










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.66 no.1 Medellín Jan./June 2013
Acumulación y Distribución de Fitomasa en el Asocio de Maíz (Zea mays L.) y Fríjol (Phaseolus vulgaris L.)
Biomass Accumulation and Distribution in Associated Crop of Maize (Zea mays L.) and Bean (Phaseolus vulgaris L.)
Astrid Elena Pérez López1; Enrique Martínez Bustamante2; León Darío Vélez Vargas3 y José Miguel Cotes Torres4
1 Ingeniera Agrónoma. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias Agrarias - Departamento de Ciencias Agronómicas. A.A. 1779, Medellín, Colombia. <astridelenap@yahoo.com>
2 Profesor Asociado. Universidad Nacional de Colombia -Sede Medellín -Facultad de Ciencias Agrarias -Departamento de Ciencias Agronómicas. A.A. 1779, Medellín, Colombia. <enmartin@unal.edu.co>
3 Profesor Asociado. Universidad Nacional de Colombia -Sede Medellín -Facultad de Ciencias Agrarias -Departamento de Ciencias Agronómicas. A.A. 1779, Medellín, Colombia <ldvelez@unal.edu.co>
4 Profesor Asociado. Universidad Nacional de Colombia Sede Medellín, Facultad de Ciencias Agrarias -Departamento de Ciencias Agronómicas. A.A. 1779, Medellín, Colombia <jmcotes@bt.unal.edu.co>
Recibido: Diciembre 14 de 2011; aceptado: Marzo 13 de 2013.
Resumen
La asociación de cultivos se define como el crecimiento y desarrollo de dos o más especies en el mismo terreno, durante parte o todo el ciclo vegetativo. El asocio de maíz con fríjol afecta los atributos morfológicos y fisiológicos de ambas especies, lo que se manifiesta en la fitomasa acumulada en los distintos compartimentos y, finalmente, en los rendimientos. El objetivo de este trabajo fue determinar la acumulación y distribución de fitomasa de maíz y fríjol en asocio, respecto a sus unicultivos. Se sembraron las variedades: Fríjol Bola roja comercial (FBR), Cargamanto rojo (FC), una Línea Élite de Bola roja (FBE) y la variedad de maíz regional Montaña de grano blanco (M). Se evaluaron siete tratamientos: cuatro unicultivos de maíz y fríjol (Mo, FBRo, FCo, FBEo) y tres asociaciones (MxFBR, MxFC, MxFBE). El diseño estadístico fue de bloques completos al azar. Las variables peso seco de: tallos, hojas, mazorcas y panículas, en el maíz asociado con fríjol, disminuyeron respecto al unicultivo. La variedad de fríjol más afectada, negativamente, por el asocio fue FBE; en la cual, las variables peso seco de: tallos, hojas y vainas, se redujeron, en comparación con el unicultivo. La contribución de biomasa a la planta de fríjol, por parte de las vainas, es mayor en el asocio que en el unicultivo, excepto en la variedad FBE.
Palabras clave: Relación fuente - vertedero, materia seca, uso de la tierra, interacción fenotípica.
Abstract
Crop association is defined as the growth and development of two or more species in the same place during a part or all of the vegetative cycle. Maize and bean association affect morphological and physiological attributes of both species, which is manifested in the biomass accumulated in the different compartments and, finally, in the yields. The aim of this research was to determine the accumulation and distribution of both maize and bean biomass in association with regard to their unicultivos. Varieties planted were: "Bola roja commercial" (FBR), "Cargamanto rojo" (FC), an elite line "Bola roja" (FBE) and the maize variety "Montaña regional", white cornel (M). Seven treatments were evaluated: four unicultivos for maize and bean (Mo, FBRo, FCO, FBEo) and three associations (MxFBR, MxFC, MxFBE). The experimental design was a completely randomized block. The dry weight variables: stems, leaves, pods and panicles, in maize associated with bean, decreased compared to unicultivo. The bean variety most affected negatively by the association, was the elite ball (FBE), in which the dry weight variables: stems, leaves and pods, were reduced compared with the unicultivo. The contribution of biomass to the plant beans, by the pods was greater in the association that in the unicultivo, except for the elite ball bean (FBE) variety.
Key words: Relation source - sink, dry matter, land use, fenotypic interaction.
La asociación de cultivos se define como el crecimiento y desarrollo de dos o más especies en el mismo terreno, durante parte o todo el ciclo vegetativo; el asocio de maíz con fríjol voluble (MxFv) es un arreglo productivo constituido por dos especies contrastantes en sus características morfológicas y fisiológicas, sembradas simultáneamente y en el mismo sitio (Sullivan, 2003), cuyo crecimiento y desarrollo genera un microclima diferente al de sus respectivos unicultivos, el cual afecta los atributos morfológicos y fisiológicos de ambas especies, en diferentes intensidades, lo que finalmente se expresa en sus rendimientos (Acevedo, 1975; Vélez, 2009).
La asociación de plantas de diversas especies, al optimizar los recursos, genera ventajas agronómicas, tales como el uso intensivo de la tierra, la diversificación de cultivos y de fuentes de alimento y la disminución en la vulnerabilidad a factores bióticos y abióticos; además, se reduce el riesgo y la incertidumbre, tanto en la producción como en la comercialización. Las condiciones ecológicas, producto del asocio, tienen impacto positivo en la conservación y recuperación de los recursos naturales, la agrobiodiversidad y el control biológico (Francis, 1990; Tobón, 1990; Vandermeer, 1990; Vélez, 2009).
Para Román (1990), la distribución de fitomasa en fríjol está determinada por las siguientes reglas:
- Las diferentes partes de la planta compiten por recursos que casi siempre están en cantidades limitantes.
- Los diferentes órganos tienden a competir por estos recursos según el orden de prioridad: tejidos reproductivos (flores y vainas) - hojas - raíces - tallos.
- Las prioridades también dependen de los pesos relativos de los tejidos. Si un cultivo tiene un desarrollo foliar demasiado abundante, éste puede provocar una demanda suficientemente alta como para inhibir el crecimiento reproductivo.
- Bajo condiciones de demanda muy alta puede ocurrir remoción de nutrientes de un órgano hacia otro. Un ejemplo común es la remoción de carbohidratos de tallos hacia vainas.
En fríjol, la producción de biomasa es lenta entre los 15 y 71 días después de siembra; luego, se reduce la materia seca total ya que el aumento del peso de vainas no compensa la pérdida de biomasa por la caída de hojas en su proceso de defoliación natural (Muñoz, 1990).
Allen et al. (1976) demuestran, para un unicultivo de maíz, que las tasas más bajas de intercambio de CO2, vapor de agua y calor sensible ocurren en el tercio inferior del dosel del cultivo, donde la radiación fotosintéticamente activa limita más la fotosíntesis que la concentración de CO2. Esto contribuiría a explicar la reducción en la biomasa en ambas especies cuando se cultivan en asocio; sin embargo, el efecto en fríjol es mayor debido a su menor altura y por la disposición de su dosel con respecto al maíz, al concentrarlo en los tercios medio e inferior.
El objetivo de este trabajo fue determinar la acumulación y distribución de materia seca durante la ontogenia de plantas de maíz y fríjol en asocio, respecto a sus unicultivos.
Materiales y Métodos
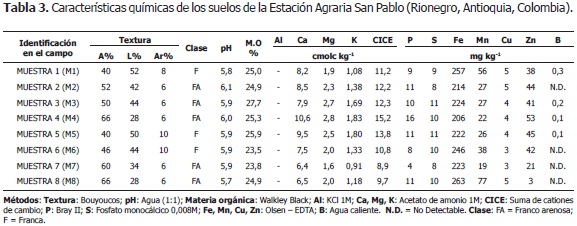
La investigación se realizó desde diciembre de 2007 hasta julio de 2008, en la Estación Agraria San Pablo de la Facultad de Ciencias Agrarias de la Universidad Nacional de Colombia, Sede Medellín, localizada en la vereda Tablacito, municipio de Rionegro, Antioquia (Colombia); cuyas coordenadas geográficas son: 06°07'52'' LN y 75° 27' 06'' LO; a una altitud de 2.100 msnm, temperatura media de 17 °C, 2.525 mm de precipitación y 75% de HR promedio, las que corresponden a la zona de vida: bosque muy húmedo Montano Bajo (bmh-MB), de acuerdo con el Sistema de Zonas de Vida (Holdridge, 1982). Los suelos de las parcelas son de textura franco arenosa, ácidos, pH de 5,7 - 6,0, contenido de materia orgánica de 23,5 - 7,7%; y capacidad de intercambio catiónico de 8,9 - 15,2 meq/100 g (Pérez, 2010).
Las variedades que se emplearon fueron: fríjol Bola roja comercial (FBR), Cargamanto rojo (FC) y una línea élite de Bola roja (FBE), la cual fue seleccionada por el Programa de Mejoramiento de la Universidad Nacional de Colombia - Sede Bogotá; y en maíz, la regional Montaña de grano blanco, línea 400.
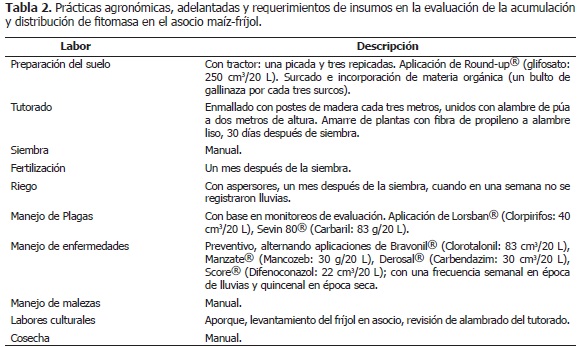
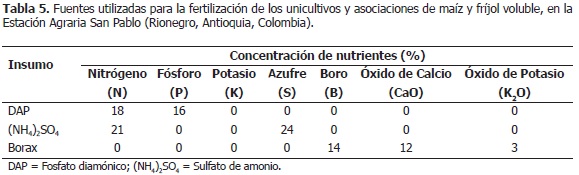
El experimento constó de siete tratamientos, los unicultivos de maíz (Mo) y fríjol voluble (FBRo, FCo, FBEo), y las siguientes asociaciones: maíz asociado con fríjol Bola roja (MxFBR); maíz asociado con fríjol Cargamanto (MxFC) y maíz asociado con fríjol Bola roja línea elite (MxFBE); los cultivos se establecieron en parcelas de 90 m2, cuya población y distribución de plantas en el campo se describe en la Tabla 1, con cuatro repeticiones, bajo un diseño de bloques completos al plagas y enfermedades (Tabla 2). La fertilización se azar para ejercer un mayor control sobre la variación realizó un mes después de la siembra, con base en del error ya que se manejaron dos especies diferentes el análisis de suelos (Tabla 3) y los requerimientos de de plantas en cuanto a arquitectura del dosel y las plantas según ICA (1992), conforme a lo cual se distintas variedades de fríjol; las cuales, conformaron ejecutó el plan nutricional presentado en la Tabla 4. unidades experimentales heterogéneas. Se realizaron Los fertilizantes utilizados fueron: fosfato diamónico las prácticas agronómicas estándar, para el cultivo en (DAP), sulfato de amonio ((NH4)2SO4) y bórax, los asocio y para el unicultivo, tales como preparación de cuales contienen la concentración de nutrimentos suelos, fertilización, labores culturales y manejo de detallada en la Tabla 5.
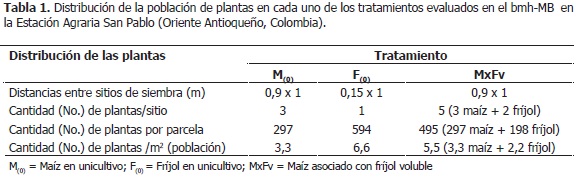

Para conocer la evolución de la fitomasa de los distintos órganos, quincenalmente se muestrearon, al azar y en total competencia (conforme a la población detallada en la Tabla 1), tres (3) plantas de maíz y dos (2) de fríjol por parcela, durante 30 y 22 semanas, respectivamente, para cada una de las especies mencionadas. Los diferentes órganos de las plantas se secaron hasta peso constante en un horno Thermolab® a 80 °C; luego se pesaron en una balanza de precisión Fisher Scientific® XL-4001. En la fitomasa del fríjol se consideraron: tallo (guías), hojas y vainas; para maíz: tallo, hojas, mazorcas e inflorescencia masculina (panícula).
Debido a que se realizaron mediciones de las variables varias veces en el tiempo, para el análisis estadístico se siguió un análisis de varianza de bloques divididos (Steel y Torrie, 1988), según el siguiente modelo:
Yijk =µ+ρi +α j +βk + (αβ )jk + (ρβ )ik +γi +δ j +λk +εijk
Donde,

Se utilizó el programa R, versión 2.11.0, para obtener las significancias de los efectos, se utilizó la prueba de tasa de máxima verosimilitud, usando una distribución chi-cuadrado. Se establecieron las diferencias estadísticamente significativas (Pérez, 2010), por medio de los intervalos de alta probabilidad a posteriori de 95%, para el valor medio de las variables de biomasa de fríjol y maíz.
Resultados y discusión
Acumulación de biomasa en maíz. En la Tabla 6, se presentan los coeficientes de determinación (R2) de los arreglos para las variables de biomasa en maíz. En los pesos secos de: hojas, tallo, mazorcas y panícula del maíz, se encontraron diferencias altamente significativas entre tratamientos (P<0,001). El peso seco de hojas en MxFBE (16,15 planta-1; semanas 6-12) fue mayor (P<0,05) que en Mo (13,23 g planta-1), cuando el maíz estaba en desarrollo de hojas y alargamiento del tallo; pero fue menor en floración y llenado de granos (MxFBE: 29,18 g; Mo: 32,49 g; semanas 16-26) (Figura 1A). En MxFBR, la fitomasa de hojas fue superior (28,09 g; semanas 9-16) que en Mo (24,87 g), durante el alargamiento del tallo y la emergencia de la panícula; mientras que fue inferior cuando el maíz estaba en llenado de granos (MxFBR: 28,75 g; Mo: 31,08 g; semanas 20-26) (Figura 1A). El MxFC no presentó diferencias significativas (p>0,05) en la fitomasa de hojas (27,42 g) respecto al unicultivo (26,21 g) (Figura 1A).).
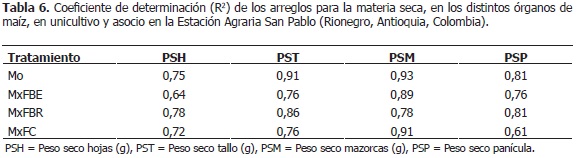
Se observó que en MxFBE y MxFBR, en comparación con Mo, el peso seco de hojas fue mayor en la fase vegetativa del maíz; tal vez, porque la planta aumentó la producción de fotoasimilados como respuesta al asocio con fríjol, pero se redujo cuando la planta estaba en llenado de granos; igualmente, se apreció en Mo y MxFC, donde esta variable disminuyó a partir de la semana 18 (Figura 1A); posiblemente, por la translocación de fotoasimilados desde las láminas foliares hacia otros órganos. Lo anterior es concordante con lo expresado por Vélez (2009) y Skinner (1991), quienes manifestaron que en asociación, las hojas de maíz acumulan menos biomasa que en unicultivo. Rajcan y Swanton (2001) infieren que el estrés por agua en maíz, causado por la interacción con arvenses, puede limitar la biomasa de las hojas; cuya competencia puede presentarse en el asocio con fríjol.
La biomasa del tallo en MxFBE (163,78 g) y MxFBR (162,32 g) fue menor (P<0,05) en relación con Mo (188,32 g), a partir de las semanas 15 y 17, respectivamente, en floración y llenado de granos en maíz; esto concuerda con Vélez (2009). En MxFC, fue mayor (133,52 g; semanas 13-17) que en unicultivo (120,91 g), en floración y estado lechoso del grano; pero, inferior a partir de la semana 21 (MxFC: 177,39 g; Mo: 204,74 g), en estado pastoso del grano (Figura 1B). Así, el asocio con fríjol redujo significativamente la fitomasa del tallo de maíz, durante el llenado de granos. Sin embargo, en MxFC disminuyó la biomasa del tallo a partir de la semana 20, posiblemente, por translocación de fotoasimilados a otros órganos; mientras que en MxFBE y MxFBR, aumentó, a pesar de ser menor que en Mo (Figura 1B).
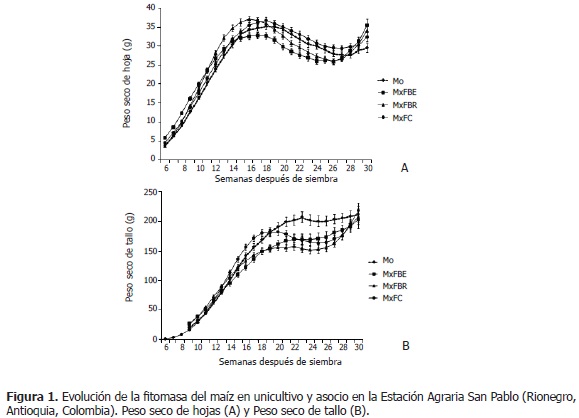
Esta situación debería verse reflejada en el peso seco de mazorcas, donde se esperaba que en MxFC fueran más pesadas que en los demás asocios, lo cual no ocurrió; pero la fitomasa de la panícula, sí fue mayor que en los demás arreglos, a partir de la semana 22 (Figura 2B); así, se puede inferir que la reducción de la biomasa del tallo en MxFC, podría ser causada por cambios en la relación fuente-vertedero, de tal manera que la translocación de fotoasimilados fueran direccionados hacia otros órganos; lo cual no se pudo clarificar con la información disponible. Desde la semana 27, cuando finalizó el llenado de granos, se observó en los asocios, más no en el unicultivo, un aumento en la fitomasa de hojas y tallo; tal vez, como respuesta del maíz a la no competencia con el fríjol, pues éste ya cumplió su ciclo de vida; así, el maíz cuenta con mayores recursos lumínicos y del suelo para aumentar la producción de fotoasimilados; ya que, si bien las plantas se pueden cosechar, sus hojas, especialmente las del tercio superior, siguen verdes.
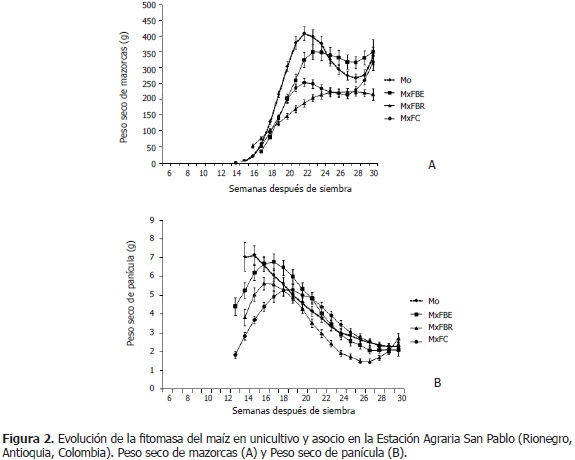
La materia seca de mazorcas fue reducida (P<0,05) en MxFC y MxFBR, en relación al unicultivo; mientras que en MxFBE, presentó superior fitomasa de mazorcas (322, 91 g; semanas 27-29) que en unicultivo (274,21 g) (Figura 2A); posiblemente, por una mayor translocación de fotoasimilados desde las hojas, lo cual podría evidenciarse con la mayor disminución de la biomasa de hojas (Figura 1A). Según Pérez (2010), la planta de fríjol en MxFBE, reduce las respuestas morfofisiológicas y de biomasa, lo que podría facilitar el mejor desarrollo del maíz; ello se evidencia en el superior peso de mazorcas, en comparación con los otros asocios. Por otra parte, se puede anotar que, tal como se comentó previamente, a partir de la semana 22 se apreció una reducción en la fitomasa de mazorcas y demás órganos aéreos a los que se les adelantó el seguimiento; para esta respuesta no se logró una explicación clara con la información obtenida; sin embargo, se sugiere que ha podido presentarse translocación de fotoasimilados hacia otros órganos (raíz, tallo, hojas, mazorca y sus componentes: pendúnculo, capacho, tuza y granos) o incrementos en los gastos energéticos de mantenimiento. Al respecto, Rajcan y Tollenaar (1999) expresaron que, durante el llenado del grano, los fotoasimilados pueden translocarse hacia las raíces, para la toma de nitrógeno necesario durante esta etapa fenológica. En este orden de ideas se tiene que, Borrás y Otegui (2001); Westgate et al. (2004) y Andrade et al. (2005), también apreciaron disminución de materia seca en los órganos, por lo que afirmaron que ello podría ser consecuencia de cambios en la relación fuente-vertedero. Este planteamiento es soportado por Borrás et al. (2004), quienes manifestaron que las disminuciones en la relación fuente-vertedero postfloración, causaron grandes reducciones en el peso final del grano; mientras que el aumento de esta relación tuvo efectos mínimos en el incremento del peso final del grano. Además, según Gambín et al. (2006) y Sala et al. (2007), un incremento en la relación fuente-vertedero durante la floración causa un aumento en el peso final del grano, debido a cambios en la tasa de crecimiento del grano durante su fase de llenado.
En la Figura 2B, se observó que el peso seco de panícula en unicultivo disminuyó en el tiempo, mientras que en los asocios, se incrementó durante la emergencia de ésta; tal vez, porque el asocio con fríjol, además de aminorar la producción de fotoasimilados, modificó la relación fuente-vertedero, de tal manera que también redujo la translocación de estos hacia la panícula, la cual perdió biomasa como consecuencia de la senescencia. En los asocios respecto al unicultivo, se encontró que el peso seco de panícula en MxFBE fue menor (P<0,05) en emergencia de ésta (5,29 g; Mo: 7,11 g; semanas 13-15), y mayor en el inicio del llenado de granos (MxFBE: 5,67 g; Mo: 4,87 g; semanas 18-21) (Figura 2B). En MxFBR, la variable siempre fue inferior comparado con Mo, aunque significativamente entre las semanas 14 y 16 (MxFBR: 4,84 g; Mo: 6,96 g), cuando la panícula emerge en la planta; como también entre las semanas 21 y 28 (2,16 g vs. 3,10 g en Mo), en el llenado de granos (Figura 2B). La biomasa de panícula en MxFC (3,54 g) fue menor que en Mo (6,74 g; semanas 13-17), en floración e inicio del llenado de granos; y superior entre las semanas 19 y 26, pero sólo significativamente en las semanas 21 y 22 (4,63 g vs. 3,99 g en Mo) (Figura 2B). Así, MxFv, respecto al unicultivo, reduce significativamente el peso seco de panícula, principalmente en la emergencia de ésta..
En resumen, MxFv redujo las variables de fitomasa en la planta de maíz. Estos resultados coinciden con los presentados por Díaz et al. (1995), donde el MxFv, acumuló 9% menos de biomasa en su parte vegetativa y 20% menos en grano, respecto al unicultivo.
Acumulación de biomasa en fríjol. Para los pesos secos de: hojas, tallo y vainas en fríjol, se encontraron diferencias altamente significativas entre los tratamientos (P<0,001). En la Tabla 7, se presentan los coeficientes de determinación (R2) de los arreglos, para las variables de biomasa en fríjol.
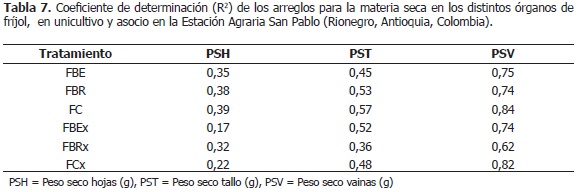
El peso seco de hojas en el fríjol Bola roja línea élite asociado con maíz (FBEx), logró en promedio 11,46 g durante todo el ciclo de vida; el cual, fue menor (P<0,05) al unicultivo (17,01 g); en fríjol Bola roja asociado con maíz (FBRx) fue inferior (10,21 g) que en fríjol Bola roja en unicultivo (FBRo) (13,42 g; semanas 8-12), en floración y crecimiento de vainas; pero superior, en llenado, maduración y cosecha de vainas (FBRx: 17,79 g; FBRo: 13,49 g; semanas 15-21); en fríjol Cargamanto asociado con maíz (FCx), la fitomasa de hojas fue menor que en fríjol Cargamanto en unicultivo (FCo), en el desarrollo de guías y floración (FCx: 8,12 g; FCo: 10,24 g; semanas 6-10), y mayor en llenado y maduración de vainas (FCx: 23,25 g; FCo: 19,44 g; semanas 14-18) (Figura 3A). En consecuencia se tiene que, con respecto a cada unicultivo, en FBRx y FCx la biomasa de las hojas fue menor al inicio de la fase reproductiva, específicamente en floración y crecimiento de vainas; pero, superior durante el llenado y maduración de estas últimas estructuras; sin embargo, la fitomasa de hojas de FBEx fue inferior durante todo el ciclo de vida; no obstante, Vélez (2009) no encontró diferencias en esta variable, entre el asocio y el unicultivo.
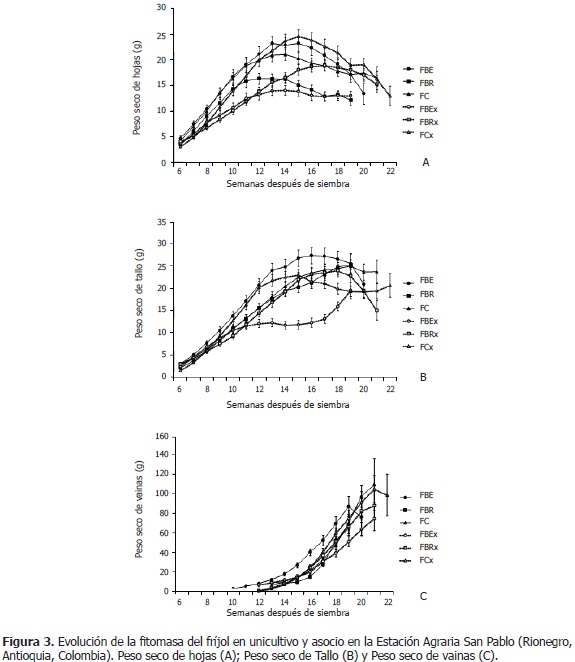
Con respecto a cada unicultivo, Pérez (2010) encontró en FBRx y FCx, una menor cantidad de hojas y área foliar al inicio de la fase reproductiva, pero al final de esta etapa fenológica la cantidad de láminas foliares y su área fue superior. Lo anterior, armoniza con lo hallado en este trabajo en cuanto a materia seca del follaje; lo que permite sugerir que la planta incrementó la cantidad de hojas para obtener un área foliar mayor, y así aumentar la producción de fotoasimilados y suplir la alta demanda energética que requería la construcción de cadenas de carbono durante esta etapa productiva. Finalmente, la mencionada investigadora expresó que FBEx con relación al unicultivo, redujo el número de hojas y el área foliar durante todo su ciclo de vida; por lo cual, las plantas disminuyeron la síntesis de sustancias, lo que se evidenció en la presente experiencia en una menor biomasa de hojas.
El peso seco del tallo en FBEx (11,01 g), es menor que en FBEo (18,84 g) (P<0,05); el FBRx, no presentó diferencias significativas en esta variable (15,21 g) respecto al unicultivo (15,15 g), lo que concuerda con lo dicho por Vélez (2009) (Figura 3B). En FCx, la variable fue mayor respecto al unicultivo (P<0,05), en el crecimiento de vainas (FCx: 19,57 g; FCo: 15,69 g; semanas 11-13), pero menor entre las semanas 17 y 20 (FCx: 20,03 g; FCo: 24,43 g); además, se observó que la variable desciende a partir de la semana 15, edad correspondiente al llenado, maduración y cosecha de vainas (Figura 3B). Se podría asumir que el peso seco del tallo en FCx, es superior que en FCo por una translocación de fotoasimilados desde las hojas, pero al observar la Figura 3A, entre las semanas 11 y 13, no se redujo la biomasa de hojas en el asocio respecto al unicultivo. También se podría pensar en el aumento de la cantidad de guías, pero según Pérez (2010), entre las semanas 11 y 13, el número de guías en FCx es inferior que en FCo. Se puede suponer entonces, que el tallo del FCx, como respuesta a la competencia con el maíz, podría realizar fotosíntesis para el crecimiento óptimo de las vainas.
El peso seco de vainas de las asociaciones, respecto a su unicultivo, presentó el siguiente comportamiento: en FBEx, fue menor (P<0,05) (FBEx: 32,49 g; FBEo: 36,31 g); en FBRx, fue mayor (FBRx: 25,27 g; FBRo: 17,32 g; semanas 15-17); y en FCx, fue superior (FCx: 50,55 g; FCo: 39,90 g; semanas 17-18); (Figura 3C). Al ser la fitomasa de vainas el componente principal en el rendimiento del fríjol (Denis, 1971), se esperó que en el asocio se redujera significativamente, como lo señalaron Tsubo et al. (2004); pero, este comportamiento sólo se presentó en FBEx, mientras que en FBRx y FCx no se disminuyó esta variable, posiblemente porque el número de vainas en FBRx y FCx es mayor que el de los unicultivos respectivos, entre las semanas 14 y 19 (Pérez, 2010); contrario a lo expuesto por Kohashi-Shiabata (1990), quien encontró un efecto negativo de la asociación en la formación de flores y vainas de fríjol. Además, se debe considerar que la biomasa de hojas en FBRx y FCx, también fue superior que en los unicultivos, por lo que podría traslocarse mayor cantidad de fotoasimilados a las vainas desde las hojas; igualmente, en FCx, como ya se discutió, se presenta una diferencia en la biomasa del tallo que podría favorecer el llenado de las vainas. También se tiene que, la etapa de crecimiento de vainas fue más larga en FBRx y FCx que en sus unicultivos (Pérez, 2010); ya que, los rendimientos en fríjol están más relacionados con la duración del llenado de grano que con su tasa, según Fukai y Trenbath (1993).
Sanabria (1975) explica que, si en un suelo no se presentaran limitaciones para absorber agua y nutrientes, la asociación de maíz por fríjol voluble (MxFv) beneficiaría al fríjol y sus rendimientos deberían ser mayores que en unicultivo, lo mismo que la biomasa de la planta, lo cual podría atribuirse al aumento del área foliar del fríjol debido al sombreamiento en el asocio. Según Ogindo y Walker (2005), MxFv no presentó diferencias significativas en la extracción total de agua durante las dos temporadas del ensayo; sin embargo, obtuvo mayor cantidad para conversión de biomasa. La absorción de agua es más intensa durante la floración del fríjol, cuando su consumo en la asociación es 30-40% mayor que en los unicultivos (Arango y Vásquez, 1979). La interacción genotipo por sistema es más marcada en fríjol que en maíz. Es así como, los genotipos de fríjol con altos rendimientos en unicultivos presentan tendencia a bajos rendimientos en asociación y viceversa (Casadiego y García, 1982; Davis y García, 1983; Angulo, 1986; Ligarreto et al., 1992).En resumen, se encontró que FBEx redujo de manera significativa los pesos secos de: hojas, tallo y vainas, respecto a su unicultivo. FBRx no presentó diferencias significativas con FBRo, en la biomasa del tallo; además, igual que en FCx, disminuyó la fitomasa de hojas en floración y crecimiento de vainas, pero la incrementó en llenado y maduración de frutos. Según Díaz et al. (1995) y Vélez et al. (2007), el fríjol asociado con maíz mermó su biomasa acumulada en más de 40% en su parte vegetativa y más de 60% en semilla, en comparación con el unicultivo.
Distribución de fitomasa en maíz. En Mo, entre las semanas seis y ocho, la biomasa de la planta la conformó el peso seco de hojas en un 100%, ya que el tallo fue perceptible a partir de la semana nueve. Entre las semanas 9 y 13, la fitomasa fue constituida por tallo en un 69% y por hojas en un 31%, en promedio. A partir de la semana 14, la planta irrumpe en estado reproductivo, donde emergen la panícula y las mazorcas. En este estado, el tallo pasa de ser el 72,4% al 36,7% de la biomasa total (-35,7 puntos) y las hojas del 21,7% al 5,0% (-16,7 puntos). La panícula inicia aportando el 5,6% a la materia seca total y termina en un 0,3% (-5,3 puntos); en cambio, las mazorcas comienzan siendo el 0,3% de la fitomasa de la planta y termina siendo el 58,0% (+57,7 puntos) (Figura 4A). Amador y Boschini (2000) advirtieron que antes de los 65 días, la acumulación de biomasa de hojas fue superior al de tallo, posteriormente se favoreció la acumulación en el tallo, superando cerca de tres veces la producción de fitomasa en hoja; las mazorcas tuvieron la mayor acumulación a los cuatro meses.
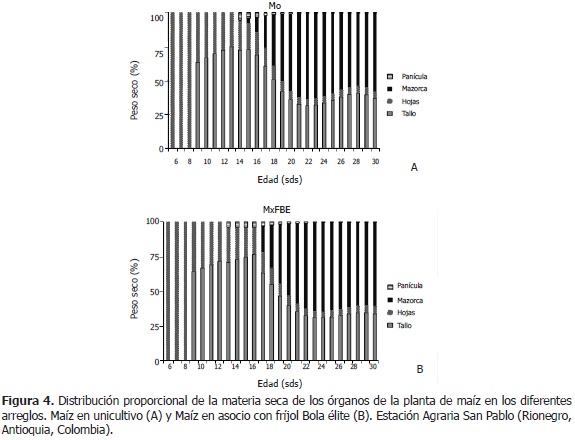
En MxFBE, entre las semanas seis y ocho, la biomasa total la conformó el peso seco de hojas en un 100%.
Entre las semanas 9 y 12, la fitomasa fue constituida por tallo, en un 68%, y por hojas, en un 32%, en promedio. El estado reproductivo, comenzó en la semana 13, donde el tallo participa inicialmente con un 70,9% de la biomasa de la planta y termina en la semana 30, con un 33,6% (-37,3 puntos); las hojas empiezan con un 25,3% y finalizan con un 5,6% (-19,7 puntos); la panícula inicia con 3,8% y acaba con un 0,3% (-3,5 puntos); las mazorcas principian con un 18,9% en la semana 17 y terminan con un 60,5% en la semana 30 (+60,5 puntos, desde la semana 13) (Figura 4B). El MxFBE, presentó el mismo comportamiento de distribución de fitomasa de sus órganos que el Mo.
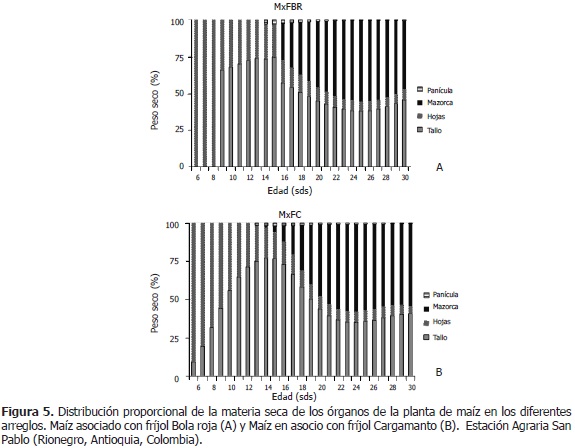
Para MxFBR, entre las semanas seis y ocho, la biomasa de la planta la conformó el peso seco de hojas en un 100%. Entre las semanas 9 y 13, la participación del tallo en la fitomasa total fue del 70%, mientras que la de hojas fue del 30%, en promedio. Desde la semana 14, la planta comenzó su estado reproductivo, distribuyéndose la biomasa de la siguiente manera: el tallo participó inicialmente con un 73,4% y terminó con un 45,6% (-27,8 puntos); las hojas, con un 23,8% al principio y un 7,2% al final (-16,6 puntos); la panícula, con un 2,8% al inicio y finalizó con un 0,5% (-2,3 puntos); las mazorcas, empezaron con un 24,9% en la semana 16 y acabaron con un 46,7% en la semana 30 (+46,7 puntos, desde la semana 14) (Figura 5A).
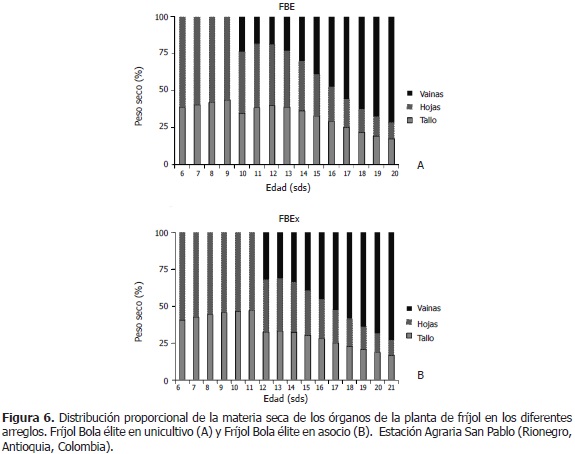
Distribución de fitomasa en fríjol. En FBEo, entre las semanas seis y nueve, la biomasa del vegetal fue constituida por el tallo (40%) y las hojas (60%); en la semana 10, se empezó a registrar el peso seco de vainas, el cual se incrementa desde un 23% hasta un 71% en la semana 20 (+48 puntos). En este lapso, las hojas pasan de un 42% a un 11% (-31 puntos) de aporte a la fitomasa total, mientras el tallo pasa de un 35% a un 18% (-17 puntos) (Figura 6A). En FBEx, entre las semanas 6 y 11, la biomasa de la planta se conformó por un 44% de tallo y un 56% de hojas, en promedio. A partir de la semana 12 hasta la 21, el peso seco de vainas pasó del 32% al 73% (+41 puntos); el de hojas, del 36% al 10% (-26 puntos) y el de tallos, del 32% al 17% (-15 puntos) (Figura 6B).
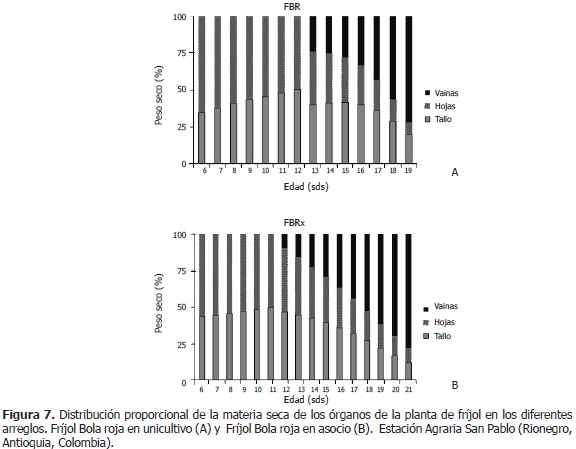
En FBRo, las hojas aportaron un 43% y los tallos un 57% de la biomasa total, en promedio, desde la semana 6 hasta la 12. Entre la semana 13 y 19, las hojas disminuyeron su aporte en 28 puntos (de 36% a 8%); los tallos, en 20 puntos (de 40% a 20%); las vainas aumentaron su aporte en 48 puntos (24% a 72%) (Figura 7A). En FBRx, desde la semana 6 hasta la 11, las hojas aportaron a la biomasa total, un 54%, mientras que el tallo, un 46%, en promedio. Entre las semanas 12 y 21, las hojas pasaron de aportar un 44% a un 10% (-34 puntos), mientras que el tallo, pasó de un 46% a un 12% (-34 puntos). Las vainas pasaron de contribuir el 10% de la fitomasa total, al 78% (+68 puntos) (Figura 7B).
El FCo, entre las semanas 6 y 12, presentó la siguiente distribución de materia seca: hojas, 59%; tallo, 41%, en promedio. Durante las semanas 13 y 21, las hojas participaron con un 47% inicialmente y terminaron con un 9% (-38 puntos); los tallos, con un 43% al comienzo y un 13% al final (-30 puntos). Las vainas integraron el 10% de la biomasa al inicio y el 78% en la última semana (+68 puntos) (Figura 8A). La distribución de fitomasa en FCx, fue de la siguiente manera: entre las semanas 6 y 11, las hojas proporcionaron a la planta, el 57% de biomasa, mientras que el tallo, el 43%. Entre las semanas 12 y 22, las hojas disminuyeron su porcentaje de 48 a 8 (-40 puntos); los tallos redujeron su proporción de 48% a 12% (-36 puntos) y las vainas, aumentaron su participación de un 4% a un 80% (+76 puntos) (Figura 8B).
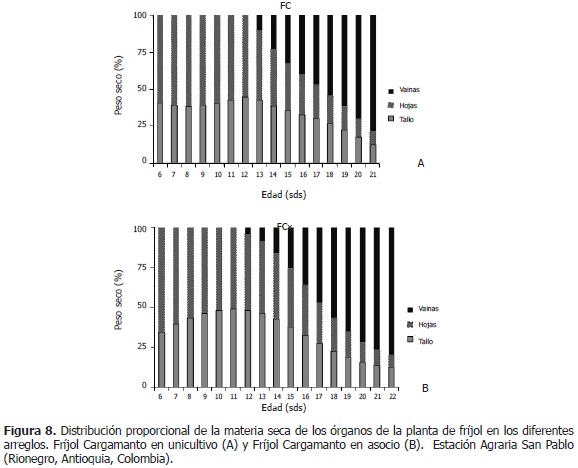
En resumen, en los distintos arreglos, en la fase vegetativa, las hojas participaron con una mayor contribución a la biomasa total, en comparación con el tallo; excepto en FBRo, en el cual el tallo tuvo una mayor participación comparado con las hojas. En el periodo reproductivo del fríjol, las hojas aportaron el mayor porcentaje de fitomasa a las vainas, en relación con los tallos. En FBRx, el aporte fue igual (34 puntos porcentuales). En la última semana del ciclo de vida del fríjol, las vainas lograron alcanzar entre el 71% y el 80% de la biomasa total, aumentando su aporte desde 41 puntos en FBEx hasta 76 puntos en FCx. Las vainas participaron con un mayor porcentaje en los asocios que en los unicultivos, excepto en FBEx, el cual tuvo una menor participación de vainas en la fitomasa del vegetal. Vélez (2009), encontró que la masa acumulada del tallo y las hojas en fríjol asociado, no fue significativamente diferente a la del unicultivo, mientras que la fitomasa de vainas fue menor en el asocio que en el unicultivo.
Conclusiones
En maíz asociado con fríjol, todas las variables de materia seca disminuyeron.
La variedad de fríjol más afectada negativamente por el asocio fue la de Bola élite, en la cual todas las variables de fitomasa se redujeron, en comparación con el unicultivo. Entre las variedades asociadas, en todas las variables de biomasa, el fríjol Cargamanto presentó los mayores valores.
La contribución de fitomasa a la planta de fríjol, por parte de las vainas, es mayor en el asocio que en el unicultivo, excepto en la variedad fríjol Bola élite, lo que se podría atribuir a efectos del sombreamiento en el asocio y a la adecuada disponibilidad de nutrientes en el suelo para el cultivo, por lo que se infiere que en este asocio, predominó la competencia por luz, más que por agua y nutrientes.
Agradecimientos
A la Vicerrectoría de Investigación y a la Dirección de Investigación Medellín - DIME, por la financiación, al Grupo de Investigación ECOFIAGRO, del Proyecto "Relaciones de Competencia en Agroecosistemas: Evaluación Ecofisiológica del Cultivo Asociado Maíz - Fríjol Voluble Trepador". A Dora Villegas, del Laboratorio de Fisiología Vegetal; Walter Osorio y Alejandro Londoño, del Laboratorio de Microbiología de Suelos y a Jorge Andrés Pérez, del herbario MEDEL. También a Edwin González, Claudia Uribe, Alexander Aguirre, Yaned Zapata y especialmente a Mauricio Valencia.
Bibliografía
Acevedo, S. 1975. Influencia de la radiación solar y otros componentes del microclima sobre el cultivo del fríjol (Phaseolus vulgaris L.) asociado con maíz (Zea mays L.). Tesis Magister. Universidad de Costa Rica. Centro Agronómico Tropical de Investigación y Enseñanza. Departamento de Cultivos y Suelos Tropicales. Turrialba, Costa Rica. 100 p. [ Links ]
Allen, L.H., T.R. Sinclair and E.R. Lemon. 1976. Radiation and microclimate relationship in multiple cropping systems. pp. 171-200. In: Multiple cropping. Matthias Stelly (ed.). ASA Special Publication No. 27. American Society Agronomic, Crop Science Society of America and Soil Science Society of America. USA. 378 p. [ Links ]
Amador, R. y F.C. Boschini. 2000. Fenología productiva y nutricional de maíz para la producción de forraje. Agronomía Mesoamericana 11(1): 171-177. [ Links ]
Andrade, F.H., V.O. Sadras, C.R. Vega and L. Echarte. 2005. Physiological determinants of crop growth and yield in maize, sunflower and soybean. Journal of Crop Improvement 14(12): 51-101. [ Links ]
Angulo, N. 1986. Comportamiento agronómico de selecciones avanzadas de fríjol voluble con maíces en asocio y en unicultivo. Trabajo de Grado Ingeniero Agrónomo. Facultad de Agronomía. Universidad Nacional de Colombia. Bogotá. 89 p. [ Links ]
Arango, O. y F. Vásquez. 1979. Evaluación de la competencia agronómica ejercida por el maíz ICA H-210 sobre cuatro materiales de fríjol en asociación. Trabajo de Grado Ingeniero Agrónomo. Facultad de Agronomía. Universidad de Caldas. Manizales. 125 p. [ Links ]
Borrás, L. and M.E. Otegui. 2001. Maize kernel weight response to postflowering source-sink ratio. Crop Science 41(6): 1816-1822. [ Links ]
Borrás, L., G.A. Slafer and M.E. Otegui. 2004. Seed dry weight response to source-sink manipulations in wheat, maize and soybean: a quantitative reappraisal. Field Crops Research 86(2-3): 131-146. [ Links ]
Casadiego, E. y J. García. 1982. Evaluación agronómica de veinticinco variedades de maíz (Zea mays L.) de clima frío bajo el sistema de asociación con fríjol voluble. Trabajo de Grado Ingeniero Agrónomo. Facultad de Agronomía. Universidad Nacional de Colombia. Bogotá. 164 p. [ Links ]
Davis, J. y S. García. 1983. Interacción de genotipos por sistema de cultivo en fríjol - maíz. Programa de Fríjol. Curso de Fríjol. CIAT. Cali, Colombia. 8 p. [ Links ]
Denis, A. 1971. Análisis factorial de las variables del tipo de planta, relacionados con el rendimiento del fríjol (Phaseolus vulgaris L.). Tesis (Ph. D.) Michigan State University. Departament of Crop and Soil Sciences. [ Links ]
Díaz, C., J. Rivera y J. Quirós. 1995. Crecimiento y desarrollo del maíz y fríjol trepador en unicultivo y asociación. Actualidades CORPOICA 9(9): 5-10. [ Links ]
Francis, C.A. 1990. Potential of Multiple Cropping Systems. pp. 137- 150. In: Altieri, M.A. and S.B. Hecht. (eds.). Agroecology and Small Farm Development. CRC Press, Boca Raton. 262 p. [ Links ]
Fukai, S. and B. Trenbath. 1993. Processes determining intercrop productivity and yields of components crops. Fields Crops Research 34(3-4): 247-271. [ Links ]
Gambín, B.L., L. Borrás and M. Otegui. 2006. Source-sink relations and kernel weight differences in maize temperate hybrids. Field Crops Research 95: 316-326. [ Links ]
Holdridge, L. R. 1982. Ecología basada en zonas de vida. (Traducción del inglés por Humberto Jiménez Saa). Primera edición. IICA, San José, Costa Rica. 216 p. [ Links ]
Instituto Colombiano Agropecuario ICA. 1992. Fertilización en diversos cultivos: quinta aproximación. Bogotá. 64 p. [ Links ]
Kohashi-Shiabata, J. 1990. Aspectos de la morfología y fisiología del fríjol (Phaseolus vulgaris L.) y su relación con el rendimiento. Centro de Botánica, Colegio de Postgraduados, Chapingo, Montecillos, México. 44 p. [ Links ]
Ligarreto, G., G. Bastidas y M. Castillo. 1992. Efectos de las densidades de siembra en el rendimiento de la asociación maíz-fríjol voluble. Revista ICA 27: 25-33. [ Links ]
Muñoz, R. 1990. Características de los suelos y la fertilización del fríjol en Colombia. Conferencia presentada en el Curso Nacional de Fríjol. C.I. La Selva, Rionegro, Antioquia, sp. [ Links ]
Ogindo, H.O. and S. Walker. 2005. Comparison of measured changes in seasonal soil water content by rainfed maize-bean intercrop and component cropping systems in a semi-arid region of southern Africa. Physics and Chemistry of the Earth 30: 799-808. [ Links ]
Pérez, L. 2010. Evaluación ecofisiológica del cultivo asociado maíz (Zea mays L.) -fríjol voluble trepador (Phaseolus vulgaris L.). Tesis Magister en Ciencias Agrarias. Facultad de Ciencias Agropecuarias. Universidad Nacional de Colombia, Medellín. 92 p. [ Links ]
Rajcan, I. and M. Tollenaar. 1999. Source: sink ratio and leaf senescence in maize: I. Dry matter accumulation and partitioning during grain filling. Field Crops Research 60(3): 245-253. [ Links ]
Rajcan, I. and C.J. Swanton. 2001. Understanding maize-weed competition: resource competition, light quality and the whole plant. Field Crops Research 71(2): 139-150. [ Links ]
Román, V. 1990. Razas de fríjol común. Conferencia presentada en el Curso Nacional de Fríjol. C.I. La Selva, Rionegro, Antioquia. sp. [ Links ]
Sala, R.G., M.E. Westgate and F.H. Andrade 2007. Source/sink ratio and the relationship between maximum water content, maximum volume, and final dry weight of maize kernels. Field Crops Research 101: 19-25. [ Links ]
Sanabria, E. 1975. Producción de biomasa, nutrición mineral y absorción de agua en la asociación maíz - fríjol cultivada en solución nutritiva. Tesis de Magister. Instituto Interamericano de Ciencias Agrícolas de la OEA. Centro Tropical de Enseñanza e Investigación. Departamento de Cultivos y Suelos Tropicales. Turrialba, Costa Rica. 80 p. [ Links ]
Skinner, E. 1991. Relaciones alométricas de diez genotipos de maíz asociados y no asociados con fríjol voluble. Trabajo de Grado Ingeniero Agrónomo. Facultad de Agronomía. Universidad Nacional de Colombia. Bogotá. 106 p. [ Links ]
Steel, R.G. y J.H, Torrie. 1988. Bioestadística: principios y procedimientos. Segunda edición. McGraw-Hill, México, D.F. 622 p. [ Links ]
Sullivan, P. 2003. Intercrop principles and practices (Agronomy Systems Guide). ATTRA- National Sustainable Agriculture Information Service. In: http://www.attra.org/attra-pub/intercrop.html. 12 p.; consulta: enero 2011. [ Links ]
Tobón, C. 1990. La asociación de leguminosas en Colombia. Conferencia presentada en el Curso Nacional de Fríjol. C.I. La Selva, Rionegro, Antioquia. 13 p. [ Links ]
Tsubo, M., H.O. Ogindo and S. Walker. 2004. Yield evaluation of maize-bean intercropping in a semi-arid region of South Africa. African Crop Science Journal 12(4): 351-358. [ Links ]
Vandermeer, J.H. 1990. Intercropping. pp. 481-516. In: Carroll, C.R., J.H. Vandermeer and P. Rosset. (eds.). Agroecology. Mc. Graw Hill, New York. 641 p. [ Links ]
Vélez, L.D. 2009. Evaluación ecofisiológica del cultivo asociado Maíz (Zea mays L.) - fríjol trepador (Phaseolus vulgaris L.). Tesis Doctoral en Ciencias Agropecuarias - Área Agraria. Facultad de Agronomía. Universidad Nacional de Colombia. Bogotá. 346 p. [ Links ]
Vélez, L.D., J. Clavijo y G.A. Ligarreto. 2007. Análisis ecofisiológico del cultivo asociado maíz (Zea mays L.) - fríjol voluble (Phaselus vulgaris L.). Revista Facultad Nacional de Agronomía-Medellín 60(2): 3965-3984. [ Links ]
Westgate, M.E., M.E. Otegui and F.H. Andrade. 2004. Physiology of the Corn Plant. pp. 235-271. In: Wayne Smith, C., J. Betrán and E.C. Runge. (eds.), Corn: origin, history, technology and production. John Wiley and Sons, Inc., Hoboken, New Jersey, 949 p. [ Links ]
