











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkPlant Genetic Resources (PGR) include genetic diversity in the plant kingdom, with current or potential value (Esquinas-Alcázar, 1993; FAO, 2015). The conservation of these resources ensures the continuous evolution of the plant populations, as well as their adaptation process to different environments. An important part of PGR is the agricultural diversity and all the cropping systems, including those species in domestication process. Thus, one of the main concerns for human well-being is to maintain genetic variation between and within populations (Nodari and Tomás, 2010; Maxted et al., 1997; Rivas, 2001).
One of the stages of conservation of PGR is its characterization, which estimates the existing variability of individuals of a certain population. Characterization, as a first step, is important when the objective is to specify the number of varieties managed by farmers and be able to determine their useful characteristics (Baena et al., 2003; Peñaloza et al., 2010). The Morpho-agronomic descriptors are used to carry out characterization; they are highly heritable attributes, which express themselves in all the environments; they are easy to measure, register, and evaluate (IPGRI, 1980). The descriptors that refer to the form, structure, or behavior of accessions are considered important and useful in describing a sample since they help to differentiate and describe the attribute in a precise and uniform way. An accurate description simplifies classification, storage, retrieval, and use of data. Descriptors are registered in qualitative or quantitative value scales (Franco and Hidalgo, 2003; Abadie and Berreta, 2001). The descriptors must have high discriminating action and low infIuence of the environment for the characterization, which in some cases allows registering the information in situ (Peñaloza et al., 2010; Polanco, 2011). It is recommended to consult the lists of descriptors for the species under study published by Biodiversity International to proceed with characterization (IPGRI, previously).
Most of the characteristics that define a phenotype correspond to the morphological description of the plant and its architecture (Franco and Hidalgo, 2003). These are called morphological descriptors and can be grouped into the following types: (A) Botanical-Taxonomic: which are morphological characters that describe and identify the species and are common to all its individuals. (B) Morpho-agronomics: morphological characteristics that are relevant in the use of the cultivated species, allowing the estimating of the phenotypical variability, quickly and reliable (expressed in visible characteristics) (de Oliveira et al., 2001). These can be qualitative or quantitative and include some of the botanical-taxonomic characteristics as well as others that do not necessarily identify the species but are important from a genetic improvement, agronomic, and market purposes. These characters can be the leaf shape, root pigmentation, stem, leaves and fIowers, color and brightness of seeds and fruits, the architecture of the plant expressed as the habit of growth and types of branching. Descriptors related to the performance components are included because of their economic importance (Franco and Hidalgo, 2003; Abadie and Berreta, 2001). (C) Evaluative characteristics are expressed in response to biotic (pest and disease) or abiotic environmental factors (i.e., temperature, water, and nutrient stress). For example, the evaluation of morphological characters in tangerine "Arrayana" through descriptors allowed establishing homogeneity in a population from Department Meta-Colombia (Orduz-Rodríguez et al., 2012).
The Molecular method is another way of characterization, it is based on the non-visible genetic variability, and it is of great interest in cropping improvement (Abadie and Berreta, 2001; Sánchez-Urdaneta and Peña-Valdivia, 2011). This method is highly sensitive and is not easily performed on promising species due to its cost, absence of molecular markers, and difficult extraction of their DNA. The main objective of characterization and evaluation is the identification and determination of the individuals' agronomical value. Nevertheless, the most reasonable approximation would be the integration of different characterizations, starting with the morphological description (Fernández and Negrillo, 2006).
Sapodilla (Matisia cordata Bonpl.) is a promising species that belongs to the Bombacoideae subfamily (Fam: Malvaceae). It is a native tree from the Amazon. Bees, hummingbirds, and bats visit its hermaphrodite fIowers, and monkeys disperse its seeds in the natural habitats (Tschapka, 2004; Álvarez-Loayza and Terborgh, 2011).
The sapodilla pulp has a sweet and nice taste, with a good content of fiber, potassium (454 mg 100g-1), phosphorus (28.5 mg 100g-1), and calcium (22-25 mg 100g-1) (USDA, 2016). Once the fruit is physiologically mature and harvested, it can be kept for up to one week under shade. The hard consistency of its shell allows the fruit to be in good condition after transport. The wood of the tree is soft and light, used in sawmills, and as firewood. The Sapodilla trees can give shade to cocoa and coffee crops, and their leaves give a good fodder to animals (Geilfus, 1994; Villachica et al., 1996).
Sapodilla gives the identity to the nearby western of Antioquia region and represents an important source of income for local producers, collectors and traders that offer Sapodilla fresh fruit, and juices to the tourists. Currently, the cropping area has been reduced due to changes in the land use caused by the tourist development of the region, without any mitigation project in this situation (CCM, 2016). The region has traditional management of sapodilla crops, with very low technology and yields. Traditional orchards can be constituted by sapodilla trees mixed with other species, such as cocoa, banana, or ornamentals. It is also common to find cropping areas where sapodilla is the dominant species with planting distances between 7 and 9 m. Irrigation is essential since M. cordata is native to the tropical rainforest; therefore, it is intolerant to drought. Annual harvest is generally obtained in the months from July to October, which corresponds to an important income for the families.
Sapodilla is currently cultivated in tropical dry forest (bs-T) area, in the nearby western of Antioquia, Colombia. Although there is little technical information about the crops, farmers of the region have accumulated traditional knowledge and experience with the diversity of this species. They recognize two phenotypes of sapodilla, the "Creole" and the "Ecuadorian," and differentiate them by morphological characteristics such as the size of their fruits, leaves, and the fIavor of their pulp. In Colombia, no studies of morphological characterization of M. cordata are reported. However, in the department of Cauca, two varieties are recognized, one from Ecuador and the other one from Cauca. The Ecuadorian has heavier fruits. On the other hand, the Caucana variety is sweeter, the color of its pulp is brighter, and the fiber content is lower, which makes it more attractive for fresh consumption (Alegría et al., 2007). In the north of Peru, there is a type of sapodilla with low fiber content and superior fIavor (Morton, 1987), and in Ecuador, there is a similar species Matisia ochrocalyx (sapotillo) with smaller fruits (Villachica et al., 1996).
This research aimed to study the phenotypic diversity of M. cordata in traditional orchards of the nearby western of Antioquia and to establish whether there are differences between the Creole and the Ecuadorian genotypes.
MATERIALS AND METHODS
This study was carried out in producing farms in the municipality of Sopetrán (Antioquia-Colombia) (6°30'21.5''N 75°44'24.8''W), in a dry forest life zone (bs-T), with an average temperature of 27.6 °C, an average annual precipitation of 1097 mm, and an average environmental humidity of 73.2% (IDEAM).
The sapodilla descriptors were elaborated using the document "Descriptors for tropical fruits (Descriptores para frutas tropicales)" (IPGRI, 1980) as a guide, since the specific descriptors for M. cordata are unknown. Interviews with producers and harvesters of the region were carried out, and field and laboratory observations were considered. The field guide was elaborated, and a pilot study with a few trees was done to discard the descriptors that did not show any variation or show many variations within the same individual. Finally, the definitive list of the descriptors was obtained.
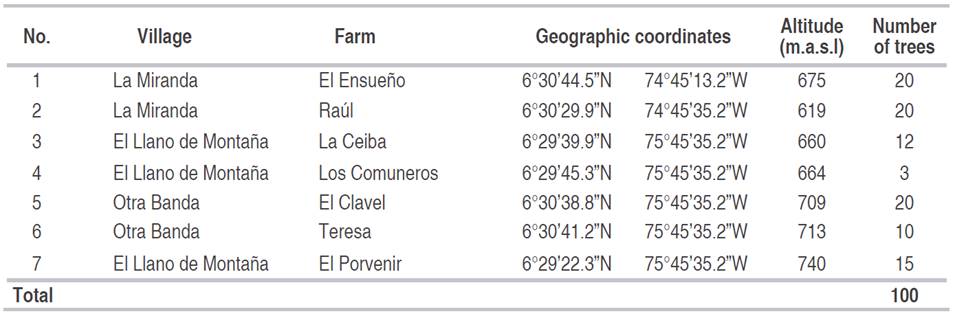
One hundred trees of M. cordata in the reproductive stage, distributed in seven farms from 619 to 740 m.a.s.l., were selected and georeferenced (Table 1).
Table 1 Sites sampled in the municipality of Sopetrán for the characterization of Matisia cordata Bonpl.
Field data
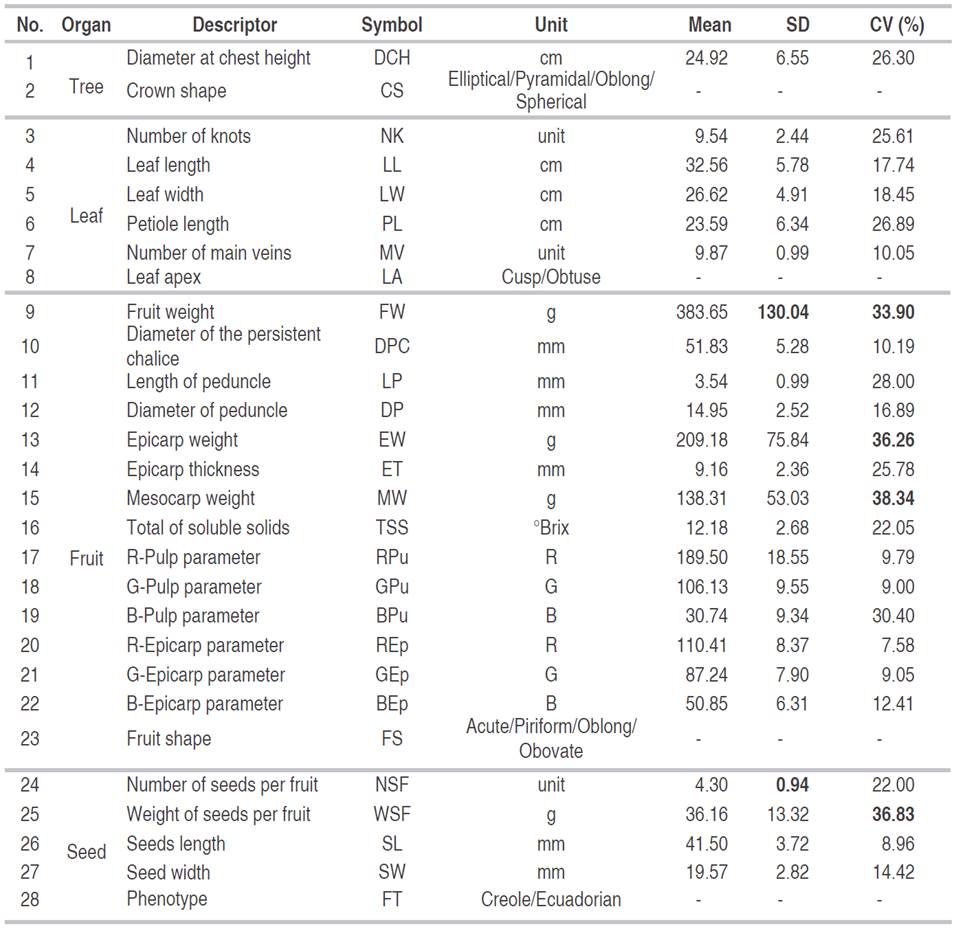
A total of 50 trees of the Creole phenotype and 50 of the Ecuadorian phenotypes were sampled. The Creole or the Ecuadorian phenotypes were established according to the criteria of farmers. Characterization was carried out during August and September 2014. The leaf, the fruit, and the seed descriptors of the plant were evaluated. The characterization process had two phases: in situ and in the laboratory. The qualitative descriptors were the crown shape (CS), the leaf apex (LA), and phenotype (PT). The quantitative descriptors were diameter at the chest height (DCH), the number of knots (NK), the leaf length (LL), the leaf width (LW), the petiole length (PL) and the number of major veins (MV), fruit weight (FW), diameter of the persistent chalice (DPC), length of peduncle (LP), epicarp weight (EW), epicarp thickness (ET), Mesocarp weight (MW), total of soluble solids (TSS), number of seeds per fruit (NSF), weight of seeds per fruit (WSF), seeds length (SL), seeds width (SW). The color was obtained from the measurement with a colorimeter, which performs the measurement in the Red, Green, and Blue (RGB) color scale. These values are expressed in a combination of three numbers whose minimum and maximum values are 0 and 255, respectively. The variables for color were defined as R-pulp parameter (RPu), G-pulp parameter (GPu), B-pulp parameter (BPu), R-epicarp parameter (REp), G-epicarp parameter (GEp), and B-epicarp parameter (BEp). Table 2 records each descriptor and its units of measurement.
Table 2 List of descriptors and general descriptive statistics for the quantitative variables evaluated in Matisia cordata Bonpl.
Five fruits per tree in harvest season were sampled. The evaluated fruits were healthy fruits, in a stage of commercial maturity, which is traditionally defined when the fruit develops a yellow halo around the persistent calyx (Figure 2). Harvested fruits were packed in bags and taken to the laboratory for the second step of characterization.
The second phase, related to the characterization of fruit and seeds, were carried out at the Botany and Plant Physiology laboratory of El Politécnico Colombiano Jaime Isaza Cadavid and at the Food Control and Quality laboratory of Universidad Nacional de Colombia, campus - in the city of Medellin. In order to obtain the color of M. cordata fruits, an X-Rite spectrocolorimeter with CIELAB color spectrum was used, and the RGB scale conversion (0-255) was made using the EasyRGB® (2014) online color calculator.
Phenotypic diversity analysis
The Gower distance (Gower, 1971) was used since it allows simultaneous analysis of quantitive and qualitative variables; distances were used to generate the dendrogram based on the paired grouping method with the unweighted arithmetic mean (UPGMA). In order to know the dendrogram reliability, the cophenetic correlation coefficient (CCC) was calculated. Subsequently, a cut distance was visually established in order to determine the groups. The R statistical environment (R Core Team, 2016) was used to do this statistical analysis.
Comparison of Matisia cordata phenotypes
The Bayesian methodology was used with the implementation of Markov chains in Monte Carlo simulation (MCMC). The mean of a posteriori distribution of the parameter was used as a Bayes estimative. Furthermore, the highest posterior density intervals (HPD) of the 95% of probability was obtained using the CODA package (Plummer, 2015) from the R environment (R Core Team, 2016).
In order to know if there was any difference between phenotypes measures, the difference between each sample of the effective Markov chain was calculated. The respective mean and HPD of the 95% of probability were obtained. For the qualitative variables (Table 2), the proportion of individuals of each category in each phenotype was determined, and such proportions were compared using the Bayesian methodology as described for quantitative variables: the Binom and the Bayesian First Aid.
RESULTS AND DISCUSSION
Phenotypic diversity analysis
Twenty-four quantitative and four qualitative descriptors were established for the discrimination between the Ecuatorian phenotype and the Creole phenotype of M. cordata (Table 2).
The Fruit weight descriptors, FW, EW, WSF, and MW, showed a greater variability with the highest CV values (33.90%, 36.26%, 36.83%, and 38.34%, respectively). Additionally, the FW was the most variable descriptor (SD=130.04).
According to the qualitative descriptors, most of the trees showed pyramidal crown shape (51.0%) (CS), leaf apex (LA), and acute fruit shape (FS) (53%) (Figure 1).
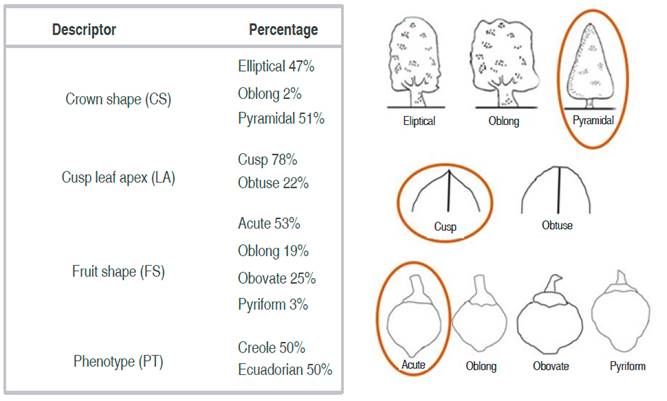
Figure 1 General frequencies of qualitative descriptors for 100 sapodilla individuals (CS: crown shape; LA: leaf apex; FS: fruit shape; PT: phenotype).
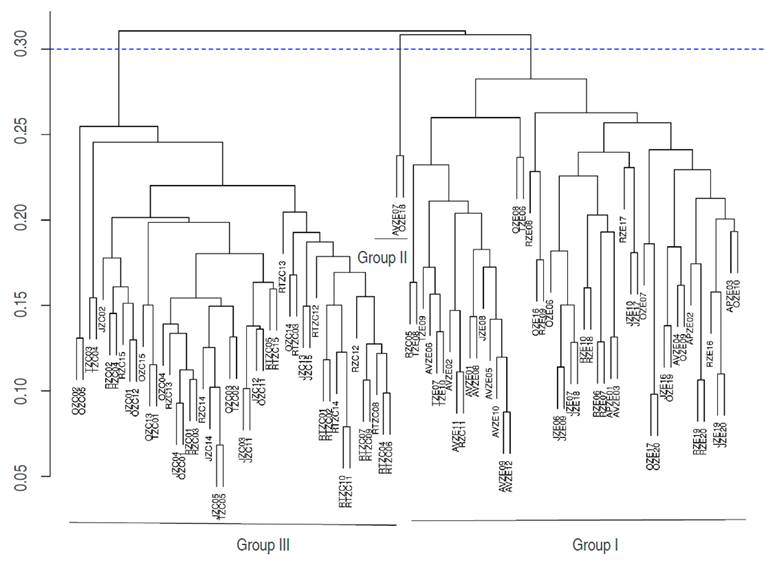
The cophenetic correlation coefficient found was 0.668, indicating that the dendrogram obtained by the UPGMA method represents 67% of the information in the Gower distances matrix, approximately. The dendrogram presents three groups of M. cordata individuals when cutting at a Gower distance of 0.30. The three groups differ by 30% (cut distance:0.30), which is an important percentage to evaluate diversity (Figure 2).
Groups characteristics
Group 1. Formed by 50 individuals, 48 of them belonging to the Ecuadorian phenotype, and two of them to the Creole phenotype. This group is constituted by trees with the pyramidal CS, the largest leaves of the groups (LL:33.54 cm, and LW:28.4 cm), the largest petioles (PL:24.68 cm), and the cuspid apices (LA). The fruit has different forms (FS), the heaviest among the groups (FW:463.08 g), with a high mesocarp weight (pulp) (MW:167.19) and a high number of seeds (NSF:4.4). The fruit has dispersed solid content (SST), reaching 5.7 and 21.8 °Brix values.
Group 2. An atypical group formed by two Ecuadorian phenotype individuals. FW, EW, MW, and WSF descriptors are the most variable descriptors in the group with higher CV values within and between groups. These trees have the thinnest stems (DCH:2.47 cm); the leaves cuspided apex (LA), which are the smallest among the groups; 50% of the individuals show leaves that have less than 20 cm in length (LL:21.15 cm, LW:17.5 cm). The descriptor with the lowest variation is the DPC (CV:4.96%), with fruit with the highest soluble solids content (TSS:16.06); this does not coincide with the 9.0 °Brix found by Alegría et al. (2007) in sampling the Ecuadorian variety in the department of Cauca. The separation of these two individuals could be due to the highest values of TSS and color components, both in pulp and epicarp, being very sweet fruit with pulp and peel of orange and brown-green colors, respectively.
Group 3. Formed by 48 Creole phenotype individuals corresponding to 96% of the Creole trees evaluated, which are distributed in five of the seven sites sampled. This indicates that their origin was not an infIuential variable in the grouping. The trees show the highest average DCH (27.29 cm); therefore, they are older trees with thicker stems. This agreed with the versions of the local farmers, who affirm that the Creole sapodilla arrived first to the region than the Ecuadorian phenotype. The fruit is the lightest among the groups (FW:297 g), with a lighter epicarp, a lighter mesocarp, and lighter seeds. Nevertheless, Bajaña (2016) found that the lightest fruit weighed 324.08 g in Naranjito, Ecuador, while in Villachica et al. (1996) found fruits of 200 g; the fruit shape is acute (FS acute:93.8 %), their pulp is less sweet with the lowest average value of SST (11.8). WSF and the BPu are the descriptors with the greatest variation.
The Group 2 presented the highest color components values in the epicarp (REp:122.5, GEp:94.37, BEp:58.55) and in the pulp (RPu:191.8, GPu:111.7, BPu:33.6), it is to say that they have very light-colored peels and pulp. Individuals in this group showed an extreme color data, possibly contributing to the separation of the other groups. The Group 3 presented the lowest values of color parameters in both the epicarp (REp:107.1, GEp:84.37, BEp:49.8) and the pulp (RPu:188.5, GPu:104.9, BPu:30.3), which corresponds to opaquer colors, i.e., a darker brown-green peel and a pale orange pulp. Group 1 presented intermediate values in color parameters; however, the values are higher than those of Group 3, i.e., the fruits have lighter tones (Table 2 and Figure 3).

Comparison of Matisia cordata phenotypes
When comparing both phenotypes, 64% of the variables showed significant differences (16 quantitative and two qualitative) according to HDP intervals of 95% of probability (Table 3).
Table 3 Comparison of the Creole phenotype and Ecuadorian phenotype of Matisia cordata Bonpl., according to the statistical significance descriptors.
When comparing the crown shape of the tree, the elliptical crown shape (61%) and the pyramidal crown shape (61%) predominated in the Creole and the Ecuadorian phenotype, respectively. On the other hand, 86% of the fruits with an acute shape were of the Creole phenotype, and the 98% of the obovate shaped were of the Ecuadorian phenotype (Figure 4).
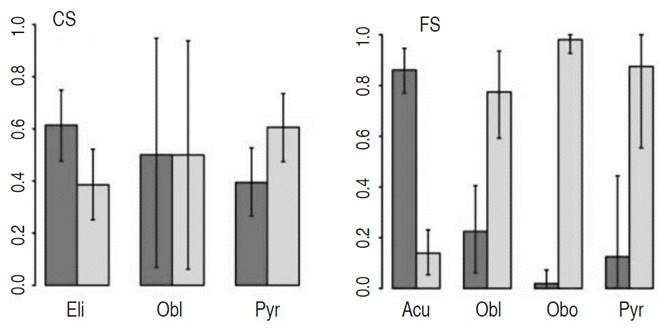
Figure 4 Comparison of the Creole and the Ecuadorian phenotypes of Matisia cordata Bonpl., according to the crown shape (CS) and the fruit shape (FS). The error bars indicate the limits of 95% of a posteriori high probability interval. The black and grey bars correspond to the Creole (C) and the Ecuadorian (E) phenotype, respectively.
In general, the Creole phenotype is characterized by a thicker stem (DCH:28.8 cm), a higher number of knots (NK:10.0), and smaller leaves (LW:25.0 cm). The trees of the Ecuadorian phenotype presented thinner stems (DCH:22.0 cm), and larger leaves (LW:28.0 cm). This coincides with the appreciations of local people, who say the Ecuadorian sapodilla tree was introduced to the region after the Creole. This indicates that these trees are younger than Creole ones; therefore, they have thinner stems. Bajaña (2016) described these phenotype leaves of the Ecuadorian with a smaller width than 24.0 cm, while the trees in Sopetrán-Antioquia presented larger leaves (LW:28.0 cm). The Creole sapodilla fruit had thicker peduncles (DP:15.6 mm) and thinner epicarp (ET:8.5 mm).
The Ecuadorian sapodilla showed a higher average of TSS (12.6) than the Creole phenotype (TSS=11.6). This does not match with the findings of Alegría et al. (2007), who found values of 9.0 °Brix for the Ecuadorian variety and 11.0 °Brix for the Caucana variety in the department of Cauca (Colombia). The Ecuadorian phenotype has higher values than those reported in the department of Cauca, which may be due to the environmental conditions of this region. The information found does not agree with the traditional differentiation in the region that considers the fruits of the Creole variety sweeter than the Ecuadorian one.
The Creole phenotype was lighter (FW:301.0 g), also the epicarp (peel) (EW:162.8 g), the mesocarp (pulp) (MW:108.6 g), and the seeds (WSF:30.0 g). Additionally, the seeds were slightly smaller (SL:40.0 mm and SW:17.7 mm) than the Ecuadorian phenotype. Alegría et al. (2007) found differences in fruit weight between the Caucana variety (lighter fruit, 197.9 g) and the Ecuadorian variety (heavier fruit, 500 g). Although the Ecuadorian phenotype is heavier (WS:465 g), findings in the Cauca variety (Colombia) showed heavier fruit, which leads to inquiring about the origin of the Ecuadorian phenotype. Differences between the Caucana variety and the nearby western sapodilla of Antioquia suggest that a study comparing the two producing regions is necessary to determine the characteristics of each variety or regional phenotypes.
Both the Creole and the Ecuadorian phenotypes showed differences in the color of the pulp and the epicarp. The Creole sapodilla showed lower values in the red and in the green components of the pulp color (RPu:185.6, GPu:104.6) and of the epicarp (REp:107.4, GEp:85.2), which give to the Creole less intense tones than the Ecuadorian fruit (Figure 4). The more intense orange color was found in the Caucana variety in the department of Cauca, Colombia (Alegría et al., 2007). This does not agree with the color results obtained in this study, as well as the appreciations of the interviewed people, who perceive the pulp of Creole sapodilla with a more intense color than the Ecuadorian pulp color.
In general, the Ecuadorian sapodilla showed better productive characteristics because it produces heavier fruit with greater pulp weight and a more attractive pulp color for the consumers. The fact of having wider leaves could infIuence the photosynthetic capacity of trees, making them more productive. In addition, this phenotype presented a large diversity of fruit shape, which questions the intraspecific diversity in the studied region.
CONCLUSIONS
The Ecuadorian sapodilla from the Nearby western of Antioquia was characterized by having larger leaves, diverse shaped and heavier fruit, and a heavier epicarp and pulp. It also has larger and heavier seeds than the Creole phenotype from the same region, which presented the fruit of acute shape, with pale orange pulp and epicarp of brown tonalities, and an elliptical-shaped tree crown. The Ecuadorians presented more intense orange pulps, dark green epicarps, and trees with Pyramidal crowns. The local knowledge of phenotypic variability of the sapodilla in the region is partially confirmed because the color and the fIavor descriptors of the pulp did not agree with the local perception.
There is a lack of information and studies of M. cordata. Therefore, this study corresponds to pioneering and base work in Colombia, and it will be useful in the application of the proposed descriptors for future studies. This morphological study could be complemented by other molecular and/or morphometric works to support the obtained results better.

