











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkIn Colombia, Physalis peruviana L., known as golden berry, is a fruit of exportation that has had a good reception in the European international market. The crop has increased from 221 cultivated hectares in 1999 (Sanabria, 2005) to more than 1,000 ha in 2009 (Agronet, 2015), and by 2005 it was considered the second more exported fruit after banana (Fischer et al., 2005). Nowadays, it has been consolidated as the second most exported fruit product of the country, and in 2016 its total exports were 5.2 million t which represent a value of 23,600 million dollars (López, 2018). Most of the production is obtained by small farmers from the Colombian departments of Cundinamarca, Boyacá and Antioquia (Fischer et al., 2014; Zapata et al., 2002).
Physalis peruviana (physalis = bladder) have numerous common names according to the country/or regions such as Cape gooseberry (South Africa), Inca berry, Aztec berry, Golden berry, giant ground cherry, African ground cherry, Peruvian ground cherry, Peruvian cherry, pokpok (Madagascar), rasbhari (India), poha aguaymanto poha aguaymanto (Peru), uvilla (Ecuador), uchuva (Colombia), harankash (Egypt), amur en cage (France), and physalis (United Kingdom) (Afsah, 2015). The mechanical, physicochemical, nutritional, and medicinal properties, so as its functionality, associated with this fruit have been described by Puente et al. (2011). Its genetic diversity and population structure have been studied by molecular markers (Garzón et al., 2015). Physalis peruviana is the object of several research in order to take advantage of its secondary metabolites, which exhibit extensive biological properties that could have a potential pharmacological, medicinal, and insecticidal use (Aguirre-Ráquira et al., 2014; Cirigliano et al., 2008).
However, there have not been developed technological programs or studies for the crop management. Therefore, the losses rise to around 13% (Sanabria, 2005). The studies of insect pests associated with this plant are incipient (Afsah, 2015). In the Noctuidae family, some polyphagous species that affect several organs at different stages of the P. peruviana phenology such as Spodoptera sp., Agrotis sp. and Feltia sp., known as cutworms. The larvae is the only harmful stage which can cut the seedling at ground level and can feed on branch, roots, leaves, and shoots (Benavides and Mora, 2005).
The damage caused by these cutworms is not harmful enough to the P. peruviana crop, as the damage made by the species that pierce the fruit like Heliothis subflexa and Copitarsia decolora, the former is considered the most important phytophagous of P. peruviana (Zapata et al., 2002). The caterpillars pierce the sepals and feed on the fruit in any of its stages, which could reduce the production to 20% according to the climatic condition and the alternative hosts; after the damage, some fruits can fall to the ground or stay in the plant, but in both cases the fruit loses its market value (Benavides and Mora, 2005).
Noctuidae has been the source of many types of researches, with the aim of understanding its behavior, life cycle, and mainly to find ways to control its populations. However, the researches in P. peruviana are incipient. The aim of this work was to determine the Noctuidae species associated with P. peruviana. The fluctuation of the immature stages according to the development of the plant, and spatial and climatic variables were studied in three farms from the municipality of La Union-Antioquia-Colombia, with the aim of building statistical models to describe the growing trend as a tool for pest management.
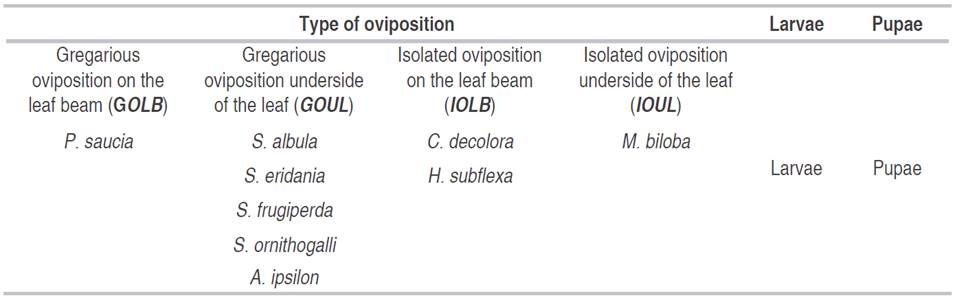
Twelve species of Noctuidae were found; cutworm and leaf eaters included Agrotis ipsilon, Megalographa biloba, Peridroma saucia and four species of Spodoptera (S. albula, S. eridania, S. frugiperda and S. ornithogalli); and two fruit borer, Copitarsia decolora and Heliothis subflexa. Six models clustering ecological and biological similarities of the species were built, having into account climatic, spatial and temporal variables. Four oviposition models were built for the species A. ipsilon and Spodoptera spp.; C. decolora and H. subflexa; M. biloba; and, P. saucia; besides, larvae and pupae models, without regards of the species. All models, except that of M. biloba, are important for predicting purposes in pest management programs.
MATERIALS AND METHODS
Location
The study included 12 sampling units distributed throughout three producer farms of P. peruviana in the municipality of La Unión, Antioquia, Colombia. The geographic coordinates of the first farm are 461186.23 m longitude W and 659246.13 m latitude N, the second farm with 460238.06 m longitude W and 659604.24 m latitude N, and the third farm with 459443.45 m longitude W and, 660349.50 m latitude N, with WGS 84 UTM 18N coordinate system, and 2.509 m mean altitude. The study was performed with plants between 30 and 45 Days After Transplant (DAT) and the sampling were made biweekly, from March 1st to August 29th of 2014. Before the P. peruviana was planted, the farms had different covers, which is a critical factor concerning the presence of phytophagous insects and the diseases. The previous cover for the farm 1 was P. peruviana, for the farm 2 was stubble low, and for the farm 3 was Solanum tuberosum.
Sampling
Every sampling unit corresponded to 10 plants, in each plant the soil under the canopy and the canopy divided into three fractions (top, mid and low) were sampled. The soil under the canopy was flipped to collect the pupae and caterpillars of noctuids. In each fraction of the canopy, the beam, the underside of 20 leaves as the minimum, and the fruits were inspected, looking for the immature stages of noctuids. All the stages were collected in plastic glasses of 16 ounces, covered with a mesh and marked with the number of the plant, the fraction, the metamorphosis stage and the date. The glasses were put in boxes and carried to the laboratory of the Museo Entomológico Francisco Luis Gallego (MEFLG).
Taxonomic identification
The eggs were placed on moistened paper with water until larvae hatching. The larvae were fed daily with leaves of Ricinus comunis until the adult emerged to be identified. The pupae were left in soil moistened with water until the imago emerged. The moths were put in a lethal chamber with ethyl acetate, to be mounted in an entomological pin. The moths were identified by comparing with the Noctuidae collection of the MEFLG with the curator guide (John Albeiro Quiroz-Gamboa), also comparisons with the taxonomic key of Angulo et al. (2008) were performed. Besides, the lepidopteran specialist of the United States Department of Agriculture (USDA) (Columbus, Ohio) Steven C. Passoa identified two of the species of larvae from this study.
Statistical analysis
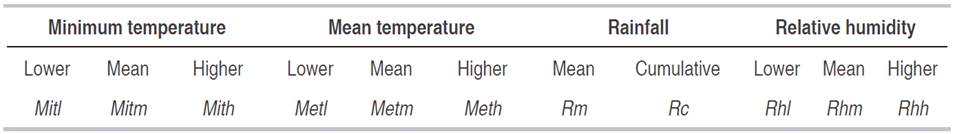
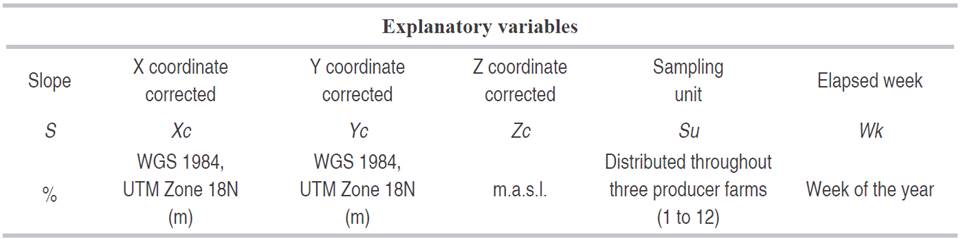
The response variable corresponded to the information of the Noctuidae species that were grouped according to features of the immature stages like pupae, larvae, and oviposition (Table 1). These were correlated with eleven climatic variables, five spatial variables, and a temporal variable. The climatic variables were related to temperature, relative humidity and precipitation, during eight days before sampling, each climatic variable was partitioned according to its features (Table 2). The spatial variable corresponded to the slope and sampling location, north latitude, west longitude, and altitude coordinates. The temporal variable corresponded to the week, correlated with the development phases of the plant as well as the agricultural practices (Table 3). The climatic information was provided by the Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia (IDEAM), and the spatial information was taken with a Global Position System (GPS) Garmin e-trx 10® and analyzed by the software Arcgis 10.2®.
Table 1 Response variables of the immature stages of Noctuidae species associated with Physalis peruviana.
The statistical analysis was made with the Tinn-R® software by using Generalized Linear Models (GLM). GLM is a statistical model for a large family of probability distributions known as the exponential family, that includes important distributions as the Gaussian, Gamma, Chi-squared, Beta, Bernoulli, Binomial and Poisson distributions in order to find statistical differences (Schabenberger and Pierce, 2001). The Poisson distribution was used because the data presented this kind of trend and was of low magnitude. Nevertheless, as there are variables with bigger magnitudes, it was required to get an adjusted model by using Akaike function (AIC) through the step command of Tinn-R®. For the statistical test, it was used the Root Mean Square Predictive Difference (RMSD) that provides a higher sameness to validate the prediction model.
Having into account that projected coordinates have a large scale, it was necessary to apply a correction to avoid masking other explanatory variables, as shown below:
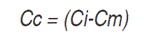
Cc: Coordinate corrected
Ci: Coordinates ith
Cm: Coordinate minimum
RESULTS AND DISCUSSION
In total, nine Noctuidae species were found. They were distributed in the subfamilies Acronyctinae, Noctuinae, Plusiinae and Cuculiinae. The Acronyctinae included the species: Spodoptera albula (Walker, 1857), S. eridania (Cramer, 1782), S. frugiperda (Abbot and Smith, 1797) and S. ornithogalli (Guenée, 1852). These species showed gregarious oviposition covered with the protection of silk and squamae made by the female (Vélez-Ángel, 1997), mainly laid on the leaf underside, in the middle and low fraction of the P. peruviana canopy.
The Noctuinae species were Agrotis ipsilon (Hüfnagel, 1766), Heliothis subflexa (Guenée, 1852) and Peridroma saucia (Hübner, 1808). A. ipsilon has gregarious oviposition mainly on the leaf underside in the middle and low fraction of P. peruviana canopy as well as the Spodoptera species. P. saucia, except for this species also have oviposition on the leaf beam of P. peruviana canopy. Finally, H. subflexa, previously classified in Heliothinae (Matthews, 1991), has isolated oviposition mainly on the leaf beam, in the middle and top of the canopy or close to the reproductive organs like branches or fruits of P. peruviana.
The only Plussinae species was Megalographa biloba (Stephens, 1830), with isolated oviposition mainly on the leaf underside and in the middle fraction of the P. peruviana canopy. The Cuculiinae species found was Copitarsia decolora (Guenée, 1852), with isolated oviposition mainly on the leaf beam, in the middle and top of the canopy or close from reproductive organs like branches or fruits of P. peruviana as well as H. subflexa. C. decolora and H. subflexa could be considered more important species than the other seven reported in this work because they both feed on the fruits because if the fruit is exported with small larvae and quality flaws could be baned. C. decolora and H. subflexa were present mainly at the crop reproductive phase, while the other seven species that feed on leaves were present in all the development phases, mainly in the vegetative stage.
The Modelling
The GLM built for each response variable showed the estimated trend for the different immature stages, considering the explanatory variables that more affected the model. Table 4 shows the statistical significance of the variables that describe the model trend and the Akaike index (AIC) that allow inferring if each model is suitable to estimate the trend of the immature stages.
Before describing each model, it is noteworthy that the most complex model, considering it was affected by 12 of the 17 variables studied, was the oviposition model of C. decolora and H. subflexa (Table 4). It could be explained because their eggs are laid individual and located from the middle to the upper part of the canopy, therefore they are more exposed and sensitive to most of the variables. On the other hand, the simplest model was the pupae one, only affected by four variables, suggesting that the states of Noctuidae located underground are less susceptible to most of the variables studied. Besides, it should be highlighted that the GOLB model of P. saucia oviposition had an index closer to zero.
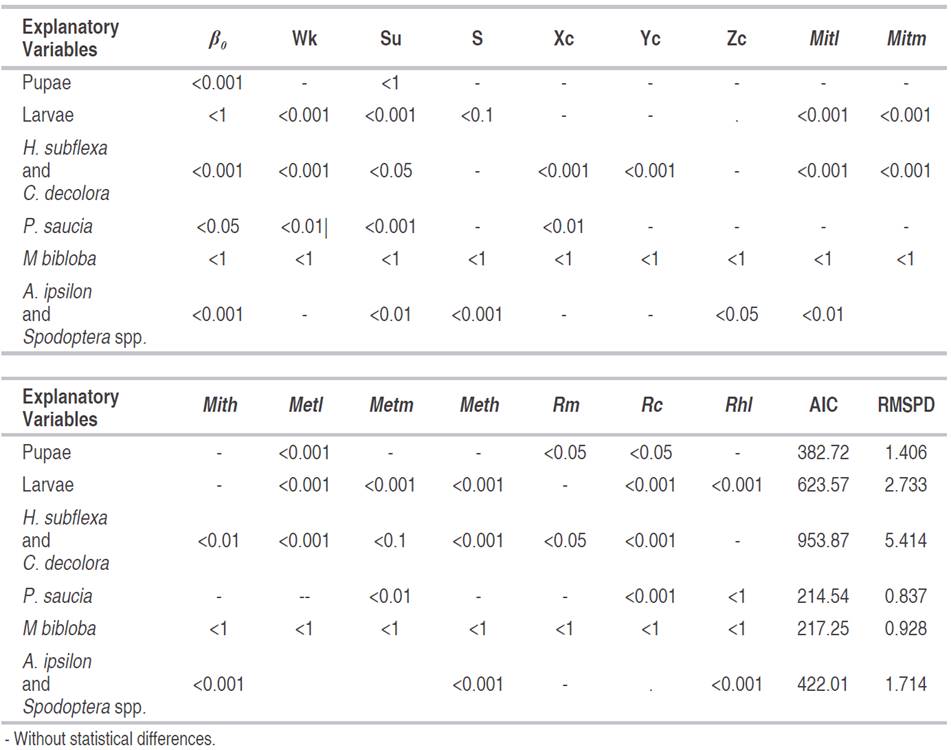
Peridroma saucia oviposition model
The gregarious oviposition of P. saucia on the leaf beam (GOLB) in P. peruviana showed a model with a RMSPD of 0.84, close enough to zero, and all variables had a P<0.05, thus the confidence interval was higher than 95%, so it could be used to estimate the oviposition trend of this moth (Figure 1). This kind of model could be used as a tool for integrated pest management, especially if it is complemented with a development model (Choi and Kim, 2014) to predict the time of the larvae first instar, as the proposed for Ascostis selenaria (Denis et Schiffermüller) (Lepidoptera: Geometridae). Allowing an effective spraying time, since the egg and the first instar are the most vulnerable to insecticides (Park et al., 2014). The equation suggested that the oviposition of P. saucia was influenced negatively by the week (Wk) and by the lower relative humidity (Rhl).
The variable Wk could be related to P. peruviana phenology as well as the agronomic activities because the plant development was matching with chemical pest management to avoid damages in leaves and fruits of P. peruviana. The relative humidity and temperature affected the developmental stages of P. saucia. The relative humidity and mean temperature were 79.79% and 15.13 °C, respectively, thus the equation model was affected negatively by the lower relative humidity. Moreno-Fajardo and Serna-Cardona (2006b) found that the stages are shorter under a relative humidity of 82.93% and a temperature of 23.97 °C than 65.96% and 17.72 °C. On the other hand, mean temperature (Metm), the rain cumulative (Rc), the sampling unit (Su) and the East corrected (Ec) had a positive effect in the oviposition model of P. saucia.
Agrotis ipsilon and Spodoptera spp. oviposition model
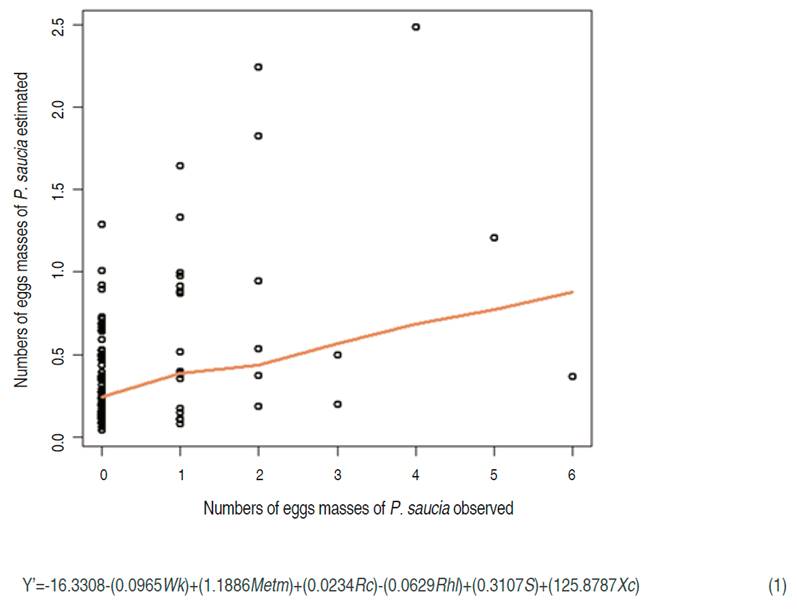
The oviposition model of A. ipsilon and Spodoptera spp., corresponding to the gregarious oviposition on the leaf underside (GOUL) (Figure 2)., had a RMSPD of 1.7142 with a P<0.05 for all the variables. It described closely the oviposition trend of the Spodoptera species found in P. peruviana crop: S. ornithogalli, S. albula, S, eridania and S. frugiperda, as well as Agrotis ipsilon. The oviposition of these species showed similar features because they were found mainly in the mid and low canopy of P. peruviana, and the eggs usually were gregarious and covered with silk and flakes (only those of Spodoptera spp).
Although this model was not as close to zero as that of P. saucia (Figure 1), it could be used to estimate the presence of oviposition of these phytophagous to design a pest management program. The variable affecting the oviposition was the higher value of minimum temperature (Mith) with a mean value of 11.8 °C. According to Milano et al. (2008), the lowest temperature threshold of S. frugiperda is 15 °C and the highest is 35 °C; in both temperatures, the mating frequency is affected. In concordance with Méndez-Barceló (2009) who found that female might laid until 1,000 eggs, but if the temperature reaches 30 °C, the oviposition drops to 386 eggs.
Conversely, our model showed that the highest values of mean temperature (Meth), with a mean of 16.14 °C and the lowest value of minimum temperature (Mitl), with a mean of 8.2 °C had a positive effect in the Spodoptera spp. oviposition process. In this case, it is necessary to evaluate the exposition time for every level of temperature, since it may have a lower exposition time in Meth and Mitl, but for Mith had a higher time of exposition. On the other hand, the other variables that affected the model negatively were the slope (S) and the sample unit (Su), both corresponding to spatial variables. Therefore it can be inferred that there are micro clime and soil variables that are not favorable for the oviposition of Spodoptera spp.
Copitarcia decolora and Heliothis subflexa oviposition model
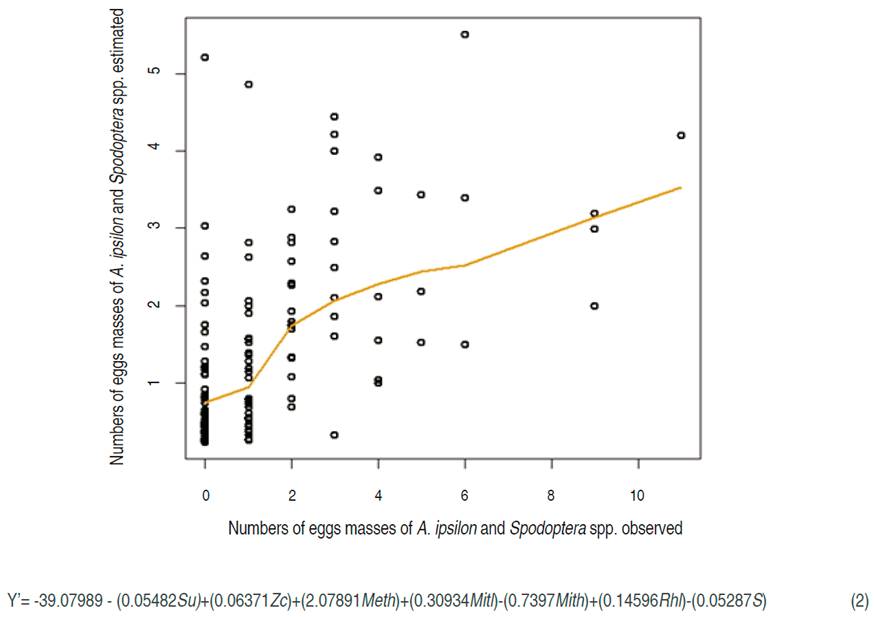
The C. decolora and H. subflexa oviposition model, corresponding to the isolated oviposition on the leaf beam in P. peruviana (IOLB) (Figure 3), had a RMSPD of 5.414, with all the variables with a P<0.05, except for the mean of the mean temperature (Metm) with P<0.1. The highest value of RMSPD allowed inferring that there may be other variables that can explain better the oviposition behavior of piercing fruit moths; despite the statistical test is supported by a high confidence level in each variable. The GLM (Figure 3) is tight enough to describe the oviposition trend for both species; therefore, it could be used to know the immature stages trend of this species.
The equation of the model showed that there was a negative effect on the following variables, with P<0.001: Wk, Mith, Mitm, and Ec. Also, the following variables had a negative impact, with P<0.05: Minth, Rm, Su, and Nc. Moreover, the variable Metm had a negative effect in the model with a P<0.1. Four of the variables named correspond to the lower level of temperature because it could be the most important variable. Moreno-Fajardo and Serna-Cardona (2006a) consider that temperature could affect the number of generations in C. decolora and probably influence the behavior of this moth, like in Alabama argillacea (Lepidoptera: Noctuidae) (Mazza et al., 2006).
On the other hand, the spatial variables Su and Nc, related to the conditions and location of the sampling units, could be associated with soil and microclimatic conditions. Nevertheless, the lowest temperature minimum registered (Mitl), with a mean of 8.02 °C, was positive among the whole variables in this model such as in the Spodoptera model, indicating that there could be a relationship between this level of temperature and the behavior of oviposition in both species.
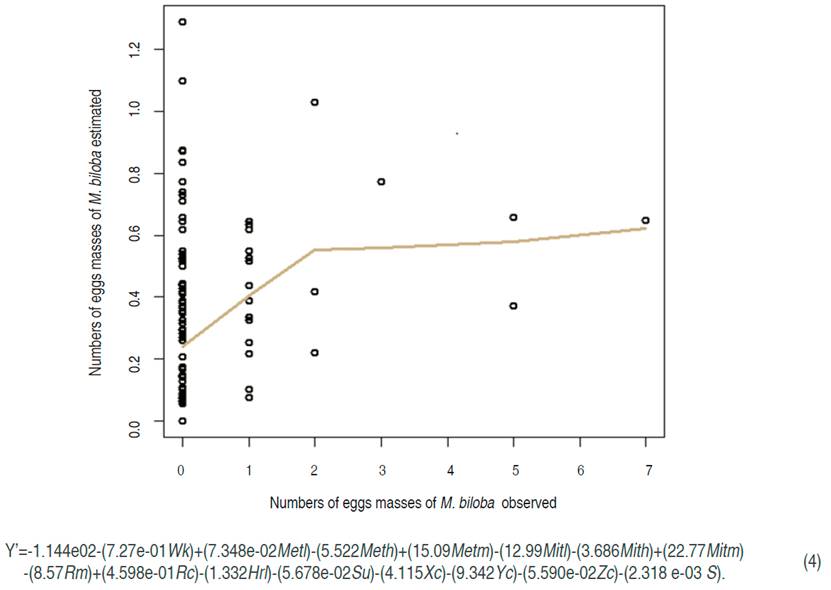
Megalographa biloba oviposition model
The oviposition model of M. biloba that corresponded to isolate oviposition under the leaf of P. peruviana (IOUL) (Figure 4) had a RMSPD of 0.9281; the second model after P. saucia with the lowest value. Nevertheless, the P-value for all variables were close to 0.9 (Table 4) which means a confidence interval lower than 10% in the relationship between the variables studied and the oviposition observed. These results indicated that there is no confidence to estimate the oviposition trend of this species, although Figure 4 showed a good description. The variables studied did not explain the egg trend for M. biloba in the P. peruviana crop.
Larvae model
The larvae model of the nine species of noctuids collected in the P. peruviana crop had a RMSPD of 2.73252 that is a value relatively far from zero, with a P<0.05 for all the variables, except for the slope (S) that presented a P<0.1. Nonetheless, the model had a good trend description of Noctuidae larvae in the crop (Figure 5), turning it in a pest management tool; therefore, with the different variables evaluated it is possible to define a behavior of the Noctuidae larvae populations in P. peruviana.
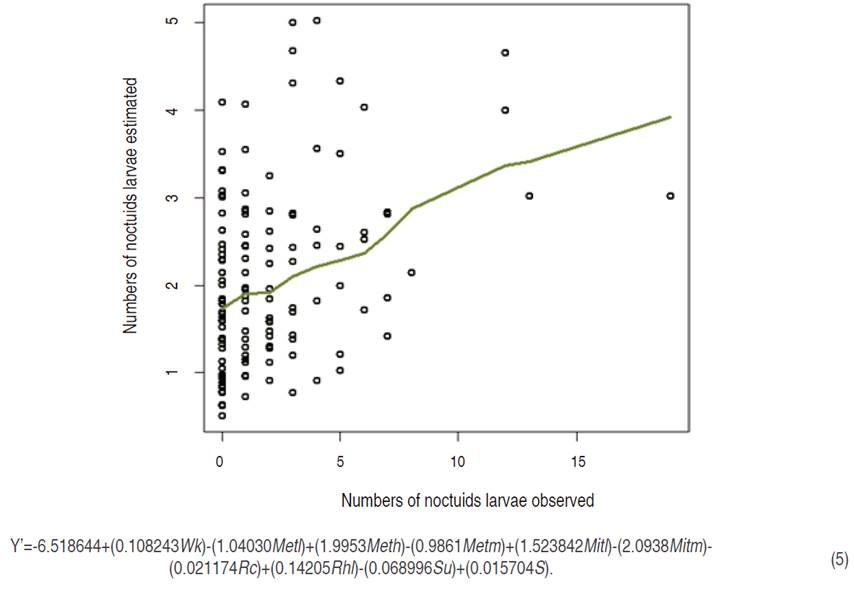
According to the larvae model equation, the lower value of mean temperature (Metl), the mean of mean temperature (Metm), the mean of minimum temperature (Mitm), the rain cumulative (Rc), and the sampling unit (Su) had a negative effect on the larvae presence in the field. The temperature had a high incidence in the metamorphosis, while the rain cumulative suggests that soil water accumulation could affect the larvae behavior in the field. Also, the sampling unit is related to microclimatic conditions and is likely that soil features are correlated as well.
Pupae model
The pupae model of the nine species of noctuids collected in the soil had a RMSPD of 1.4056, which is a value relatively close to zero. The variables had a P<0.05, meaning a confidence interval higher to 95%. Therefore, the model had a great trend to describe the pupae of Noctuidae in the crop (Figure 6).
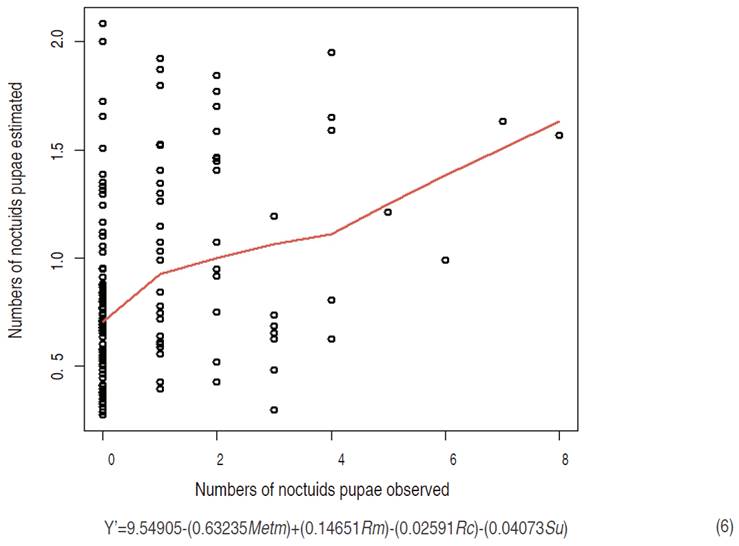
According to the pupae model equation, the mean of mean temperature (Metm), the rain cumulative (Rc), and the sampling unit (Su) had a negative effect in the pupae presence in the field, so as it was observed in the larvae model equation. Considering that the pre-pupae and pupae stages are carried out in the soil, it is important to highlight the possibility that some features of the soil, related to the rain cumulative and the sampling unit, are the reason of the negative effect in this model.
CONCLUSIONS
In most cases, the more important explanatory variables to estimate the trend of noctuids immature stages in the P. peruviana crop were the temperature, rainfall cumulative and the elapsed week. The temperature is known as one of the most important variables because it determines the time for the development of the insects’ immature stages, considering that they are ectothermic. Therefore, the kind of models built in this work, combined with Grades-Day (used to predict the most important thermal events of the insects), should be considered to reduce its populations, if it is necessary. The cumulative rain dropped the larvae populations and probably reduced the imago activity. Finally, the elapsed week of the crop, another crucial variable, should be considered for insect pest management.
The oviposition found for each noctuid species recorded, as well as each model built, constitutes a tool for integrated pest management. The noctuid oviposition agreed with the feeding preference of each species. Those species that feed on leaves laid their eggs frequently on the canopy middle and low part of the leaves, and those that feed on P. peruviana fruit, as C. decolora and H. subflexa, laid their eggs close to the reproductive organs or on it. These behaviors are important for Noctuidae sampling, monitoring and management.

