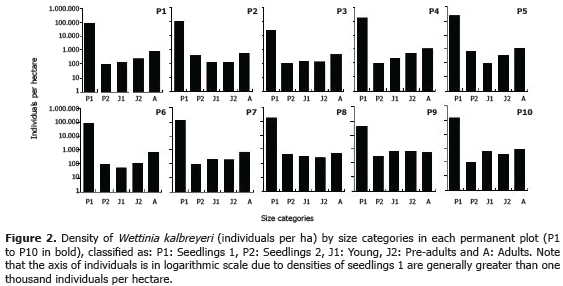
The average basal area of W. kalbreyeri was 21.34 m2 ha-1 (SD=8.84; n=10). In total were registered 78 tree species with diameter at breast height (DBH)>10 cm. In each plot (20 x 50 m) were found on average 23 ± 5 (n = 10) tree species (DBH>10 cm), its density was 1,752 ± 505 (n= 10) individuals ha-1, with a basal area of 16.40 m2 ha-1 (SD=6.41; n=10).
Population dynamics. The average transition matrix (Table 2) showed high values of permanence in all categories evaluated [0.707-0.992], with an increasing trend through categories: lower values occurred in the first three categories (seedlings 1, seedlings 2 and young) and maximum values were found in both pre-adults and adults. On the other hand, the probability of proceed to the next category in one year was limited [0.009-0.0833], with the lowest values for seedlings and the highest ones in pre-adults and adults. On 100 adult palms surveyed along one year, 62 infructesences were formed, with an average of 1,268.4 seeds each (SD=366.2; n=10). In total, we estimated a production of 603,770 seeds ha-1 year-1. The germination rate was about 15% (X=0.151; SD=0.042; n=1,200), and based on these figures, the estimated fertility was 118.71 seedlings per adult.
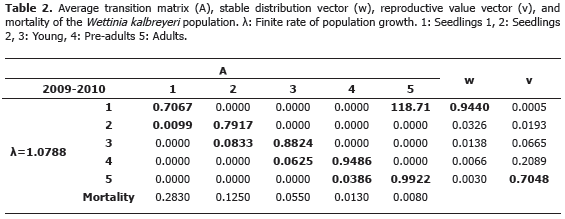
The mortality rate was higher at the initial steps of development and lower in adults (Table 2). The finite rate of increase was positive (95% confidence interval = 1.0321 - 1.1156), and the intrinsic rate of increase was 0.076; both numbers reflected a slight increase of the population density from 2009 to 2010. Seedlings 1 showed the largest number of individuals when the stable distribution of categories w (0.9440) is reached. Meanwhile, adults had the highest reproductive value v (0.7048; Table 2).
Prospective disturbance analysis. The average matrix of elasticity (Table 3) showed the relative contribution of three demographic processes: growth SGi,j = 0.122, fertility SFi,j = 0.003, and permanence SGi,j = 0.848. As size of individuals increases, their proportional contribution also increase (Table 3); these results show the importance of adults on the population dynamics and in management strategies. Likewise, permanence was the most important of the processes evaluated. The simulation of the effect of adult declination (which occurs frequently because of logging) showed that a negative population rate (l<1) occurs when the number of adult individuals is reduced in 288 ± 98 individuals (37.54%).
DISCUSSION
The number of individuals of W. kalbreyeri declined through categories, with a huge number of individuals in the first categories and a marked decrease in the following ones; this trend is similar to that reported for other tropical palms (Durán and Franco, 1992; Bernal, 1998; Bonesso et al., 2008; Thompson et al., 2009; Arango et al., 2010). The density of adults of W. quinaria was 1.160 per hectare (Waldrón, 2001), a number quite similar with our results. Though a special characteristic of Wettinia seems to be the high density of adults, more evaluations of density in other species of this genus are still necessary to uncover the potential factors involved in its high abundance in tropical montane forests.
The high probability of permanence and the low probability of growth found in W. kalbreyeri (Table 2) occur in populations with low rates of growth and mortality, which also correlates with long life cycles (Bernal, 1998; Arango et al., 2010). We found a marked reduction in mortality rates as size category increased. Despite the reduced number of studies on this subject, Bernal (1998) found that the high rate of mortality at the initial stages of plants growing in the understory is due to high intraspecific competition and the dependence on canopy gaps for their successful establishment. On the other hand, the mortality rate around 1% of W. kalbreyeri forests are similar to the rates reported for other tropical forests dominated by trees (Condit et al., 1995; Clark et al., 2004).
The low germination rate found in W. kalbreyeri (0.151) is common in palms; for instance, Bernal (1998) reported a value of 0.23 in Phytelephas semannii; nonetheless, even lower germination rates have been reported on Thrinax radita (0.08-0.17) (Olmsted and Álvarez, 1995). Several factors, either micro-climate or soil conditions, as well as seed predation (Fleury and Galetti, 2004; Salm, 2006; Grenha et al., 2010), may be involved in the current low rate of germination of W. kalbreyeri. Although we did not systematically evaluate seed predation, the attack of many seeds by a borer (Pachymerus sp, Bruchidae, Figure 3) was evident. This insect could be an important factor in the low rate of germination found in the present study because this genus has been reported as a seed predator in another palms species (Quiroga and Roldán, 2001; Ramos et al., 2001; Costa and Ramos, 2006); nonetheless, our report is atypical (Nuñez, personal communication) as much as 2,000 masl is the highest elevation previously recorded for this genus of insects, and the altitude of our study sites ranges between 2,200 and 2,800 m. This point is extremely important because it could be an effect of the current global warming (IPCC, 2007), which could bring an additional threat for the persistence of W. kalbreyeri; however, its evaluation is beyond the scope of this paper. Finally, we also observed a rodent (Heteromys sp, Heteromyidae), which has been reported as a predator of seeds in populations of W. kalbreyeri in the Central Cordillera of Colombia (Sánchez and Díaz, 2010), whose activity could also affect the germination of seeds.

The finite rate of population growth showed a slight trend of growth (l=1.079), which has been reported in several palms studied in similar periods of time of one year or even less than a year (Durán and Franco, 1992; Olmsted and Álvarez, 1995; Holm et al., 2008; Bernacci, 2001; Bernal, 1998; Zea, 1997; Suárez, 2001; Waldrón, 2001). However, studies involving two or more years of survey found variations in the rate of population growth (e.g. De Cássia et al., 2010; Souza and Martins, 2004; Arango et al., 2010). Fluctuations in the growth rate usually occur because density-dependence processes influence the growth and population dynamics. Therefore, the value of l found in our study should be interpreted as a preliminary estimation, which should be contrasted with rates obtained in future studies. Likewise, elasticity values reported here should be taken carefully, because this study was restricted to one year. In consequence, the population growth in the long term is still unknown (Benton and Grant, 1999; de Kroon et al., 2000). Despite these weaknesses, this is the first study on the demography of this important palm species and gives valuable information for its management and conservation.
We also found that the fragility of the population increases through size categories, being the adult’s size the most sensitive, which was also reported in other palm species (Souza and Martins, 2004; De Cássia et al., 2010, Arango et al., 2010, Rodríguez et al., 2005; Durán and Franco, 1992; Bonesso, 2006). This result is particularly critical in this population, given the current situation of intensive extraction of adult individuals of W. kalbreyeri.
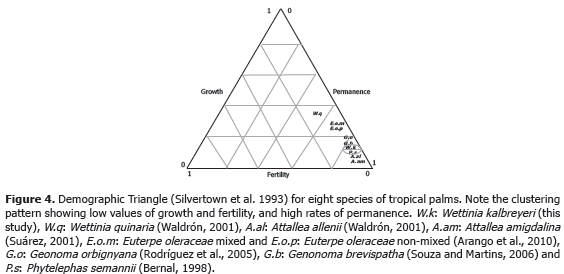
In summary, we conclude that W. kalbreyeri has a high productivity of fruits; although the germination rate was low, there were a high number of individuals in the categories of seedlings, which also have a higher mortality rate. Nevertheless, so far, the most important demographic process in this population was the permanence of adult individuals (Figure 4), which under natural conditions seems to guarantee the long-term persistence of this species. In addition, other palms species also have a similar pattern in their distribution of the demographic processes of fertility, permanence and growth (Figure 4).
Finally, the slow growth and high elasticity values found in the adult category of W. kalbreyeri, suggest that its removal could potentially threaten the persistence of the species in the long term. Consequently, the intensive and messy logging of adult individuals of W. kalbreyeri that has been ongoing for several decades has endangered the permanence of their populations in this region. For this reason, it is imperative to collect information during a longer time period in order to better characterize its life cycle and growth rates. With more information we could design management systems to achieve a balance between conservation and sustainable use (Schöngart, 2008).
ACKNOWLEDGMENTS
Thanks a lot to all who helped us in several steps of this study: L.A. Núñez, J.L. Toro, D. Suescún, J.C. Carvajal, J.E. Calle, A. Caro, D. Guerrero, S.M. Suárez, Z. Restrepo, N. Herrera, P.A. Gómez and G.A. Buitrago. We are especially grateful to J.G. Zamora, M.R. Rojas, and two anonymous reviewers for comments that improved the manuscript. This research was funded by the Research Institute of the School of Agrarian Sciences (IIFCA) and the National Research Direction (DINAIN, grant number-20101007362), both at Universidad Nacional de Colombia, and the Ecosystems Direction of Corporación Autónoma Regional del Centro de Antioquia (CORANTIOQUIA).
BIBLIOGRAPHY
Arango, D.A., A.J. Duque and E. Muñoz. 2010. Dinámica poblacional de la palma Euterpe oleracea (Arecaceae) en bosques inundables del Chocó, Pacífico colombiano. Revista de Biología Tropical 58(1): 465-481. [ Links ]
Benton, T.G. and A. Grant. 1999. Elasticity analysis as an important tool in evolutionary and population ecology. Trends in Ecology and Evolution 14(12): 467-471. [ Links ]
Bernacci, L.C. 2001. Aspectos da demografia da palmeira native Syagrus romanzoffiana (Cham.) Glassman, jerivá, como subsidios ao seu manejo. Tesis de doctorado. Instituto de Biología. Universidade Estadual de Campinas. Brazil. 138 p. [ Links ]
Bernal, R. 1998. Demography of the vegetable ivory palm Phytelephas seemannii in Colombia, and the impact of seed harvesting. Journal of Applied Ecology 35(1): 64-74. [ Links ]
Bonesso, M. 2006. Ecologia populacional da palmeira Geonoma schottiana Mart. em mata de galleria no Brasil Central. Tesis de Maestría. Instituto de Ciencias Biológicas. Universidade de Brasília. 83 p. [ Links ]
Bonesso, M., I, Belloni and I. Benedetti. 2008. Harvesting effects and population ecology of the buriti palm (Mauritia flexuosa L. f., Arecaceae) in the Jalapão Region, Central Brazil. Economic Botany 62(2): 171-181. [ Links ]
Caswell, H. 2001. Matrix population model: construction, analysis and interpretation. Second Edition. Sinauer Associates. Sunderland, Massachussets. 722 p. [ Links ]
Clark, D., C. Soto, L.D. Alfaro and J.M. Read. 2004. Quantifying mortality of tropical rain forest trees using high-spacial-resolution satellite data. Ecology Letters 7: 52-59. [ Links ]
Condit, R., S.P. Hubbell and R.B. Foster. 1995. Mortality rates of 205 neotropical tree and shrub species and the impact of a severe drought. Ecological Monographs 65(4): 419-439. [ Links ]
Corporación Autónoma Regional del Centro de Antioquia (Corantioquia). 2007. Plan de manejo Reserva Cuchilla Jardín - Támesis. Corporación Autónoma Regional del Centro de Antioquia - Corporación Académica Ambiental, Medellín. 377 p. [ Links ]
Henderson, A., G. Galeano and R. Bernal. 1995. Field guide to the palms of the Americas. Princeton University, New Jersey. 352 p. [ Links ]
Holdridge, L.R. 1978. Ecología basada en zonas de vida. Instituto Interamericano de Ciencias Agrícolas - IICA. San José. 216 p. [ Links ]
Holm, J A., C.J. Miller and W.P. Cropper. 2008. Population dynamics of the dioecious amazonian palm Mauritia flexuosa: Simulation analysis of sustainable harvesting. Biotropica 40(5): 550-558. [ Links ]
IPCC, 2007: Cambio climático 2007: Informe de síntesis. Contribución de los Grupos de trabajo I, II and III al cuarto informe de evaluación del Grupo Intergubernamental de Expertos sobre el Cambio Climático [Equipo de redacción principal: Pachauri, R.K. and A. Reisinger (Directores de la publicación)]. IPCC, Ginebra, Suiza. 104 p. [ Links ]
Lefkovitch, L.P. 1965. The study of population growth in organisms grouped by stages. Biometrics 21(1): 1-18. [ Links ]
Olmsted, I. and E.R. Álvarez. 1995. Sustainable harvesting of tropical trees: Demography and matrix models of two palm species in Mexico. Ecological Applications 5(2): 484-500. [ Links ]
Quiroga, V.D. and A.I. Roldán. 2001. The Fate of Attalea phalerata (Palmae) seeds dispersed to a tapir latrine. Biotropica 33(3): 472-477. [ Links ]
Ramos, F.A., I. Martins, J.M. Farias, I.C. Silva, D.C. Costa and A.P Miranda. 2001. Oviposition and predation by Speciomerus revoili (Coleoptera, Bruchidae) on seeds of Acrocomia aculeata (Arecaceae) in Brasilia, DF, Brazil. Brazilian Journal of Biology 61(3): 449-454. [ Links ]
Rodríguez, S., M.A Orjuela and G. Galeano. 2005. Demography and life history of Geonoma orbignyana: An understory palm used as foliage in Colombia. Forest Ecology and Management 211: 329-340. [ Links ]
Salm, R. 2006. Invertebrate and vertebrate seed predation in the Amazonian palm Attalea maripa. Biotropica 38(4): 558-560. [ Links ]
Sánchez, C. and J.F. Díaz. 2010. Hábitos alimenticios de Heteromys australis (Rodentia: Heteromyidae) en el norte de la Cordillera Central de Colombia. Mastozoología Neotropical 17(1): 189-194. [ Links ]
Schöngart, J. 2008. "Growth-Oriented Logging (GOL): A new concept towards sustainable forest management in Central Amazonian várzea floodplains". Forest Ecology and Management. 256(1-2): 46-58. [ Links ]
Silvertown, J., M. Franco and E. Menges. 1996. Interpretation of elasticity matrices as an aid to the management of plant populations for conservation. Conservation Biology 10(2): 591-597. [ Links ]
Suárez, Z.J. 2001. Evaluación del estado de conservación de la palma de almedrón o taparo (Attalea amigdalina Kunth, Palmae). Trabajo de grado de Biología. Facultad de Ciencias. Pontificia Universidad Javeriana. Bogotá. 116 p. [ Links ]
Svenning, J.C. and H. Balslev. 1997. Small-scale demographic disequilibrium of Iriartea deltoidea (Arecaceae) in Amazonian Ecuador. pp: 263-274. En: Valencia, R. and H. Balslev (eds.). Estudios sobre diversidad y ecología de plantas. Pontificia Universidad Católica del Ecuador, Quito. 289 p. [ Links ]
Thompson, L.N., M. Moraes and M. Baudoin. 2009. Estructura poblacional de la palmera endémica Parajubaea torallyi (Mart.) Burret en zonas aprovechadas del área natural de manejo integrado El Palmar (Chuquisaca, Bolivia). Ecología en Bolivia 44(1): 17-35. [ Links ]
Waldrón, T. 2001. Estudio de la dinámica poblacional de las palmas de Táparo (Attalea allenii) and Memé (W. quinaria), como herramienta para su uso sostenible en la Costa Pacífica del Chocó. Trabajo de grado Biología. Universidad Nacional de Colombia. Facultad de Ciencias, Bogotá. 81 p. [ Links ]
Zea, C. E. 1997. Demografía de Mauritia flexosa en una sábana mal drenada de la Orinoquia Colombiana and sus aplicaciones en la evaluación de alternativas de manejo. Trabajo de grado Biología. Pontificia Universidad Javeriana. Facultad de Ciencias. Bogotá 185 p. [ Links ]
