










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.66 no.2 Medellín July/Dec. 2013
Dinámica de las Poblaciones de Lombrices en un Andisol Sometido a Distintos Sistemas de Uso del Suelo
Population Dynamics of Earthworms in an Andisol under Different Soil Use Systems
Ramiro Ramírez Pisco1; María Elena Guzmán Álvarez2 y Edna Ivonne Leiva Rojas3
1 Profesor Asociado. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias. Calle 59 A No. 63-20, Medellín, Colombia. <rramirez@unal.edu.co>
2 Ingeniera Agrónoma. Unión Temporal Gestión Ambiental y Patrimonial - UTGAP. Universidad Nacional de Colombia - Sede Medellín. Medellín, Colombia. <meguzman2009@hotmail.com>
3 Profesora Asociada. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias Agrarias. Calle 59 A No. 63-20, Medellín, Colombia. <eileiva@unal.edu.co>
Recibido: junio 18 de 2012; aceptado: marzo 31 de 2013.
Resumen. Se evaluó la composición taxonómica de la población de lombrices en un Andisol en la vereda Montañitas del municipio Marinilla (Antioquia, Colombia) luego de 5 y 10 años de labranza, después de 5 y 10 años de barbecho y en condiciones prístinas. El muestreo fue realizado por el método del monolito, propuesto por el programa Tropical Soil Biology and Fertility (TSBF). Se realizó extracción directa de los individuos, los cuales fueron caracterizados y descritos según los géneros encontrados en cada condición, para luego relacionarlos con los usos del suelo; fueron encontrados tres géneros: Pontoscolex corenthrurus, Amynthas corticis y Martiodrilus sp. En el suelo virgen se presentó P. corenthrurus, pero luego de 5 años de labranza se evidenció un predominio del género A. corticis y una baja presencia del género Martiodrilus sp. Luego de 10 años de labranza no se encontraron anélidos; sin embargo, durante el barbecho la población de lombrices incrementó en forma significativa. Los agregados estables del suelo disminuyeron en la medida en que el suelo fue sometido a labranza, y se incrementó durante los periodos de barbecho, asociado a la densidad de lombrices y a su concentración en los primeros 10 cm de profundidad, corroborando su participación en la agregación del suelo.
Palabras clave: Oligocheta, labranza, barbecho, estructura del suelo.
Abstract. The taxonomic composition of the population of earthworms in Andisol was evaluated on the rural area of Montañitas, Marinilla municipality (Antioquia, Colombia), under different conditions: after 5 and 10 years of farming, after 5 and 10 years of fallow as well as under pristine conditions. The sampling was performed by using the method of the monolith, proposed by the program Tropical Soil Biology and Fertility (TSBF). The specimens were obtained by direct extraction from each sample, which then were characterized and described according to genera found under each condition, and then relate land usage. Three different genera of earthworms were found: Pontoscolex corenthrurus, Amynthas corticis and Martiodrilus sp. In the virgin soil P. corenthrurus was present, the soil with five years of tillage showed a predominance of A. corticis and low presence of Martiodrilus sp. In soils after 10 years of tillage no annelid was found; however, during the fallow, earthworm population increased significantly. It was found that the stable aggregates decreased depending on the duration of soil tillage, that it increased during fallow periods associated with the density of earthworms and their concentration in the first 10 cm of depth, confirming their involvement in the aggregation of soil.
Key words: Olygocheta, , tillage, fallow, soil estructure.
Generalmente los suelos agrícolas no son hospitalarios para la macrofauna, ya que se mantienen productivos mediante los procesos de labranza y la aplicación de fertilizantes (inorgánicos y orgánicos), agroquímicos y encalamiento. Es ampliamente reconocido que las lombrices de tierra son uno de los grupos de macroinvertebrados más importantes en el suelo, destacándose por los significativos efectos sobre el medio edáfico a partir de su acción excavadora y porque mezcla de restos vegetales con el material inorgánico del suelo, estimulando la población microbial (especialmente de hongos) mediante la deposición de moco de su tracto digestivo. Estas acciones conducen a la formación de microagregados estables que contienen subproductos de excreción llamados coprolitos, los cuales generan una estructura migajosa muy estable dentro del suelo. Una vez estabilizados estos microagregados fecales dentro de los macroagregados del suelo, estos pueden funcionar como micrositios anaeróbicos propios para altos niveles de desnitrificación (Burbano, 1998; Six et al., 2000; Jongmans et al., 2001).
Las poblaciones de lombrices de tierra en suelos cultivables dependen, en gran medida, de las prácticas de manejo empleadas, ya que estas generan disturbios sobre y debajo de la superficie del suelo afectando los organismos que lo habitan. La diversidad edáfica debe exceder un mínimo para mantener la estabilidad del ecosistema. La desaparición de una sola especie clave, que no se compensa, se puede manifestar súbitamente con el colapso del ecosistema (Lee, 1985; Edwards y Bohlen, 1996; Benckiser, 1997).
Otro efecto importante producido por las lombrices, es que promueven la aireación y porosidad a través de la formación de macroporos continuos, e incrementan la proporción de grandes agregados en el suelo, efecto especialmente importante en suelos con estructura pobre. Al aumentar la tasa de infiltración de agua, las lombrices pueden reducir la pérdida de suelo y se aumenta el aprovechamiento del riego en los cultivos (Lee, 1985; Lavelle et al., 1998).
Según Lavelle (1997), las actividades desarrolladas por las lombrices de tierra afectan la producción de la biomasa vegetal tanto del área visible, como del área radical (no visible). Estas actividades son producidas por dos grandes grupos funcionales de lombrices: el de los "transformadores del mantillo" y el de los "ingenieros del ecosistema" (Jones et al., 1994). Las lombrices "transformadoras del mantillo" son las epigeas, y se encuentran dentro de la hojarasca en la superficie del suelo (Bouché, 1984). Sus actividades se reducen a unos pocos centímetros en la capa superior, aunque son vitales para una gran parte de la microflora y fauna edáficas. Las epigeas desmenuzan la materia orgánica vegetal, modificando sus propiedades físico-químicas, facilitando su disposición a la microflora para una posterior descomposición (Lavelle, 1997). Las lombrices consideradas "ingenieros del ecosistema", las endogeicas y anécicas, afectan la disponibilidad de recursos por parte de otros organismos mediante la modificación del ambiente físico (Jones et al., 1994).
Muchas de las especies de lombrices de tierra nativas no pueden tolerar los cambios en el hábitat del suelo causados por las prácticas agrícolas, lo que conduce a la dominación de unas pocas especies exóticas (Lavelle y Pashanasi, 1989; Fragoso y Lavelle, 1992; Fragoso et al., 1995; Edwards y Bohlen, 1996; Lavelle et al., 1998).
En el presente estudio, se evaluó la agregación y la presencia de las diferentes lombrices de tierra encontradas en historiales de manejo de un Andisol. Se efectuaron las identificaciones de los ejemplares extraídos y se realizó una breve descripción de cada uno.
MATERIALES Y MÉTODOS
Área de estudio. La colecta se realizó en el municipio de Marinilla (Antioquia, Colombia) ubicado a 45 km del oriente de la ciudad de Medellín, a 6° 09' 50,05 N y 75°17'15,8 O en la vereda el Montañitas, a una altitud de 1.800 m con una precipitación promedio anual de 1400 mm y una temperatura promedio de 18 °C. Corresponde a zona de bosque húmedo montano bajo (bh-MB) según la clasificación propuesta por Holdridge. La distribución de la precipitación es bimodal, con presencia de dos periodos de lluvia inter semestrales (marzo-mayo y septiembre-noviembre) (CORNARE, 1995).
Los suelos estudiados correspondieron a un Andisol con diferentes historias de manejo las cuales correspondieron a: suelo virgen, suelo con 10 y 5 años de barbecho, y suelo bajo 5 y 10 años de labranza convencional (LC), dedicados a la producción intensiva de hortalizas. Los suelos fueron caracterizados químicamente y además se determinó la agregación por el método propuesto por De Leehneer y De Boodt (1958) modificado por Ramírez (2008).
Diseño de muestreo y variables. Los muestreos de suelo se efectuaron con el método recomendado por el programa Tropical Soil Biology and Fertility (TSBF) (monolitos de 25x25x30 cm), y se hizo extracción manual de los ejemplares de lombrices. En cada tipo de manejo del suelo se colectaron 5 monolitos. Las lombrices se llevaron inmediatamente al laboratorio para su separación, identificación y cuantificación.
Diagnosis de las lombrices recolectadas. Los especimenes fueron extraídos, lavados y posteriormente sacrificados con una solución de alcohol al 10%. Después de 12 h, se utilizó formol al 4% como solución fijadora, y para la conservación se empleó alcohol al 70% (Darrigran et al., 2007). Se separaron debidamente por morfoespecies y de cada una se tuvo en cuenta la madurez sexual, la longitud total, el número de segmentos totales, el ancho anterior al segmento VI o VII, el ancho posterior al segmento X, el color del dorso y vientre, la forma del prostomio, y el largo y ancho (mm) del prostomio, se empleó un microscopio óptico.
Se extrajeron las setas dorsales de la parte media de cada espécimen y luego fueron colocadas en placas portaobjetos, fueron observadas y fotografiadas en microscopio. También, de cada morfoespecie, se definió la forma de los poros ventrales (ovalado, redondeado, entre otros), el segmento en que se encontraron (XVII, XVIII, XIX) y se describieron de acuerdo a la posición y el tamaño.
RESULTADOS
Condiciones del suelo. La labranza afectó las condiciones de agregación del suelo de manera significativa, disminuyendo el diámetro medio ponderado en húmedo (DMPh) en la medida en que se incrementó el periodo de labranza y durante el barbecho. El contenido de materia orgánica presentó una estrecha relación con el DMPh. Con el paso de este suelo a barbecho, se incrementó la materia orgánica alcanzando casi el 60% del contenido del suelo virgen, lo cual se logra a partir de la contribución permanente de la hojarasca de arbustos y las raíces de las gramíneas de la sucesión secundaria; además, disminuyó el contenido de N-NO3, Ca, Mg, K y P, debido a que en el descanso no se aportan fuentes de estos minerales, asemejándose a las condiciones del suelo virgen (Tabla 1).
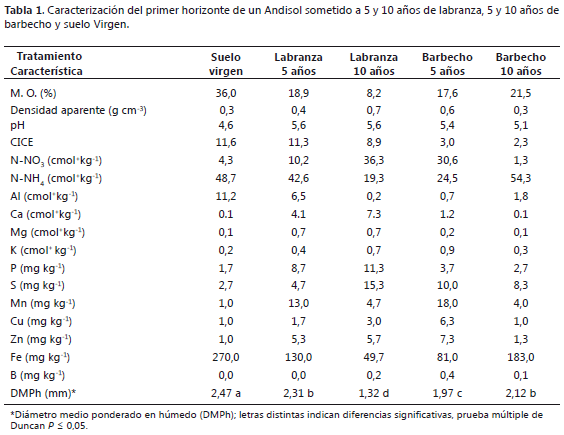
Diagnosis de la población de lombrices. La población de lombrices en suelo virgen correspondió a 160 individuos m-2; este valor se redujo al 73% luego de 5 años de labranza, y no se observaron individuos después de 10 años; no obstante, al pasar 5 años el suelo en barbecho la población se duplicó con respecto al suelo prístino, expresando así su gran capacidad de colonización, asociada a la alta disponibilidad de raíces, las cuales son comunes en las primeras fases de la sucesión vegetal secundaria desarrollada en el barbecho; resultados similares fueron reportados por Coral y Bonilla (1998) bajo presencia de gramíneas. Luego de 10 años de barbecho, la población de lombrices disminuyó, pero continuó siendo un 10% más alta que en suelo virgen (Tabla 2).

Se encontraron tres especies de oligoquetos, entre ellas dos de la familia Glossoscolecidae (Martiodrilus sp, y Pontoscolex corethrurus) y una de la familia Megascolecidae (Amynthas corticis). Solo la especie Pontoscolex corethrurus se encontró en el suelo prístino.
Martiodrilus sp. En total fueron 12 especímenes revisados minuciosamente, este género sólo se halló en el suelo con 5 años de labranza.
Descripción: Es una especie anécica con una longitud entre 40-95 mm, con un color más oscuro en la parte dorsal que en la parte ventral. Los especímenes encontrados tuvieron: ancho en segmentos VI-VII de 1-3,1 mm; ancho posterior al último segmento de 1-2,8 mm. El número de segmentos del cuerpo varió entre 71-111 anillos. El prostomio presentó características redondeadas en todos los especímenes, con 0,1-0,4 mm de largo y 0,1-0,4 mm de ancho. Todos los especímenes fueron dorsalmente y ventralmente de color blanco o pálido, con poros laterales (lejos de una línea media ventral), uno en cada lado, ubicados en el segmento XVII en forma de cono invertido.
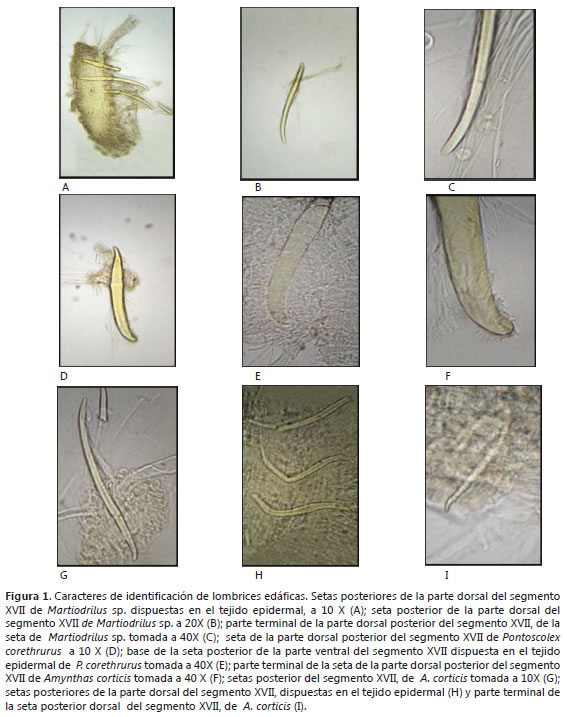
Los individuos exhibieron setas pequeñas de forma redondeada en la parte terminal y puntiaguda en la base de forma curva (Figura 1 A-C), dispuestas en un continuo anillo organizadamente en filas alrededor de cada segmento. También se observaron setas gruesas en cada segmento, se puede sentir frotando el cuerpo de la lombriz de cola a la cabeza. El clitelo se encuentra entre el segmento XVIII-XX.
Pontoscolex corethrurus (Müller, 1856). En total fueron 23 especímenes revisados detalladamente; se extrajeron del suelo con 5 años de labranza, con barbecho y del suelo virgen.
Descripción: Esta es una especie tropical, común en pastos y selvas. Se encontró a una profundidad de 0-10 cm, esta fue la única especie que se halló en suelo virgen. El cuerpo es pálido y sin pigmentación, puede haber un ligero abultamiento en el cuerpo alrededor de un tercio de la longitud de la cola. Los especímenes tuvieron una longitud 22-51 mm; ancho en segmentos VI-VII de 2-3,1mm; ancho posterior al último segmento de 1,1-3 mm. Número de segmentos entre 71-181. Prostomio invaginado en todos los especímenes. Todos los especímenes dorsalmente y ventralmente de color blanco o pálido. No hay poros dorsales bien definidos. Setas muy visibles, curvadas tanto en la parte terminal como en la base y redondeadas, un poco más delgada en la base (Figura 1 D-F), no emparejadas, dispuestas aleatoriamente alrededor de cada segmento en hileras regulares y parejas en la parte anterior del cuerpo, pero cambia a un patrón diferente, en el cual las setas de cada uno de los segmentos son equidistantes entre sí alrededor de la circunferencia del cuerpo. Setas muy gruesas en cada segmento, se puede sentir frotando el cuerpo de la lombriz de cola a la cabeza. El clitelo se encuentra entre el segmento XV-XX y se interrumpe en la parte ventral.
Amynthas corticis (Kinberg, 1867). Fueron en total 23 especímenes revisados detalladamente, se extrajeron del suelo con 5 años de labranza y con barbecho.
Descripción: Pertenece la familia Megascolecidae. Esta especie tropical, acorde con las características de la familia, son de color castaño rojizo en la parte dorsal y blanquecina en la parte ventral. Es la especie más abundante en el suelo laboreado. Los especímenes encontrados tuvieron una longitud de 28-74 mm; ancho en los segmentos VI-VII de 1-2,1 mm; ancho posterior al último segmento 0.8-2,1 mm. Número de segmentos entre 83-115. Prostomio redondeado en todos los especímenes con 0,2-0,5 mm de largo y 0,2-0,4 mm de ancho. Todos los especímenes dorsalmente de color marrón y ventralmente de color blanco o pálido. Poros masculinos ventrales (cerca a una línea media ventral); 2 poros rodeados de hendiduras concéntricas ubicados en el segmento XVIII, de forma ovalada. Setas pequeñas, muy delgadas como cerdas (Figura 1 G-I), con una ligera curva en la parte terminal y en la base, dispuestas en hileras regulares organizadas en cada segmento. Tiene numerosas setas por cada segmento. El clitelo anular ocupa los segmentos XIV-XVI.
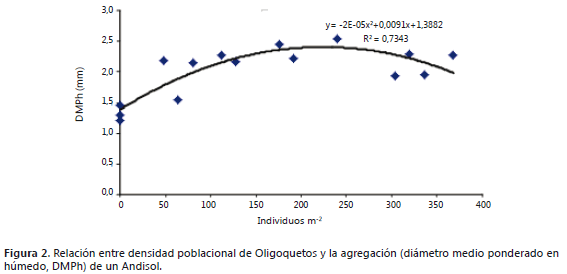
Agregación y oligoquetos edáficos. La población de lombrices presentó notable correspondencia con el tamaño de los agregados, hubo alta correlación entre la densidad de lombrices y el diámetro medio ponderado en húmedo (DMPh, R2 = 0,7343) validando la participación de esta edafofauna en la formación de agregados (Figura 2). La observación directa por disección del tracto digestivo evidenció su aporte en la formación de macroagregados del suelo (Figura 3).

DISCUSIÓN
Los procesos de labranza afectaron las poblaciones de lombrices, alcanzando una situación extrema de ausencia total luego de 10 años. Sin embargo, los periodos de barbecho permitieron que los anélidos colonizaran de nuevo estos sitios; el paso del suelo y de materiales orgánicos por el tracto digestivo de la lombriz, junto con su movimiento horizontal en el perfil, mejoraron la estructura y la fertilidad del suelo (Lavelle et al., 1994; Beare et al., 1995; Brown, 1995).
Con el laboreo del suelo la población de lombrices disminuyó, principalmente por exposición, predación y desecación durante la labranza (Holland, 2004); después de 5 años predominó A. corticis con un 53% de los individuos, seguido de Martiodrilus sp con el 27%, y P. corethrurus con el 20%, los cuales no se hallaron después de 10 años debido a la pérdida de su hábitat por la labranza. Estos anélidos conjugan dos estrategias ecológicas: modifican la circulación de los nutrimentos de la materia orgánica de la superficie y procesan las partículas minerales en su intestino modificando la estructura del suelo por su actividad cavadora y la producción de heces al interior y exterior del suelo.
La perturbación debida a 5 años de laboreo del suelo disminuyó la población de lombrices nativas del género Martiodrilus sp., y permitió que las exóticas colonizaran (Lavelle y Pashanasi, 1989; Pizl, 1992; Fragoso et al., 1995; González et al., 1996). Esta situación es similar a la observada por Jiménez et al. (1998) bajo condiciones de sabana en los Llanos Orientales colombianos; en donde, al establecer pasturas mejoradas Martiodrilus, que es endémica, coexiste con especies exóticas en agroecosistemas alterados, pero su abundancia puede ser parcialmente controlada por la masa vegetal que cae al suelo. Feijoo et al. (2004) mencionan que estas lombrices nativas presentan una amplia distribución, viven en ambientes que van desde selvas hasta sistemas de cultivo multiestratificados y pastizales. En el presente estudio solo se encontró en los suelos de cultivo. A. corticis, probablemente originaria de Asia, se encuentra muy bien distribuida en América en un gradiente altitudinal amplio, en ambientes antropocoros, en reservas con sucesión secundaria y también en áreas bien preservadas de Brasil. Al ser una especie migratoria e invasiva, con gran capacidad de colonizar áreas nuevas, se vuelve la especie dominante, pues las nativas y otros organismos edáficos no soportan los cambios; además, ésta se reproduce por partenogénesis, proceso que le favorece en la invasión (Brown et al., 2006; Azevedo, 2010), pueden ser estas las características que se reflejan a través de la explosión poblacional oligoqueta en el suelo con 5 años de barbecho.
Con 5 años en barbecho las poblaciones de lombrices se recuperaron en el suelo; P. corethrurus y A. corticis representaron el 20 y 80% de la población total, respectivamente; después de 10 años de barbecho la cantidad de especímenes bajó casi al 50%, incrementado la proporción de P. corethrurus; ambas especies son reconocidas por su gran tolerancia a diversos factores ambientales (Fragoso et al., 1999) y con potencial para el manejo y rehabilitación de suelos tropicales con exploración minera (García y Fragoso, 2003), gracias a el buen comportamiento y eficiente reproducción en sustratos de baja calidad y lenta descomposición, incluyendo aserrín (García y Fragoso, 2002).
A. corticis comúnmente habita en suelos con abundante cobertura vegetal superficial (Feijoo et al., 2004) por eso se estableció en los suelos con barbecho; mientras que P. corethrurus, originaria del noreste de Suramérica, mora en ambientes antropocoros, en suelos cultivados y pastizales (Azevedo, 2010; Feijoo et al., 2004). Este espécimen halló un favorable entorno y sustrato en los suelos con barbecho, gracias a que presenta un sistema digestivo de alta eficiencia y con excepcionales características demográficas, con lo cual le permitieron colonizar rápidamente el suelo en 5 años de barbecho, lugares perturbados de donde las lombrices de tierra nativas fueron desplazadas (Fragoso y Lavelle, 1992, Edwards y Bohlen 1996, Lavelle et al., 1998; Pashanasi, 2002; Tapia-Coral, 2004). La explosión demográfica fue similar a la encontrada en agroecosistemas café-cítricos (Feijoo et al., 2007) pero allí predominó P. corethrurus con valores que superaron los 750 individuos m-2.
Lee (1985), afirma que la composición de las especies de lombrices en una comunidad depende de la naturaleza de su hábitat y de la capacidad de otras especies para colonizarlo; en Nueva Zelanda el reemplazo de la vegetación natural por pastizales causó prácticamente la pérdida de las especies nativas de lombrices, situación similar a la de este trabajo donde las especies nativas del género Martiodrilus desparecieron con 10 años de laboreo. La colonización posterior en la pastura neozelandesa, fue por especies exóticas, de igual manera en este estudio, en los suelos cultivados y alterados aparecen las especies exóticas en mayor densidad.
Feijoo et al. (2004) mencionan que de las especies exóticas con origen en el neotrópico, como P. corethrurus, son capaces de invadir sistemas de cultivo altamente degradados, pero en este estudio curiosamente apareció en bosque no intervenido, en suelos poco degradados y en recuperación, mientras que no se presentó en suelos con 10 años de labranza, calificados como degradados en sus características físicas. Estos investigadores también asocian la presencia de A. corticis y de P. corethrurus a suelos con 5-10% de materia orgánica, aspecto que no se observa en este caso, pues se presentan en suelos mayores a 17% de materia orgánica. La presencia de estas lombrices en zonas no intervenidas o en recuperación se puede interpretar como un desplazamiento de individuos desde suelos con pobres condiciones para establecerse en las zonas aledañas prístinas o en barbecho y se mantiene por su rápida reproducción (Römbke y Verhaagh 1992; Lavelle et al., 1994).
La mayor cantidad de individuos de Martiodrilus sp y A. corticis se hallaron en el suelo labrado, en los primeros 10 cm de profundidad, concordando con la alta presencia de residuos vegetales que hacen parte de su dieta alimenticia. Sánchez et al. (2003), en estudios realizados en Costa Rica, observaron que las lombrices de tierra mostraron una clara concentración en los 10 cm superiores del suelo, tanto en los pastos como en los bosques jóvenes secundarios, condiciones semejantes a las evaluadas en este trabajo. Según Bouché (1984), Martiodrilus sp. muestra una fuerte evidencia de migración cuando las condiciones son adversas, proceso que pudo ocurrir en suelos con 10 años de labranza. La distribución, el número de individuos, las especies y su categoría ecológica cambiaron con el manejo del suelo y el patrón de comportamiento de las lombrices, que coincidió con el señalado por Fragoso et al. (1999) para otras comunidades, además, afirman que el cambio de la selva natural y las sabanas a agroecosistemas modifican la abundancia, biomasa, número de especies, categorías ecológicas y composición de las especies de lombrices en el suelo.
Las lombrices de tierra presentan una clara importancia como indicadoras de la calidad y salud del suelo están relacionadas; en primer lugar, con el funcionamiento del sistema y los servicios ambientales, tales como fertilidad del suelo y biodiversidad; su hábito alimentario aporta información respecto a la materia orgánica disponible y su categoría ecológica permite inferir sobre la calidad física del perfil del suelo. En segundo lugar, las lombrices juegan un papel importante como bioindicadores o señales del estado de degradación o de recuperación de los agroecosistemas; en este sentido se puede aportar con este trabajo que: la pérdida de la calidad o capacidad productiva se revela con la ausencia de oligoquetos, pero en la medida que el proceso de recuperación se desencadena la población aumenta, hasta cuando alcanza un estado tal de restablecimiento en el que su aporte es mínimo, en consecuencia el número de individuos disminuye.
Las lombrices participan en el proceso de agregación tanto de manera directa al ingerir suelo, como es el caso de las endogeicas que se alimentan principalmente de suelo para generar luego deyecciones; en condiciones prístinas este proceso fue evidenciado gracias a la presencia de la lombriz endogeica de P. corethrurus, que con su hábito alimenticio y proceso digestivo juega un papel crucial en la estructuración del suelo (Figura 3) (Lavelle y Spain, 2001; Domínguez et al., 2004). Este proceso es similar a lo reportado en la sabana de Carimagua en los Llanos Orientales colombianos, en donde se estimó que la población total de lombrices consume entre 30-100 ton suelo ha-1año-1, un 35% del suelo ingerido es depositado en turrículos superficiales y al interior del perfil del suelo; estas deyecciones que logran tamaños superiores a los 5 mm, como en el caso de Martiodrilus carimaguensis, alcanzan una densidad en fresco de 1,3-1,4 g cm-3 y diámetro medio ponderado de 13 mm; sin embargo, después de un año disminuye la densidad a 1,1 g cm-3 y el diámetro a 5,6 mm, en donde es posible que otros invertebrados los disgregan en menores tamaños (Decaens et al., 2001).
Las especies epigeas presentan un alto consumo, digestión y asimilación de la materia orgánica, produciendo deyecciones holorgánicas que juegan un papel clave en la transformación del mantillo (Liu y Zou, 2002), en donde se genera un efecto indirecto en la agregación debido al ataque microbiano de las heces (Domínguez et al., 2009; Lores et al., 2006), que sería el caso de A. corticis en el suelo con barbecho. Las lombrices además de la producción de moco, excretan sustancias como la urea y el amonio que constituyen una fuente de nutrientes fácilmente asimilables para los microorganismos (Dominguez et al., 2009), todas estas sustancias orgánicas son agentes agregantes que propician estructuración del suelo.
CONCLUSIONES
La labranza afecta el número de individuos, la densidad y la distribución de lombrices en el suelo, presentando resistencia a disminuir la población durante los primeros 5 años; pero al prolongarse esta condición aun las poblaciones más fuertes desaparecen. Por su parte, el barbecho del suelo durante los primeros 5 años genera condiciones para que las lombrices colonicen y recuperen su población, la cual supera significativamente lo cuantificado en suelo virgen; no obstante, luego de 10 años la densidad se aproxima a la de suelo no intervenido. La alta población de lombrices puede indicar que se está desarrollando el proceso de recuperación del suelo, posteriormente su descenso puede ser señal de rehabilitación. Es así como un dato puntual de población de lombrices no indica el estado de degradación o recuperación del suelo.
AGRADECIMIENTOS
Al doctor Mario Hernán Londoño Mesa, Profesor Asistente, Instituto de Biología, Universidad de Antioquia Medellín, Colombia, por su apoyo en la identificación de los especimenes.
BIBLIOGRAFÍA
Azevedo, P.T. 2010. Minhocas, fungos micorrizicos arbusculares e bacterias diazotroficas en mudas de Araucaria angustifolia. Tesis de Maestría, Escuela Superior de Agricultura Luiz de Queiroz, San Pablo Brasil. 75 p. [ Links ]
Beare, M.H., D.C. Coleman, D.A. Crossley, P.F. Hendrix and P.E. Odum. 1995. A hierarchical approach to evaluating the significance of soil biodiversity to biogeochemical cycling. Plant and Soil 170:5-22. [ Links ]
Benckiser, G. 1997. Organic inputs and soil metabolism. pp. 6-62. In : Benckiser, G. (ed). Fauna in soil ecosystems: recycling processes, nutrient fluxes, and agricultural production. Marcel Deker Inc., New York. 400 p. [ Links ]
Bouché, M. 1984. Los gusanos de tierra. Mundo Científico (Montpellier) 4(40): 954-963. [ Links ]
Brown, G.G. 1995. How do earthworms affect microfloral and faunal community diversity? Plant and Soil 170:209-231. [ Links ]
Brown, G.G., S.W. James, A. Pasini, D.H. Nunes, N.P. Benito, P.T. Martins and K.D. Sautter. 2006. Exotic, peregrine, and invasive earthworms in Brazil: diversity, distribution, and effects on soils and plants. Caribbean Journal of Science 42(3): 339-358. [ Links ]
Burbano, O. 1998. La materia orgánica: origen, propiedades y su relación con la calidad/salud del suelo. En: Memorias IX Congreso Colombiano de la Ciencia del Suelo. Sociedad Colombiana de la Ciencia del Suelo. Paipa. [ Links ]
Coral, D.M. y C.R. Bonilla. 1998. Impacto de las prácticas agrícolas sobre la macrofauna del suelo en la cuenca alta del lago Guamues, Pasto, Colombia. Suelos Ecuatoriales 32(1): 157-160. [ Links ]
CORNARE, 1995. Corporación Autónoma Regional Rio Negro y Nare, Antioquia. Uso potencial del suelo en los municipios del altiplano del oriente antioqueño colombiano. 390 p. [ Links ]
Darrigran, G., A. Vilches, T. Legarralde y C. Damborenea. 2007. Guía para el estudio de macroinvertebrados. Métodos de colecta y técnicas de fijación. ProBiota, FCNyM. Serie Técnica Didáctica No. 10: 45-46. [ Links ]
Decaens, T., J.J. Jiménez, A.F. Rangel, A. Cepeda, A.G. Moreno y P. Lavelle. 2001. Capítulo 7: La macrofauna del suelo en la sabana bien drenada de los Llanos Orientales. pp. 111-137. En: Rippstein, G., G. Escobar and F. Motta (eds.). Agroecología y Biodiversidad de las Sabanas en los Llanos Orientales de Colombia. CIAT-CIRAD. Colombia. 302 p. [ Links ]
De Leehneer and M. De Boodt. 1958 Soil structure index and plant grow. pp. 234-244. In: International Symposium on Soil Structure. Ghent, Belgian. [ Links ]
Domínguez, J., P.J. Bohlen and R.W. Parmelee. 2004. Earthworms increase nitrogen leaching to greater soil depths in row crop agroecosystems. Ecosystems 7:672-685. [ Links ]
Domínguez, J.M., L. Aira y M. Gómez-Brandón. 2009. El papel de las lombrices de tierra en la descomposición de la materia orgánica y el ciclo de nutrientes. Ecosistemas 18(2): 20-31. [ Links ]
Edwards, C.A. and P.J. Bohlen. 1996. Biology and Ecology of Earthworms. Tercera Edición. Chapman and Hall. London. 426 p. [ Links ]
Feijoo, A., H. Quintero, C. Fragoso y A. Moreno. 2004. Patrón de distribución y listado de especies de las lombrices de tierra (Annelida: Oligochaeta) en Colombia. Acta Zoológica Mexicana 20(2): 197-220. [ Links ]
Feijoo, A., M. Zuniga, H. Quintero y P. Lavelle. 2007. Relaciones entre el uso de la tierra y las comunidades de lombrices en la cuenca del río La Vieja, Colombia. Pastos y Forrajes 30(2): 1-1. [ Links ]
Fragoso, C., J. Kanyonyo, A. Moreno, B.K. Senapati, E. Blanchart and C. Rodríguez. 1999. A survey of tropical earthworms: taxonomy, biogeography and environmental plasticity. pp. 1-26. In: Lavelle, P., L. Brussaard and P.F. Hendrix. (eds.), Earthworm management in tropical agroecosystems. CABI International, Wallingford. 300 p. [ Links ]
Fragoso, C., S.W. James and S. Borges. 1995. Native earthworms of the north neotropical region: current status and controversies. pp. 67-99. In: Hendrix, P.F. (ed.). Earthworm Ecology and Biogeography in North America. Lewis Publishers, Boca Raton, Florida. 256 p. [ Links ]
Fragoso, C. and P. Lavelle. 1992. Earthworm communities of tropical rain forests. Soil Biology and Biochemistry 24(12): 1397-1408. [ Links ]
García, J.A. and C. Fragoso. 2002. Growth, reproduction and activity of earthworms in degraded and amended tropical open minded soils: laboratory assays. Applied Soil Ecology 20, 43-56. [ Links ]
García, J.A. and C. Fragoso. 2003. Influence of different food substrates on growth and reproduction of two tropical earthworm species (Pontoscolex corethrurus and Amynthas corticis). The 7th International Symposium on Earthworm Ecology - Cardiff - Wales - 2002. Pedobiologia 47(5-6): 754-763. [ Links ]
González, G., X. Zou and S. Borges. 1996. Earthworm abundance and species composition in abandoned tropical croplands: comparisons of tree plantations and secondary forests. Pedobiologia 40: 385-391. [ Links ]
Holland, J.M. 2004. The environmental consequences of adopting conservation tillage in Europe: reviewing the evidence. Agriculture Ecosystems Environment 103(1): 1-25. [ Links ]
Jimenez, J.J., A.G. Moreno, T. Decaëns, P. Lavelle, M.J. Fisher and R.J. Thomas. 1998. Earthworm communities in native savannas and man-made pastures of the Eastern Plains of Colombia. Biology and Fertility Soils 28:101-110. [ Links ]
Jones, D.T., W.R. Taylor and J.M. Thornton. 1994. A model recognition approach to the prediction of all-helical membrane protein structure and topology. Biochemistry 33(10): 3038-3049. [ Links ]
Jongmans, A., M. Pulleman and Marinissen. 2001. Soil structure and earthworm activity in a marine silt loam under pasture versus arable land. Biology and Fertility of Soil 33: 279-285. [ Links ]
Lavelle, P., M. Dangerfield, C. Fragoso, V. Eschenbrenner, D. López, B. Pashanasi and L. Brussaard. 1994. The relationship between soil macrofauna and tropical fertility. pp. 137-169. In: Woolmer, P.L. and M.J. Swift (eds.). The biological management of tropical soil fertility. John Wiley and Sons, Chichester. 243 p. [ Links ]
Lavelle, P. 1997. Faunal activities and soil processes: adaptive strategies that determine ecosystem function. Advances in Ecological Research 27: 93-132. [ Links ]
Lavelle, P., B. Pashanasi, F. Charpentier, C. Gilot, J.P. Rossi, L. Derouard, J. Andre, J.F. Ponge and N. Bernier. 1998. Large-scale effects of earthworms on soil organic matter and nutrient dynamics. pp. 103. In: Edwards, C.A. (ed.). Earthworm Ecology. Saint Lucie Press, London. 389 p. [ Links ]
Lavelle, P. and A.V. Spain. 2001. Soil ecology. Kluwer Academic Publishers, London. 684 p. [ Links ]
Liu, Z.G. and X.M. Zou. 2002. Exotic earthworms accelerate plant litter decomposition in a Puerto Rico pasture and a wet forest. Ecological Applications 12(5): 1406-1417. [ Links ]
Lores, M., M. Gómez-Brandón, D. Pérez-Díaz y J. Domínguez. 2006. Using FAME profiles for the characterization of animal wastes and vermicomposts. Soil Biology and Biochemistry 38(9): 2993-2996. [ Links ]
Lee, K. 1985. Earthworms: their ecology and relationships with soils and land use. Academic Pres, New York. 411 p. [ Links ]
Pashanasi, B. 2002. Estudio cuantitativo de la macrofauna del suelo en diferentes sistemas de uso de la tierra en la Amazonía Peruana. Folia Amazónica 12(1-2): 75-97. [ Links ]
Pizl, V. 1992. Succession of earthworm populations in abandoned fields. Soil Biology and Biochemistry 24(12): 1623-1628. [ Links ]
Ramírez, R. 2008. Cambios en los macroagregados por prácticas agrícolas y procesos de recuperación, en un Andisol, en la Vereda Montañitas, Municipio de Marinilla, oriente antioqueno colombiano. Tesis Doctoral. Universidad Nacional de Colombia - Sede Palmira. 204 p. [ Links ]
Römbke, J., and M. Verhaagh. 1992. About earthworm communities in a rain forest and an adjacent pasture in Peru. Amazoniana 12(1): 29-49. [ Links ]
Sanchez - de Leon, Y., Z. Xiaoming, S. Borges and R. Honghua. 2003. Recovery of native earthworms in abandoned tropical pastures. Conservation Biology 17(4): 999-1006. [ Links ]
Six, J., E.T. Elliot and K. Paustian. 2000. Soil macroaggregate turnover and microaggregate formation: a mechanism for C sequestration under no-tillage agriculture. Soil Biology and Biochemistry 32(14): 2099-2103. [ Links ]
Tapia-Coral, S.C. 2004. Macro-invertebrados do solo e estoques de carbono e nutrientes em diferentes tipos de vegetação de terra firme na Amazônia peruana. Tese de Doutorado en Ciencias Biológicas. Instituto Nacional de Pesquisas da Amazônia (INPA). Universidade Federal do Amazonas (UFAM). Brasil. 135 p. [ Links ]
