










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.66 no.1 Medellín Jan./June 2013
Evaluation of the Heritability of Resistance to Phytophthora infestans (Mont) de Bary in a Population of Solanum phureja Juz et Buk
Evaluación de la Heredabilidad de la Resistencia a Phytophthora infestans (Mont) de Bary en una Población de Solanum phureja Juz et Buk
Luz Fanny Orozco Orozco1; Liliana Andrea Ramírez Franco2 and José Miguel Cotes Torres3
1 Agronomic Engineering. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias Agrarias - Departamento de Ciencias Agronómicas. A.A. 1779, Medellín, Colombia. <lforozco@unal.edu.co>
2 Agronomic Engineering. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias Agrarias - Departamento de Ciencias Agronómicas. A.A. 1779, Medellín, Colombia. <liaramirezfr@unal.edu.co>
3 Associate Professor. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias Agrarias - Departamento de Ciencias Agronómicas. A.A. 1779, Medellín, Colombia. <jmcotes@unal.edu.co>
Received: May 23, 2012; acepted: November 21, 2012.
Abstract
Potato late blight is a common name for the most important and destructive disease caused by Phytophthora infestans (Mont. de Bary) in potato crops. Resistance genes for the disease have been found in Solanum phureja, and are of great importance in implementing important tools such as plant breeding in disease management The aim of this study was to estimate the heritability of resistance to P. infestans in a population of S. phureja. This research was carried out in potato cultivations in two locations in the municipality of La Unión (Antioquia, Colombia). Pathogen severity was evaluated in leaves using a diagrammatic scale. Heritability was measured between families and within families by stratified mass and simple mass selection. There were no statistically significant differences among the heritabilities from the methods of selection; however, significant differences were found between locations. For the first location, heritability estimations were in the range of 0.35 and 0.54, while values obtained in the second location were between 0.05 and 0.08.
Key words: Potato late blight, yellow creole potato, half-sib families, potato breeding.
Resumen
El tizón tardío o gota de la papa producido por Phytophthora infestans (Mont. de Bary), es la enfermedad más limitante del cultivo. El mejoramiento genético, con la obtención de variedades resistentes es un componente importante del manejo integrado del patógeno; en la especie Solanum phureja se han encontrado genes de resistencia a la enfermedad. La heredabilidad permite estimar la proporción de la variabilidad total debida al efecto de los genes; y esta depende de la población en estudio y de la unidad de selección que se utilice. El objetivo de este estudio fue estimar la heredabilidad de la resistencia a la gota de la papa en una población de S. phureja. Esta investigación, se desarrolló en dos localidades del municipio de La Unión (Antioquia, Colombia), en la cual se evaluó la severidad de la enfermedad utilizando una escala diagramática de severidad para hoja. La heredabilidad se evaluó bajo los métodos de selección entre familias, entre y dentro de familia, masal estratificada y masal simple; sin encontrar diferencias estadísticamente significativas entre la heredabilidad obtenida para los diferentes métodos de selección, pero sí entre las localidades; así, en la vereda La Cabaña, la estimación de la heredabilidad varió entre 0,35 a 0,54, y en la vereda San Juan la estimación de la heredabilidad varió entre 0,05 a 0,08.
Palabras clave: Gota o tizón tardío de la papa, papa criolla, familia de medios hermanos, mejoramiento genético de papa.
The potato is the fourth largest food crop around the world after wheat, maize and rice. World production of fresh potatoes in 2006 was about 314 Megatons, almost half (159 Megatons) was produced by developing countries in Asia, Africa and Latin America (FAO, 2008); in these countries, potatoes are consumed as a carbohydrate source; furthermore, it contains vitamins C, B6, and B1 and minerals such as phosphorus, potassium, calcium, and magnesium; and micronutrients such as iron and zinc which are assimilated through its consumption (Bradshaw and Ramsay, 2009).
The potato is an agricultural product which is widely consumed by the Colombian population. The annual diet of an average Colombian is 71 kg. On the other hand, the available total production is used for either fresh consumption (80-92%) or processed (7-8%) (Díaz et al., 2003).
In Colombia, Solanum phureja is known as the yellow creole potato. It is produced in the departments of Nariño, Cundinamarca, Boyacá, Antioquia, Santander and Norte de Santander. The crop is cultivated on small farms, with low levels of technology (Jaramillo and Botero, 2007). In 2008, in Colombia, there were 138,315 ha with potato; 6% of which were with S. phureja (Rodríguez et al., 2009).
Potato production is affected by a diversity of pests and diseases. Gabriel et al. (2007) reported 25 viruses, 38 fungi, 6 bacteria, 2 phytoplasma and 1 viroid. The potato crop is also affected by 68 species of nematodes and 128 insect pests. Furthermore, abiotic factors can also affect yields, such as frosts and droughts.
Phytophthora infestans causes the most limiting disease of the potato; this pathogen belongs to the Phytophthora genus which contains 60 species which are cataloged as important agents of plant diseases (Laxalt et al., 2002; van't Klooster et al., 2000). P. infestans is the biological agent of the late blight or potato blight; it affects several crops from the Solanaceae family. In the potato, this disease dries up leaf tissues and also affects tuber tissues (Xiben et al., 2005; Skottrup et al., 2006).
In Colombia, late blight is considered one of the most devastating plant diseases; with economic importance not only in the potato (S. tuberosum, S. phureja L.) but also in the tomato (Solanum lycopersicum M.), this pathogen also represents a serious threat to other crops that are being grown on a large scale, such as the cape gooseberry (Physalis peruviana), tree tomato (Solanum betaceum Cav.) and lulo (Solanum quitoense Lamarck) (Vargas et al., 2009).
P. infestans dispersion is carried out by wind, the sporangia are able to detach, travel in the air and germinate directly; and penetrate plant leaves under moist conditions (Skottrup et al., 2006). This pathogen can infect both leaves and tubers and it also can spread rapidly through host tissues causing brownish-black lesions or necrosis. Under favorable conditions, this pathogen can reduce the plant yield due to damage to the foliage and decrease in tuber development. Likewise, it causes tubers putrefaction before and after the storage process. For these reasons, it is considered the most powerful disease that diminishes crop yields (Runno - Paurson et al., 2010).
Some of the mechanisms that confer a greater pathogenicity to this organism have been identified; some of which are the migration on a global scale of two mating types of P. infestans and the constant sowing of the same potato varieties, containing the major genes. These genes can contribute to selection of pathogenic strains of the fungus which are able to show major aggressiveness against susceptible crops (López et al., 1997). From the population point of view, sexual recombination and selection of resistant individuals are considered the evolutionary mechanisms that have allowed the reproductive success of P. infestans. This last reason has been the main cause of variants of the pathogen (Reis et al., 2006).
Disease management generally consists of fungicide applications, according to the International Centre of Potato (2010), ten or more foliage applications are needed per crop cycle to achieve profitability, representing between 10 and 15% of expenditures in the production. It has been estimated that world costs to control P. infestans are close to a billion dollars annually (Fiorini et al., 2010). Inappropriate use of this control strategy could contribute to the appearance of new resistant fungus strains (Gabriel et al., 2007); and moreover, to environmental contamination of soils and water (Estrada et al., 2002).
Successful management of P. infestans entails the consideration of several control measures, such as using resistant and high yielding cultivars (Gabriel et al., 2007). Fiorini et al. (2010) determined that using resistant cultivars to control late blight is the most effective strategy because it prevents or makes the establishment of the pathogen difficult in the crop and reduces the contamination of soils, water and the food due to fungicide use.
At the beginning of the twentieth century, most breeding programs were focused on finding cultivars with increased resistance to this pathogen; these programs were not successful because their work just considered one single gene resistance obtained from a pair of genes of the Mexican species Solanum demissum. Those results could be related to the pathogen capacity to develop new biotypes (Nústez et al., 2000).
There is a type of resistance that confers stability to the potato throughout the time and space to the damage caused by late blight, it is called durable resistance to P. infestans or polygenic resistance; responses to damage are related to an apparent control made by four or more pairs of genes found in S. tuberosum, S. andigena and S. phureja. Besides wild species with durable resistance to P. infestans, there are: S. avilesíi, S. bulbocastanum, S. brachvcarpum, S. conrmersonii, S. iopetalun, S. stoloniferurn, S. palustre, (brevidens) and S. acaule (Nústez et al., 2000).
In S. bulbocastanum, durable resistances have been found to all known strains of P. infestans. A cultivar's progeny has resistance even after several crop cycles. In this cultivar, a group of four homologous genes has been found in chromosome number 8 which is responsible for this type of resistance (Irzhansky and Cohen, 2006).
Estrada (2000) identified S. phureja as a valuable cultivar in breeding programs, this species presents disease resistance genes and it is also compatible with wild cultivars. Field and lab experiments made by Gabriel et al. (2007) in Bolivia, determined that S. Phureja is one of the cultivars with a good resistance response to P. infestans after three cultivation cycles.
Escallón et al. (2005) carried out evaluations and determined the absence of major genes involved in P. infestans resistance in some genotypes of S. phureja. Evers et al. (2006) established that there is a family of genes in S. phureja called PR-1, which codifies pathogen toxic proteins and therefore is involved in the horizontal resistance of the species.
Heritability measures the fraction of phenotype variability of a characteristic that can be attributed to genetic differences (Pierce, 2010). The characteristics of P. infestans resistance in a S. tuberosum population were evaluated by Zuñiga (2000) under field conditions in Toluca's valley (Mexico) and relatively low values of heritability were obtained (0.27). Haynes et al. (2007) developed a population of tetraploid potatoes adapted to short days and with horizontal resistance to late blight, this population was evaluated during four crop cycles and its heritability of resistance to P. infestans was estimated at 0.78, researchers considered this population as an important source of material for breeding programs in order to get resistant cultivars. Evaluations of heritability of late blight resistance in populations of mating S. phureja x S. stenotomum were carried out by Haynes and Christ (1999) obtaining a value of 0.78, qualified as high heritability.
The aim of this study was to estimate and characterize the heritability of resistance to P. infestans in a population of half-sib families under field conditions in the municipality of La Unión (Antioquia, Colombia).
Materials and methods
Location. Pre-basic potato seeds were obtained from the Experimental Station Paysandú of the Universidad Nacional de Colombia, Sede Medellín located in the Santa Elena region (Medellín - Antioquia) with an altitude of 2,550 masl, with an annual average temperature of 14 °C and cumulated rainfall of 2,000 mm per year. The genetic assessment of the resistance was done in field conditions in two different places in the La Unión (Antioquia, Colombia) municipality. The experiments were established in the La Cabaña and San Juan region with altitudes of 2,512 and 2,530 masl, respectively, an annual average temperature of 14 °C and an annual average relative humidity of 87%.
Plant material. 75 unrelated S. phureja genotypes were taken in the first stage of the research; they produced good sexual seeds from half-sib families under the agroecological conditions in Santa Elena. In the second stage of the research, 49 half-sib families were selected by agroecological adaptation and higher rates of reproduction. These families were assessed on potato farms.
Sexual seed obtainment. True potato seeds (TPS) were obtained from potato berries. These fruits were produced by natural pollination and the procedure was carried out according to the methodology for extraction, washing and storage proposed by the International Centre of Potato (2010). A mixture of soil and peat (3:1) was applied to prepare the seedbeds; then the mixture was placed on properly labeled Styrofoam plates. Under mesh house conditions, transplanting was done two and a half months after seedbed preparation. The plants were placed in pots or plastic bags with 1 kg of soil and 25 g of 10-2020 grade granular fertilizer. The harvest and genotype selection for planting in field conditions were realized three months after transplanting. Families with the largest number of tubers were selected.
Establishment and management of field experiments. The minitubers were planted in field crop conditions; the distance between individuals was 0.3 m and 0.9 m between rows. Two replicates of each family were placed in the ground with a border crop of susceptible S. phureja (cv. Criolla Colombia) that was planted in order to ensure the inoculums. Agronomical practices for controlling weeds and pests were employed during the field experiment. Four weeks after planting, P. infestans control was carried out when most of plants had developed 30% of the leaf area. Afterwards, neither chemical nor biological controls were applied to control the disease.
Assessment of P. infestans resistance. The disease scores were taken every week in the S. phureja population located in the La Unión municipality by using the 1-9 grade scale proposed by the International Centre of Potato (Henfling, 1987) (Table 1). These readings were taken until most of the genotypes evaluated reached a value of 7 and 9.
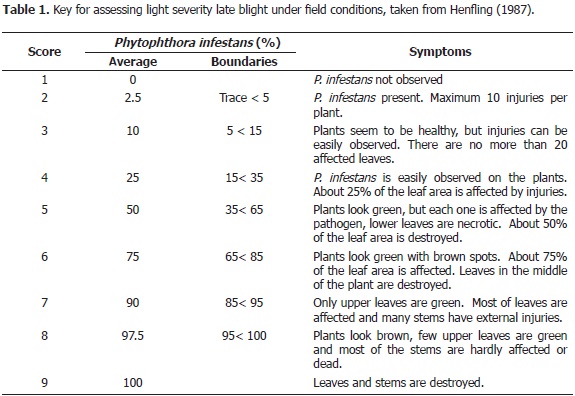
The average disease severity values were analyzed and the Area Under the Disease Progress Curve (AUDPC), was calculated as it describes IPC (2010). Relative AUDPC was obtained using the quotient between observed AUDPC and potential AUDPC, adopting a completely susceptible genotype from the beginning of the assessment. Selection of late blight resistance was determined in an inverse sense, meaning, a lower relative AUDPC corresponded to a higher level of resistance of the genotype to late blight.
Experimental design and statistical analysis. Each trial was laid out as a 7x7 lattice, unbalanced design with two replicates, each experimental unit was formed by five half-sib families and the border crop was a cultivar susceptible to P. infestans (S. phureja cv. Criolla Colombia).
The univariate linear additive genetic model (Sorensen and Gianola, 2002) was used for obtaining variance components to calculate heritability of P. infestans resistance in the farms, the following model was used:
y=1µ + z1f + z2h + z3b + e
where y is a vector of size n of the disease incidence variable, µ is the general mean of disease incidence, f is a vector of size containing genetic effects of families, h is a vector of size containing genetic effects of half-sibs, b is a vector of size containing systematic environmental effects associated with incomplete blocks and Z is the known incidence matrices and localized values of the effects corresponding to each observed data.
In this model, it is assumed that the vectors f, h, b, and e have normal distribution with a mean value of zero and variances Iσ2f,Iσ2h,Iσ2b and Iσ2e. The Bayesian estimation technique was applied for parameter estimation, and the median of a posteriori distribution was obtained as a pointed estimation because this estimate minimizes the risk of Bayes under absolute loss function. For all parameters, non-informative prior distribution was used (Sorensen and Gianola, 2002). The data were analyzed by using the free program language R (2011) and the R package Markov Chain Monte Carlo MCMCglmm (2010) which runs the Gibbs sampling algorithm, which is very usual in these models. In order to explore the complex posterior distribution of interested parameters, 1,030,000 MCMCs were obtained, the first 10,000 iterations were considered as the burn in period and one sample of each 10 was taken to obtain the marginal distribution of each parameter.
The heritability estimates were obtained from the corresponding MCMC values of the variance parameter. Because heritability depends on the population and the selection unit used and its calculation is the quotient between additive variance in the selected units and phenotypic variance in the selected units (Cruz and Carneiro, 2003), the heritability was evaluated by using four plant selection methods: 1) between families, 2) between and within families, 3) simple mass selection and 4) stratified mass selection (Table 2).
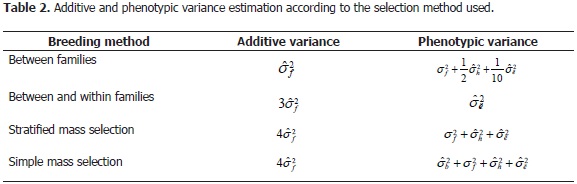
For each breeding method, the expected genetic gain was calculated by using the following equation:
Expected Genetic Gain = i x h2 x  p
p
where i represents the intensity of selection and depends on the population or genotype of a group
selected and also depends on phenotypic values corresponding to a normal distribution; h2 is called the narrow-sense heritability and it is a useful statistic to describe relative amounts of additive genetics, and p is the phenotypic standard deviation for the selection units (Falconer and Mackay, 1996).
Results and discussion
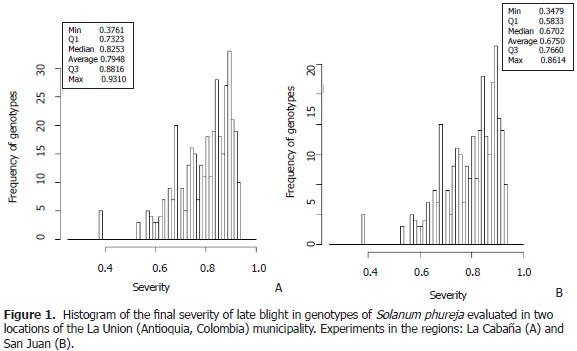
The experimental results allowed for establishing that the trials with late blight presented high levels of pathogen inoculums, and that the last evaluation of disease severity fluctuated between 38 and 93% in La Cabaña; and 35 and 86% in San Juan (Figure 1). It was seen that individuals from the population were submitted to high inoculum pressures.
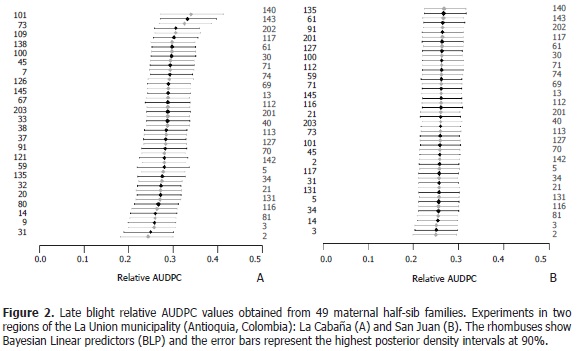
Based on the estimate of relative AUDPC, the five maternal families with lowest relative AUDPC were the families 2, 31, 3, 9, and 8 for the La Cabaña experiment; and the families 142, 3, 70, 14, and 33 were found in the San Juan experiment. It is important to highlight that family number 3 presented low values of relative AUDPC in both locations. Note that in the population located in La Cabaña, the relative AUDPC had a greater variation range (between 0.24 and 0.34) when compared with the population located in San Juan (Figure 2) where values were more uniform (between 0.25 and 0.27).
These results allow for establishing that the evaluated population of S. phureja contains P. infestans resistance genes, because the field conditions guarantied high expression of the symptoms. According to Castillo et al. (2000), S. phureja is one of the potato species with a high resistance level to late blight. Furthermore, Gabriel et al. (2007) carried out evaluations during three crop cycles to observe the response of different materials exposed to P. infestans, finding that genotypes of S. phureja finally presented a high resistance level to late blight and the relative AUDPC was 541.
There were no statistically significant differences in the narrow sense heritability evaluated under the four selection methods. However, there were significant differences between the locations evaluated. Interval estimates of heritability in La Cabaña were between 0.35 and 0.54 while values in San Juan were between 0.05 and 0.08 (Table 3). The low heritability presented in the experiment located in San Juan could be associated with a narrow range of variation observed in the predicted values of the families. Thus, in this experiment, the environmental effect was determinant for the phenotypic response.
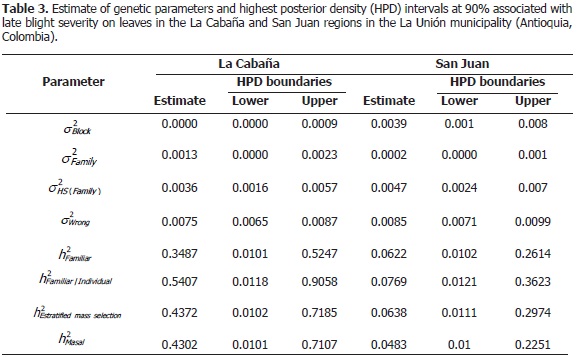
Poehlman (2003) mentioned that genetic traits such as yield and disease resistance are highly influenced by environmental conditions and, generally, scored heritability values are low; however, Nústez (2011) explained that increasing heritability estimations could be related to two important aspects: presence of a greater genetic variation in the population and environmental conditions where individuals are commonly developed.
Medium and high heritability values of late blight resistance were obtained in La Cabaña. Constanzo et al. (2004) found high narrow sense heritability (0.67) for the same trait; they evaluated a population of 230 full-sib families as a product of mating S. phureja x S. stenotomum, during three crop cycles in Pennsylvania (U.S). Haynes and Christ (2006a) evaluated a crossing population during two crop cycles; they also observed resistance levels to late blight and the estimated heritability ranged between 0.77 and 0.78. Their conclusion focused on the important genetic component in this hybrid. Landeo et al. (1999) found intermediate and high heritability values for late blight resistance. In this research, narrow sense heritability for late blight horizontal resistance was estimated in a population of B3C1 Potato and the values obtained were 0.40, 0.43 and 0.53, ensuring successful selection.
Haynes and Christ (2006b) started with incorporation of horizontal type resistance genes in a population of tetraploid potatoes adapted to short days in Peru; when researchers estimated narrow sense heritability, they found a range between 0.41 and 0.71, which induced the conclusion that P. infestans resistance improvement is possible.
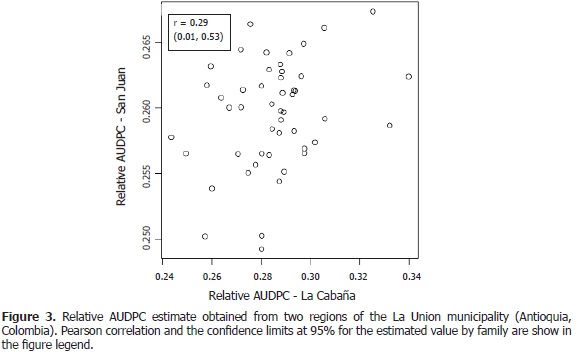
The lack of correlation between environments is due to the presence of the genotype X environment interaction and this is one way to measure its magnitude (Falconer and Mackay, 1996). Genetic correlation of the trait severity to late blight between the locations La Cabaña and San Juan is low (Figure 3), which evidences a high genotype X environment interaction in resistance expression, this means that perhaps the gene expression changes according to the environments or different genes are expressed in different environments. Furthermore, according to Sousa et al. (2003) when a genetic correlation coefficient has low values between locations, this could indicate that different genes in each environment are involved in the phenotypic response or the expression of the same genes are affected by the environment.
There were no statistically significant differences between the expected genetic progresses in the four plant selection methods evaluated (Figure 4). However, it must be noted that for both methods, selection between families and between and within families, two crop cycles are required for the construction of a new selection cycle. Thus, these methods have a disadvantage compared with other evaluated selection methods. Therefore, simple mass selection and stratified mass selection are better alternatives to promote genetic improvement of late blight resistance characteristics based on plant selection methods.
In La Cabaña, high values of heritability were obtained (Table 3); and therefore the highest expected genetic progress of the selection. So, this location could be recommended for progeny tests in breeding programs in order to obtain a new cultivar with field late blight resistance. In this location, if fixed the truncated selection point at 20% of genotypes with a lower relative AUDPC, then we expected a progress of the resistance close to 15 points in percentage in the level of AUDPC resistance for late blight resistance. The same procedure in San Juan only presented a progress of three points in percentage in the level of late blight resistance. This result could be due to the lack of additive, dominant and epispastic gene actions and the presence of an important environmental effect (Soomro et al., 2010).
Conclusion
Heritability of late blight resistance in the population of S. phureja presented contrasting results between the evaluated locations. The trial developed in the La Cabaña region gave medium values of the trait (between 0.35 and 0.54) while very low values (0.05 and 0.08) were obtained in the trial developed in San Juan. Medium to low values of heritability indicate that the additive effect of genes does not have an influence on the proportion of observed phenotypic variability, while environmental conditions have a very strong effect. In addition, there was no genetic correlation for late blight severity between the locations; this might be evidence that the same genetic mechanisms are not involved in the expression of pathogen severity in both locations.
Acknowledgements
This research was carried out under the projects: 20101008596 Evaluación de heredabilidad de la resistencia de Solanum phureja Juz et Buck a Spongospora subterranea f. sp subterranea-Jóvenes investigadores 2009, supported by COLCIENCIAS and the Universidad Nacional de Colombia - Medellín; and 20101008002 Evaluación fenotípica y genotípica de la colección colombiana de Solanum phureja por resistencia a Spongospora subterranea - Fase 2, supported by the Facultad de Ciencias Agrarias of the Universidad Nacional de Colombia - Medellín. The authors express their gratitude to all members of the research groups Mejoramiento y Producción de Especies Andinas y Tropicales (COL0039484) and Grupo de Investigación en Papa (COL0010065), who participated in the develop of this research. Finally, the authors give special thanks to Oscar Jair Rodríguez Rodríguez for his important help in reviewing the final version of the manuscript.
Bibliography
Bradshaw, J.E. and G. Ramsay. 2009. Potato origin and production. In: Advances in Potato Chemistry and Technology. Singh, J. and L. Kaur. Elsevier Inc. In: http://www.rlc.fao.org/es/agricultura/produ/papa.htm. accessed: March 2011. [ Links ]
Castillo, J., A. Estévez, M.E. González, O. Moré, E. Ortiz y U. Ortiz. 2000. Selección de progenitores para el mejoramiento de papa (Solanum tuberosum L.). Cultivos Tropicales 21(2): 49-59. [ Links ]
Costanzo, S., B.J. Christ, and K.G. Haynes. 2004. Late blight resistance in a diploid full-sib potato family. Plant Breeding 123(4): 377-381. [ Links ]
Cruz, C.D. and P.C. Carneiro. 2003. Modelos biométricos aplicados ao melhoramento genético Volumen 2. Universidade Federal de Viçosa (UFV), Viçosa. 585 p. [ Links ]
Díaz, M.M., D.A. Fajardo, J.D. Moreno, C. García y V.M. Núñez. 2003. Identificación de genes R1 y R2 que confieren resistencia a Phytophthora infestans en genotipos colombianos de papa. Revista Colombiana de Biotecnología 5(2): 40-50. [ Links ]
Escallón, R., M. Ramírez y E. Ñústez, 2005. Evaluación del potencial de rendimiento y de la resistencia a Phytophthora infestans (Mont. de Bary) en la colección de papas redondas amarillas de la especie Solanum phureja (Juz. et Buk.). Agronomía Colombiana 23(1): 35-41. [ Links ]
Estrada, N. 2000. La biodiversidad en elmejoramiento genético de papa. Centro de Información para el Desarrollo CIP, La Paz, Bolivia. 372 p. [ Links ]
Estrada, N., C. Ñústez y J. Gabriel. 2002. La ampliación de la base genética de la resistencia a Phytophthora infestans en los programas de mejoramiento genético de la papa en la zona andina de Suramérica. pp. 81-87. En: Libro de Actas del Congreso Iberoamericano de Investigación y Desarrollo en Patata. Vitoria-Gasteiz, España. [ Links ]
Evers, A., C. Schweitzer, N. Nicot, S. Gigliotti, M.R. Herrera, J.F. Hausman, L. Hoffmann, B. Trognitz, J. Dommes and M. Ghislain. 2006. Two PR-1 loci detected in the native cultivated potato Solanum phureja appear differentially expressed upon challenge by late blight. Physiological and Molecular Plant Pathology 67(3-5): 155-163. [ Links ]
Falconer, D.S. and T. Mackay. 1996. Introduction to quantitative genetics. Fourth edition. Addison Wesley Longman, England. 464 p. [ Links ]
FAO. 2008. La papa: una aliada clave de la seguridad alimentaria. En: http://www.rlc.fao.org/es/agricultura/produ/papa.htm; consulta: agosto 2011. [ Links ]
Fiorini, C., D. Silva, E. Mizubuti, J. Barros, L. Silva, C. Milagres e M. Zaparoli. 2010. Caracterização de linhagens de tomateiro originadas de cruzamento interespecífico quanto à resistência à requeima. Horticultura Brasileira 28(2): 197-202. [ Links ]
Gabriel, J., F. Forqueda, G. Plata y E. Fernández. 2007. Caracterización de genotipos de papa de Europa y Latinoamérica por resistencia a tizón y propiedades culinarias. Revista Latinoamericana de la Papa 14(1): 10-23. [ Links ]
Haynes, K.G and B.J. Christ. 1999. Heritability of resistance to foliar late blight in a diploid hybrid potato population of Solanum phureja x Solanum stenotomum. Plant Breeding 118(5): 431-434. [ Links ]
Haynes, K.G. and B.J. Christ. 2006a. Improvements in foliar late blight resistance in a diploid hybrid Solanum phureja S. stenotomum population. [abstract]. American Journal of Potato Research 83(2): 112-113. [ Links ]
Haynes, K.G. and B.J. Christ. 2006b. Recurrent maternal half-sib selection improves resistance to foliar late blight in a diploid hybrid Solanum phureja-Solanum stenotomum population. American Journal of Potato Research 83(2): 181-188. [ Links ]
Haynes, K., R. Goth, D. Lambert and B. Christ. 2007. Evaluation of a short-day adapted tetraploid potato population with horizontal resistance to Phytophthora infestans under long-day conditions in northern Maine. American Journal of Potato Research 84: 459-466. [ Links ]
Henfling, J.W., 1987. El tizón tardío de la papa: Phytophthora infestans. Boletín de Información Técnica Centro Internacional de la Papa CIP 4: 25. [ Links ]
International Centre of Potato. 2010. Procedimientos para pruebas de evaluación estándar de clones avanzados de papa. Guía para colaboradores internacionales. Centro Internacional de la Papa (CIP), Lima, Perú. 151 p. [ Links ]
Irzhansky, I. and Y. Cohen. 2006. Inheritance of resistance against Phytophthora infestans in Lycopersicon pimpenellifolium L3707. Euphytica 149(3): 309-316. [ Links ]
Jaramillo, S. y J.M. Botero. 2007. Respuesta de diferentes poblaciones de Spongospora subterranea f. sp subterranea a la rotación con dos variedades de papa (Solanum tuberosum ssp. andigena). Revista Facultad Nacional de Agronomía Medellín 60(2): 3859-3876. [ Links ]
Landeo, J.A., M. Gastelo, G. Beltrán, and L. Díaz.. 1999. Quantifying genetic variance for horizontal resistance to late blight in potato breeding population B3C1. pp. 63 - 68. CIP Program Report 1999 - 2000. Research on Potato, Lima, Perú [ Links ].
Laxalt, A.M., M. Latijnhouwers, M. Hulten and F. Govers. 2002. Differential expression of G protein α and β subunit genes during development of Phytophthora infestans. Fungal Genetics and Biology 36(2): 137-146. [ Links ]
López, J.B., M.E. Márquez, S. Jaramillo, J.L. Zapata, J.J. Mazo y L.F. Patiño. 1997. Determinación de razas fisiológicas y tipo de apareamiento en aislamientos de Phytophthora infestans (Mont) de Bary. Revista Latinoamericana de la Papa 9(10): 156-170. [ Links ]
Nústez, C.E. 2011. Estudios fenotípicos y genéticos asociados a la calidad de fritura en Solanum phureja Juz et Buk. Tesis de grado. Universidad Nacional de Colombia, Sede Bogotá [ Links ].
Ñústez, C.E., N. Estrada, C. García, S. Tinjacá, J.P. Medina y L. Schilde. 2000. Evaluación de la resistencia a P. infestans, cruzabilidad y potencial de rendimiento de 15 clones de la hibridación somática Solanum bulbocastanum + Solanum phureja en Colombia. pp. 427-434. En: Libro de Actas del Congreso Iberoamericano de Investigación y Desarrollo en Patata, Vitoria-Gasteiz, España. [ Links ]
Pierce, B. 2010. Genética un enfoque conceptual. Tercera edición. Editorial Médica Panamericana, Madrid. España. 940 p. [ Links ]
Poehlman, J.M. 2003. Mejoramiento genético de las cosechas. Segunda edición. Editorial Limusa, México. 506 p. [ Links ]
Reis, A., F. Ribeiro, e E. Mizubuti. 2006. Caracterização de isolados de Phytophthora infestans do Distrito Federal e de Goiás. Fitopatologia Brasileira 31(3): 270-276. [ Links ]
Rodríguez, L.E., C.E. Ñústez y N. Estrada. 2009. Criolla Latina, Criolla Paisa y Criolla Colombia, nuevos cultivares de papa criolla para el departamento de Antioquia (Colombia). Revista Agronomía Colombiana. 27(3): 289-303. [ Links ]
Runno-Paurson, E., T. Remmel, A. Ojarand, A. Aav and M. Mänd. 2010. The structure of Phytophthora infestans populations from organic and conventional crops. Eurpean Journal Plant Pathology 128(3): 373-383. [ Links ]
Skottrup, P., M. Nicolaisen and A. Justesen. 2006. Rapid determination of Phytophthora infestans sporangia using a surface plasmon resonance immunosensor. Journal of Microbiological Methods 68(3): 507-515. [ Links ]
Sorensen, D. and D. Gianola. 2002. Likelihood, Bayesian, and MCMC methods in quantitative genetics. First edition. Springer-Verlag, New York. 740 p. [ Links ]
Soomro, Z.A., A.S. Kumbhar, A.S. Larik, M. Imram and S.A. Brohi. 2010. Heritability and selection response in segregating generations of upland cotton. Pakistan Journal of Agricultural Research 23(1-2): 25-30. [ Links ]
Sousa, VO., R. Rea y R. Briceño. 2003. Uso de la repetibilidad clonal en la selección de ambientes en ensayos regionales de variedades de caña de azúcar en los estados de Lara y Yaracuy. Bioagro 15(02): 77-82. [ Links ]
The R Foundation for Statistical Computing, 2009. http://www.r-project.org; accessed: March 2010. [ Links ]
Van't Klooster, J.W., G. van den Berg-Velthuis P. van West and F. Govers. 2000. Tef1, a Phytophthora infestans gene encoding translation elongation factor 1alpha. Gene 249(1-2): 145-151. [ Links ]
Vargas, A., L. Quesada, .M. Céspedes, N. Carreño, A. González, A. Rojas, P. Zuluaga, K. Myers, W. Fry, P. Jiménez, A. Bernal and S. Restrepo. 2009. Characterization of Phytophthora infestans populations in Colombia: First report of the A2 mating type. Phytopathology 99(1): 82-88. [ Links ]
Xiben, W., E. Abdelbasset, R. Lorne, and D. Fouad. 2005. Genes encoding pathogenesis-related proteins PR-2, PR-3 and PR-9, are differentially regulated in potato leaves inoculated with isolates from US-1 and US-8 genotypes of Phytophthora infestans (Mont.) de Bary. Physiological and Molecular Plant Pathology 67: 49-56. [ Links ]
Zúñiga, L.N. 2000. Resistencia al tizón tardío de la papa (Phytophthora infestans) en cruzamientos de cultivares y clones de papa (Solanum tuberosum L.). Revista Mexicana de Fitopatología 18(1): 1-9. [ Links ]
