










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
versión impresa ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.70 no.1 Medellín ene./abr. 2017
https://doi.org/10.15446/rfna.v70n1.61765
DOI: http://dx.doi.org/10.15446/rfna.v70n1.61765
Effect of post-harvest forestry residue management practices on the diversity of epigeal coleopterans
Efecto de prácticas de manejo de residuos post cosecha sobre la diversidad de coleópteros epigeos
Priscila Ramírez Aliaga1, Amanda Huerta Fuentes1 and Jaime E. Araya2
1 Facultad de Ciencias Forestales y Conservación de la Naturaleza. Universidad de Chile. Casilla 9206, Santiago, Chile. <ahuertaf@gmail.com>
2 Facultad de Ciencias Agronómicas. Universidad de Chile. Casilla 1004, Santiago, Chile.
Received: April 4, 2016; Accepted: October 27, 2016
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License.

ABSTRACT
The aim of this work was to study the effect of post-harvest forestry residue management practices on the epigeal coleopterans diversity and abundance in a 1-yr old Pinus radiata plantation. Approximately 25-yr old P. radiata plantation was available, which was harvested by clear-cutting at the beginning of 2010 in Constitución, central Chile. Three post harvest residue management treatments were installed in a fully randomized design in blocks: 1) a control, where residues were intact left; 2) residue removed (> 2.5 cm diameter); and 3) burned. The coleopterans were sampled in spring and summer, using linear transects with pitfall traps. The abundance and richness, and diversity index of coleopterans were determined. A total of 23 species of 13 families were obtained. Both in the spring and summer, the treatment with residues left registered significantly greater abundance and species richness. In the residue management, the practices of intact leaving residue should be considered as the least impacting on diversity and abundance of epigeal coleopterans and is recommended for creating refugee areas to promote diversity of beetles in this area of study.
Key words: Insects, Pinus radiata, Diversity, Richness, Abundance.
RESUMEN
El objetivo de este trabajo fue estudiar el efecto de prácticas de manejo de residuos de post-cosecha en plantaciones de un año de Pinus radiata sobre la diversidad y abundancia de coleópteros epigeos. El estudio se realizó en Constitución, Chile central, donde existía una plantación de P. radiata de cerca de 25 años, que se cosechó con el método de tala rasa a comienzos de 2010. Se consideraron tres tratamientos de post-cosecha con un diseño completamente aleatorio en bloques: 1) uno control, con los residuos intactos; 2) con extracción de residuos (> 2,5 cm de diámetro); y 3) con quema. Los coleópteros se muestrearon en primavera y verano mediante transectos lineales con trampas de caída. Se determinó la abundancia y riqueza, y un índice de diversidad de coleópteros epigeos. Se obtuvieron un total de 23 especies de 13 familias. En primavera y verano, el tratamiento con residuos dejados intactos tuvo una abundancia y riqueza de especies significativamente mayor. Cuando se apliquen tratamientos de post-cosecha la práctica de dejar los residuos intactos debería ser considerada como la de menos impacto sobre la diversidad y abundancia de coleópteros epigeos y ser recomendada por crear áreas de refugio para promover la diversidad de coleópteros en esta área de estudio.
Palabras claves: Insectos,Pinus radiata, Diversidad, Riqueza, Abundancia.
Insects play important role in ecosystem functioning, and act as predators, pollinators, or decomposers of organic matter, and as indicators of biodiversity capable of reflecting the level of conservation, diversity, endemism, and degree of intervention or disturbance of the ecosystem (Coddington et al., 1991; Colwell and Coddington, 1994). Also most insects possess the ability of detecting changes in the functioning of forest ecosystems (Didham, 1997; Langor and Spence, 2006), and are very vulnerable to fragmentation and habitat loss. Similarly, habitat loss has strong effects on biodiversity, and is considered the most important factor in the extinction of populations and species (Fahrig, 2003). The negative effects of habitat loss refer not only to attributes of biodiversity, such as species richness (Steffan-Dewenter et al., 2002; Steffan-Dewenter, 2003), the abundance and distribution of populations (Best et al., 2001), and genetic diversity, but also to patterns or ecological processes that depend of biodiversity (Keller et al., 2005). The loss of biodiversity due to human activities has become one of the major environmental problems of global concern. Anthropogenic activities like habitat overuse, deforestation, pollution, and introduction of exotic species, among others, are the main forces of changes in community diversity and composition throughout the world (Spooner, 2005; Coates et al., 2006; Roe et al., 2006). Thus, the replacement of natural vegetation by forest plantations as those Pinus radiata D. Don, can be considered a building process of "green deserts" in which biodiversity is scarce when compared with those harboring native forests of the same geographic areas (Bonham et al., 2002).
Human endeavors, such as harvesting, modify forest composition and dynamics, affecting biodiversity. Different forest management systems influence the edaphic microarthropod fauna, which in turn is important to maintain the properties of the soil subsystem (Covarrubias and Contreras, 2004). Forest practices may have profound effects on population levels and species composition of diverse organism groups (Jactél et al., 2005). Also, managing vegetation allows to control forest pests and to keep groups of undesired insects below damaging levels (Smith, 1990). Currently, in Chile, there are about 1,600,000 ha to P. radiata (INFOR, 2011). This large area represents a high risk for the propagation of pests and diseases, even though at world level, P. radiata is one of the forest species most resistant to sanitary problems (Clapp, 2001). Despite the extensive territory covered by forest plantations in the country, the effect of forest residue managements has been documented scarcely (Briones and Jerez, 2007). Most insects have the ability to detect changes in the functioning of forest ecosystems. This is evident when observing alterations in the distribution, abundance and composition in the communities of these organisms (Langor and Spence, 2006). Among the organisms most potentially affected by plantations of exotic species are epigeal (i.e. ground-dwelling) insects such as beetles and ants, given their high sensitivity to changes in ecosystems (Sackman et al., 2008). The epigeal coleopteran constitutes a taxonomically and ecologically diverse group, and hold documented potential as model organisms or indicators for ecological impact research (Paritsis and Aizen, 2008; Koivula, 2011; Roberge and Stenbacka, 2014). The hypothesis of this investigation was that the post-harvest residue managements affect the diversity of epigeal coleopterans by season. In this study we aim to examine if the post harvest residue management of P. radiata plantations affect the diversity of epigeal coleopterans in the Maule region of Chile.
MATERIALS AND METHODS
The survey area is located in an approximately 25-yr old plantation of P. radiata (3 ha) (35°43' - 35°47'S; 72°29' - 72°31'W), which was harvested by clear-cutting at the beginning of 2010 in Constitución, Maule Region, central Chile. This area belongs to the Experiment Center Dr. Justo Pastor León of the Faculty of Forestry Sciences and Nature Conservation, University of Chile. The area is located in the dry coastal Talca Province in the district of Agroclimatic Empedrado-Coronel, which is characterized by an average annual rainfall of 897 mm, with minimum and maximum temperature of 6 °C and 24.4 °C, respectively. The soils belong to the Constitución series, originating from metamorphic-granitic (Peralta, 1976). The study area is between coastal Nothofagus glauca and Azara petiolaris, and N. glauca and Persea lingue Mediterranean deciduous forests, being the first forest dominated by N. glauca, Gevuina avellana, Lomatia hirsuta, and A. petiolaris as major tree species, and the second forest dominated by N. glauca, Nothofagus obliqua, G. avellana, and P. lingue (Luebert and Pliscoff, 2006). In the study area original sclerophyllous forests and shrubs have been subjected to a profound transformation so that in some deeply disturbed places the original vegetation has been completely transformed into ruderal shrubs associated with the introduction of allochthonous species Pinus radiata plantations (Amigo et al., 2000; Luebert and Pliscoff, 2006).
In three different sites three post-harvest treatments (plots of 30m x 30m) were applied in a fully randomized design: i) a control in which harvest residues were left intact, ii) a in which harvest residues greater than 2.5 cm in diameter were removed, and iii) a in which all the harvest residues were burned (letting the fire to grow against the breeze or down the slope) (Julio, 2005). These three sites are separated around 20 m between them. Three linear transect were randomly selected in each one the three post harvest residue management treatments (experimental unit). In each linear transect of 20 m was delimited placing three pitfall traps within it at 0, 10 and 20 m of distance. Thus, a total of 27 pitfall traps (3 traps x 3 transects x 3 treatments) were finally disposed to examine the variation in epigeal coleopterans. The pitfall traps have a capacity of 750 mL being filled with 250 mL of 75% ethanol to preserve the specimens and 500 mL of distiller water with traces of detergent to decrease surface tension. The beetles were sampled in October 2010 (spring) and January 2011 (summer) due to temporal variations. It was considered that each pitfall trap had an activity radius of 5 m (Péfuar and Pérez, 1995), and then each transect was transformed to an area of 0.0236 ha. All the pitfall traps were removed 15 days after installation (Briones and Jerez, 2007).
The insects were stored in plastic vials with 70% ethanol and later counted and identified using taxonomic keys and by comparison with specimens in entomological collections in the Forest Entomology Laboratory, Faculty of Forestry Sciences and Nature Conservation, University of Chile, and the Entomology Institute, Metropolitan University of Education Sciences, in Santiago, Chile (Escobar, 2000).
The alpha diversity index of the epigeal coleopterans was determined for each treatment and season by calculation of means of relative abundance and specific richness (Moreno, 2001a). To determine the diversity of taxonomic groups the Shannon-Wiener (H') index was calculated (Magurran, 1988). In addition, a factorial ANOVA was used to determine statistical differences in species abundance and richness between residue treatments and season as factors. Finally, the HSD Tukey tests (P<0.05) a posteriori was executed.
RESULTS AND DISCUSSION
With 27 traps we obtained 176 specimens that represent 23 species of 13 families of Coleoptera. Five most abundant species were Enneboeus sp. (Archaeocrypticidae), Homalotrichus striatus, Cyanotarsus foveolatus (Staphylinidae), Lathrobium dimidiatum (Staphylinidae) and Hylurgus ligniperda (Curculionidae). Some species are represented in nearly all treatments, as Hylurgus liniperda and Homalotrichus striatus. Few species are present only in one treatment and scarce abundance, as Acanthinodera cumingii (Cerambycidae), Germarostes posticus (Hybosoridae), Deromecus vulgaris (Elateridae) and Afrasida propensa (Tenebrionidae) (Table 1).
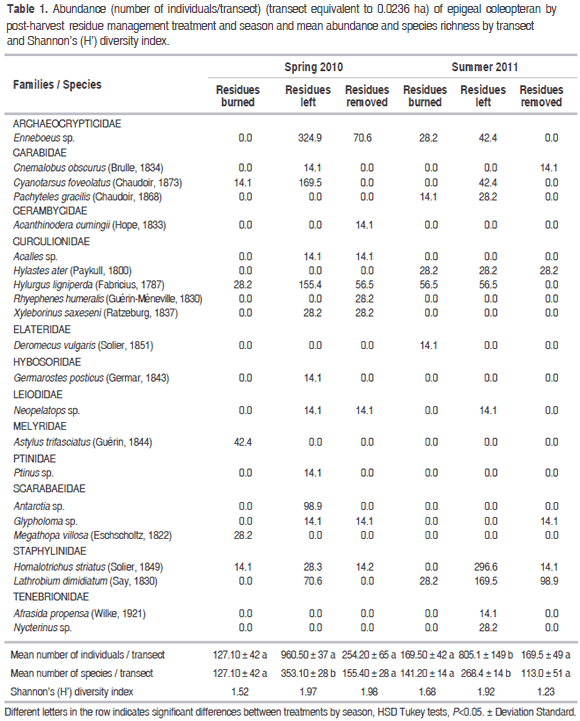
The treatment is the only factor affecting abundance variations (F2,12=61.02, P<0.001) while season factor and the "treatment by season" interaction term are not statistically significant (F1,12=1.06, P=0.32 and F2,12=0.82, P=0.45, respectively), showing that effect of the different treatments seem to not depend of the season. During spring, the treatment with residues left registered significantly greater species abundance than the other two (87% greater than the treatments with residues burned, and 73% more than that with the residues extracted). This trend also occurred during the summer, when the treatment with residues left continued to have a significantly greater abundance (79% greater than the treatments with residues burned or extracted). When comparing the abundance in each treatment between spring and summer no differences occurred between the treatment with residues burned, extracted or left (Table 1).
Similar results can be observed in the case of species richness. The species richness of epigeal coleopterans significantly differ between treatments (F2,12=23.56, P<0.001), but no when the two seasons are compared (F1,12=2,97, P=0.11). During the spring, the treatment with residues left had a significantly greater richness than the other two treatments (64 and 56% greater than in the treatment with residues burned and extracted, respectively). During the summer, the treatment varied slightly, and the treatment with residues left continued having greater species richness, 47% greater than the treatment with residues burned and 58%, a significantly greater level than the treatment with the residues extracted, respectively. Again, the "treatment by season" interaction is not statistically significant (F2,12 =1.74, P=0.21) (Table 1).
Shannon's diversity index (H') did not register significant differences between species of epigeal coleopterans.The greatest abundance of epigeal coleopterans occurred in the treatment with residues left in both seasons, maybe because many species found there better habitat conditions and trophic resources necessary for survival (Grez et al., 2003; Correa and Roa, 2005). Work et al. (2013), evaluate the initial changes in the abundance, species richness and community composition of rove (Staphylinidae) and ground beetles (Carabidae), immediately following 1) stem-only harvesting, in which logging debris (e.i. tree tops and branches) are retained on site, and 2) whole-tree harvesting, in which stems, tops and branches are removed in mature balsam fir stands in Quebec, Canada. Catch rates in whole-tree harvesting were greater than stem-only harvesting sites; however, they were attributable to increased catches of only three species of Staphylinidae.
The greatest richness of epigeal coleopterans occurred in the treatment with residues left in both seasons, which may be due to the greater availability of food, favorable habitat and resource conditions preserved in this treatment, allowing for their establishment and survival, and resulting in an increased attraction for diverse species (Moreno, 2001b; Grez et al., 2003). The treatment with residues burned had the least species richness, probably because burning had a greater effect on natural habitat of insects affecting the ground and vegetation, thus the insects did not find the adequate conditions for establishment. Besides, insects are indirectly affected by fire given soil warming, increased metabolic activity and CO2 emissions from litter and soil (Price et al., 2011).
The variation in abundance and species richness of epigeal coleopterans in both seasons could be caused by some species being univoltine while others may develop several life cycles in a year; an example is Hylurgus ligniperda, one of the most abundant species with not a seasonal cycle, as the females begin galleries any time of the year, although more frequently in the warmer months, with 10-12 wk cycles (Artigas, 1994).
The results obtained with the post-harvest treatments indicate that the effects of management of forest residues in harvest impacts on biodiversity and that different treatments influence biodiversity in diverse ways. The production process should consider preserving fragments of biodiversity reservoirs for preservation (Fisher and Lindenmayer, 2002; Tscharntke et al., 2002). Also, it is known that vegetation with residues possess a specific fauna and provides refugees for many sensitive species, and it is necessary to keep this in mind when anthropogenic changes of landscape occur. The preservation of biodiversity depends progressively on the protection of small areas and biota outside protected wild areas (Simonetti, 1998). This may be part of the strategies to follow to maintain biodiversity, particular in the region studied, that presents a high degree of endemism (Simonetti et al., 2002; Simonetti, 2006).
CONCLUSIONS
Both in the spring and summer, the treatment with residues left registered significantly greater abundance and species richness of epigeal coleopterans. This treatment is then recommended in this area of study as it increases food availability and creates refugee site for beetles because of its greater vegetation cover, an important factor to promote their diversity. Then, when applying post-harvest residue management practices in this area of study is important to consider the treatment with the least alterations of the site, to generate adequate refugee areas to promote epigeal coleopteran diversity.
ACKNOWLEDGMENTS
Project financed by CORFO - INNOVA - BIOCOMSA.
REFERENCES
Amigo J, San Martín J y García L. 2000. Estudio fitosociológico de los bosques de Nothofagus glauca (Phil.) Krasser del Centro-Sur de Chile. Phytocoenologia 30:193-222. [ Links ]
Artigas J. 1994. Entomología económica: Insectos de interés agrícola, forestal, médico y veterinario (nativos, introducidos y susceptibles de ser introducidos). Vol 2. Ediciones Universidad de Concepción, Concepción, Chile. 943 p. [ Links ]
Best L, Bergin T and Freemark K. 2001. Influence of landscape composition on bird use of rowcrop fields. Journal of Wildlife Management 65: 442-449. doi: 10.2307/3803096 [ Links ]
Bonham K, Mesibov R and Bashford R. 2002. Diversity and abundance of some ground-dwelling invertebrates in plantation vs. native forests in Tasmania, Australia. Forest Ecology and Management 158: 237-247. doi: 10.1016/S0378-1127(00)00717-9 [ Links ]
Briones R y Jerez V. 2007. Efecto de la edad de la plantación de Pinus radiata en la abundancia de Ceroglossus chilensis (Coleoptera: Carabidae) en la Región del Bíobío, Chile. Bosque 28: 207-214. [ Links ]
Clapp R. 2001. Tree farming and forest conservation in Chile: Do replacement forests leave any originals behind? Society and Natural Resources: An International Journal 14: 341-356. doi: 10.1080/08941920119176 [ Links ]
Coates F, Lunt I and Tremblay R. 2006. Effects of disturbance on population dynamics of the threatened orchid Prasophyllum correctum D.L. Jones and implications for grassland management in the south-eastern Australia. Biological Conservation 129: 59-69. doi: 10.1016/j.biocon.2005.06.037 [ Links ]
Coddington J, Griswold C, Silva D, Peñaranda E and Scott S. 1991. Designing and testing samplings protocols to estimate biodiversity in tropical ecosistems. pp. 44-60. In: Duddley E. (ed.). The unity of evolutionary Biology: Proceedings of the Fourth International Congress of Systematic and Evolutionary Biology. Dioscorides Press, Portland OR. 1048 p. doi: 10.2307/2413587 [ Links ]
Colwell R and Coddington J. 1994. Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of The Royal Society B: Biological Sciences 345: 101-118. doi: 10.1098/rstb.1994.0091. [ Links ]
Correa P y Roa A. 2005. Relaciones tróficas entre Oncifelis guigna, Lycalopex culpaeus, Lycalopex griseus y Tyto alba en un ambiente fragmentado de la zona central de Chile. Mastozoología Neotropical 12: 57-60. [ Links ]
Covarrubias R. y Contreras A. 2004. Variación en los microartrópodos del suelo, por manejos forestales de raleo selectivo y cortes en hoyos de luz. Bosque 25: 103-116. [ Links ]
Didham R. 1997. An overview of invertebrate responses to forest fragmentation. pp. 303-320 In: Watt AD, Stork NE and Hunter MD (eds.). Forest and insects. Chapman and Hall, London. [ Links ]
Escobar F. 2000. Diversidad de coleópteros coprófagos (Scarabaeidae: Scarabaeinae) en un mosaico de hábitats en la Reserva Natural Nukak, Guaviare, Colombia. Acta Zoológica Mexicana 79: 103-121 [ Links ]
Fahrig L. 2003. Effects of habitat fragmentation on biodiversity. Annual Review of Ecology, Evolution and Systematics 34: 487-515. doi: 10.1146/annurev.ecolsys.34.011802.132419 [ Links ]
Fischer J and Lindenmayer D. 2002. Small patches can be valuable for biodiversity conservation: two case studies on birds in southern Australia. Biological Conservation 106: 129-136. doi: 10.1016/S0006-3207(01)00241-5 [ Links ]
Grez A, Moreno P y Elgueta M. 2003. Coleópteros (Insecta: Coleoptera) epigeos asociados al bosque maulino y plantaciones de pino aledañas. Revista Chilena de Entomología 29: 9-18. [ Links ]
INFOR (Instituto Forestal). 2011. Anuario Forestal 2011. Boletín Estadístico No. 132. INFOR, Santiago, Chile. 136 p. [ Links ]
Jactél H, Brockerhoff E and Duelli P. 2005. A test of the biodiversity - stability theory: meta-analysis of tree species diversity effects on the insect pest infestations, and re-examination of responsible factors. pp. 235-262. In: M. Scherer-Lorenzen , C. Körner, E.D. Schulze. Forest diversity and function: temperate and boreal systems Ecological studies 176. DEU: Springer-Verlag. doi: 10.1007/3-540-26599-6_12 [ Links ]
Julio G. 2005. Fundamentos del manejo del fuego. Quinta edición. Facultad de Ciencias Forestales, Universidad de Chile, Santiago, Chile. 329 p. [ Links ]
Keller I, Excoffier L and Largiadèr C. 2005. Estimation of effective population size and detection of a recent population decline coinciding with habitat fragmentation in a ground beetle. Journal of Evolutionary Biology 18: 90-100. [ Links ]
Koivula M. 2011. Useful model organisms, indicators, or both? Ground beetles (Coleoptera, Carabidae) reflecting environmental conditions. ZooKeys 100: 287-317. doi: 10.3897/zookeys.100.1533 [ Links ]
Langor D and Spence J. 2006. Arthropods as ecological indicators of sustainability in Canadian forests. The Forestry Chronicle 82: 344-350. [ Links ]
Luebert F y Pliscoff P. 2006. Sinopsis bioclimática y vegetacional de Chile. Editorial Universitaria, Santiago, Chile. 316 p. [ Links ]
Magurran A. 1988. Ecological diversity and its measurement. Princeton University Press, New Jersey. 256 p. [ Links ]
Moreno C. 2001a. Métodos para medir la biodiversidad. Vol. I, Zaragoza, España, M&T-Manuales y Tesis SEA. 86 p. [ Links ]
Moreno P. 2001b. Efecto de la fragmentación del bosque maulino sobre la composición, abundancia y distribución de tamaños corporales de coleópteros epigeos. Memoria Médico Veterinario. Santiago, Universidad de Chile, Facultad de Ciencias Veterinarias y Pecuarias. 71 p. [ Links ]
Péfuar J y Pérez R. 1995. Zoogeografía y variación espacial y temporal de algunos vertebrados epígeos de la zona xerófila de la cuenca media del río Chama, Mérida, Venezuela. Ecotrópicos 8(1-2):15-38. [ Links ]
Peralta M. 1976. Uso, clasificación y conservación de suelos. Ministerio de Agricultura, Servicio Agrícola y Ganadero, Santiago, Chile. 340 p. [ Links ]
Paritsis J, Aizen MA. 2008. Effects of exotic conifer plantations on the biodiversity of understory plants, epigeal beetles and birds in Nothofagus dombeyi forests. Forest Ecology Managemet 255: 1575-1583. doi.org/10.1016/j.foreco.2007.11.015 [ Links ]
Price P, Denno R, Eubanks M, Finke D and Kaplan I. 2011. Insect ecology. Behavior, population, and communities. Cambridge University Press, New York. 801 p. [ Links ]
Roe J, Gibson J and Kingsbury B. 2006. Beyond the wetland border: Estimating the impact of roads for two species of water snakes. Biological Conservation 130: 161-168. doi: 10.1016/j.biocon.2005.12.010 [ Links ]
Roberge JM, Stenbacka F. 2014. Assemblages of epigaeic beetles and understory vegetation differ between stand of an introduced pine and its native congener in boreal forest. Forest Ecology and Management 318: 239-249. doi.org/10.1016/j.foreco.2014.01.026 [ Links ]
Sackmann P, Corley J, Villalobos J. 2008. La importancia del manejo de las plantaciones de pinos en la conservación de la diversidad de insectos epígeos. pp. 1-15. In: Villacide J, Coreley J. (Eds). Manejo integrado de Plagas. Laboratorio de Ecología de Insectos. Ediciones Instituto Nacional de Tecnología Agropecuario, Bariloche, Argentina. [ Links ]
Simonetti J. 1998. Áreas silvestres protegidas: ¿protegidas y protectoras? pp. 123-131. In: Díaz-Pineda F, De Miguel J y Casado M (eds.). Diversidad biológica y cultura rural en la gestión ambiental del desarrollo. Ediciones Mundi-Prensa, Madrid. [ Links ]
Simonetti J. 2006. Conservación de biodiversidad en ambientes fragmentados: El caso del bosque Maulino. pp. 213-228. En: Grez A, Simonetti J y Bustamante R (eds.). Biodiversidad en ambientes fragmentados de Chile: patrones y procesos a diferentes escalas. Editorial Universitaria, Santiago, Chile. [ Links ]
Simonetti J, Grez A y Bustamante R. 2002. El valor de la matriz en la conservación ambiental. Revista Ambiente y Desarrollo 18: 116-118. [ Links ]
Smith S. 1990. The greening of the forest: forest pest management into the 21st century. Proceedings of the Entomological Society of Ontario 121: 49-60. [ Links ]
Spooner P. 2005. Response of Acacia species to disturbance by roadworks in roadside environments in southern New South Wales, Australia. Biological Conservation 122: 231-242. doi: 10.1016/j.biocon.2004.07.012 [ Links ]
Steffan-Dewenter I. 2003. Importance of habitat area and landscape context for species richness of bees and wasps in fragmented orchard meadows. Conservation Biology 17: 1036-1044. doi: 10.1046/j.1523-1739.2003.01575.x [ Links ]
Steffan-Dewenter I, Münzenberg U, Bürger C, Thies C and Tscharntke T. 2002. Scale-dependent effects of landscape context on three pollinator guilds. Ecology 83: 1421-1432. doi: 10.1890/0012-9658(2002)083[1421:SDEOLC]2.0.CO;2 [ Links ]
Tscharntke T, Steffan-Dewenter I, Kruess A and Thies C. 2002. Contribution of small habitat fragments to conservation of insect communities of grassland-cropland landscapes. Ecological Applications 12: 354-363. doi: 10.1890/1051-0761(2002)012[0354:COSHFT]2.0.CO;2 [ Links ]
Work T, Klimaszewski J, Thiffault E, Bourdon C, Paré D, Bousquet Y, Venier L, Titus B. 2013. Initial responses of rove and ground beetles (Coleoptera, Staphylinidae, Carabidae) to removal of logging residues following clearcut harvesting in the boreal forest of Quebec, Canada. ZooKeys 258: 31-52. doi.org/10.3897/zookeys.258.4174 [ Links ]

