











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkCarludovica palmata is a plant that belongs to the Cyclanthaceae family. It is typically found in the Neotropical region in the Americas, ranging from the northern provinces of Perú and Mexico (Bennett et al., 1992). In Colombia, this species is called "Palma de Iraca" (Iraca palm), and inhabits forests in altitudes from 900 to 1,800 m.a.s.l. and temperatures from 22 °C to 28 °C. This plant exhibits great economic potential for many families that live in the rural areas due to its use as a non-wood forest product (te Velde et al., 2006; Galviz-Quezada et al., 2019), an industrial product (Linares et al., 2008), an alimentary product (Álvarez, 2014), or a possible source of plant fibers in bio-composites (Bourmaud et al., 2018). Its reproduction can be sexual through seeds; however, germination problems associated with pollination events have been reported (Gómez et al., 2011). Therefore, C. palmata is usually asexually reproduced by rhizomes or underground stems, which have several apical meristems with pyramidal shapes. The Cyclanthaceae family is comprised of monocotyledons, and it is characterized by having leaves that exhibit folded blades joined to a petiole that sprouts from the rhizome on the ground level. In the specific case of the iraca palm, the leaves are recognized by having a palmate shape (Wilder, 1977). When the plants are young, they form leafy shoots known as buds, whose fibers are used to produce the traditional Panama hat (Linares et al., 2008) and other handicrafts of economic importance. These products are manufactured in small Colombian cities. For instance, in La Unión, Genova, Linares, and Sandoná located in the Nariño Department (Reyes, 2007); Aguadas in the Caldas Department; Usiacurí in the Atlántico Department (Córdoba and Portilla, 2005); Arusí in the Chocó Department (Muñoz and Tuberquia, 1999); and Suaza and Guadalupe in the Huila Department.
The diminish of agricultural production in Colombia (SAC, 2011) and the slow growth of the economic sector in the last few years have affected the production of C. palmata, causing low production of raw material and affecting the handicraft production chain negatively. A protocol for the mass multiplication of C. palmata is needed to satisfy its demand. A plant tissue culture is a group of techniques that allow the multiplication and rooting of plant material (Davey and Anthony, 2010). This important technology of mass multiplication has been applied with much success in medicinal plants (Fallah et al., 2019), as well as industrial plants and edible crops. In clonal multiplication, the ability of the plant to generate new plants from buds is potentialized through hormonal relations. This effect is achieved through the presence of plant growth regulators such as cytokinins. The generation of roots in the protocol of clonal multiplication in vitro is important because it allows the adaptation of the plants to ex vitro conditions.
The aim of this work was to determine the effect of 6-benzylaminopurine (BAP) in the formation of new shoots and 1-naphthaleneacetic acid (NAA) in the formation of roots of Carludovica palmata Ruíz & Pavón, as well as its adaptation in greenhouse conditions.
MATERIALS AND METHODS
Plant material and seed sterilization
The plant material used in this study was obtained from 504 seeds of plants located on the campus of the Universidad Nacional de Colombia in Medellin (6°15'44"N 75°34'37"W). Dried and mature seeds were collected and submerged in an aqueous solution 3% (w/v) of NaClO with five drops of Tween 20® for 20 minutes in a laminar flow cabinet. Afterward, the seeds were washed in a solution of ethanol 95% (v/v) for one minute. Any amount of alcohol left in the seeds was eliminated by washing them three times with sterile distilled water.
Establishment of in vitro cultures
Ten seeds were placed in each glass jars containing 15 mL of a semi-solid Murashige and Skoog culture medium (MS) (Murashige and Skoog, 1962), supplemented with 0.5 mg L-1 of pyridoxine, 0.5 mg L-1 of thiamine, 0.5 mg L-1 of niacin, 2 mg L-1 of glycine, and 100 mg L-1 of myo-inositol. Sucrose 2% (w/v) was used as the source of carbon and IAA (0.025 mg L-1), BAP (0.02 mg L-1), and gibberellic acid (GA3) (0.02 mg L-1) were added as growth regulators. The pH of the medium was adjusted from 5.8 to 6.0, with drops of aqueous solutions of NaOH. Finally, 1.8 g L-1 of phytagel were added as a gelling agent. 20 glass jars were used to set 10 seeds per glass jar; they were kept in a culture room with controlled temperature conditions (22±1 °C) and a photoperiod of 12 h for 60 days. The explants used in this work (Figure 1B) were obtained from the resulting seedlings of the germination; then, they were placed in glass jars containing 15 mL of culture medium.

Culture media and in vitro propagation
Bud induction. For in vitro micropropagation, the same MS basal medium with the same proportion of vitamins, sugars, and the gelling agent was used. The medium was supplemented with different concentrations of BAP (Table 1) to evaluate the rate of bud induction.
Table 1 Hormonal treatments with BAP to evaluate the rate of bud induction and NAA to measure root formation of iraca palm under in vitro conditions.
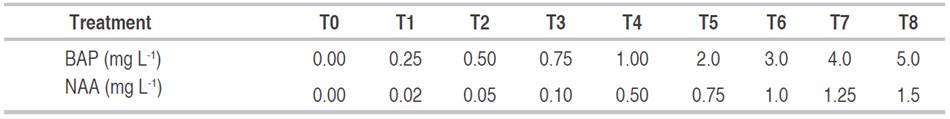
The experiments were randomly carried out with nine treatments; they were applied to seven glass jars each with 15 mL of culture medium. Four explants were planted in each glass jar and kept under the same conditions (temperature and light exposure), used in the first establishment of the culture, for 90 days. After each treatment, the number of shoots per explant (NSE), the length of shoots (LS), and length of roots (LR) were determined by manual counting.
Root formation. Different concentrations of NAA were added to the MS modified medium as a root generation factor in each treatment (Table 1). The experiments were randomly carried out with nine treatments. They were applied to seven sterilized glass jars with 15 mL of culture medium each one. Buds were put in each glass jar and then kept under the same constant conditions (temperature and light exposure) as the ones described when establishing the parameters for the in vitro culture. After each treatment, the number of roots per explant (NRE), length of plantlets (LP) and the length of roots (LR) of each explant was obtained by measuring its longest root counting from its tip to the base where the root system ends, and the stem begins.
Establishment of plantlets to ex vitro culture
The in vitro plantlets obtained from the rooting process were used for this stage. The selected plantlets were washed with distilled water to eliminate any residual culture medium and then submerged for 10 min in an aqueous solution with the fungicide propamocarb hydrochloride at 0.1% to prevent fungal proliferation. For the acclimating experiments, a factorial array of 2×3 in a completely randomized design was used. The first factor was the fertilization with two levels (with and without fertilizer), and the second factor was the composition of the substrate with three levels: I) soil-rice husk in a proportion of 1:1, II) peat, and III) the mixture soil-peat-rice husk in a proportion of 1:1:1. There were carried out six treatments with five repetitions each, totaling 30 experimental units, being each growth chamber an experimental unit. The fertilizer used was 15-15-15 (Triple 15), formulated at a concentration of 0.5 g L-1. The substrates were deposited in plastic containers where they were sterilized with boiling water and left to cool down for an hour; this procedure was performed twice. Then, the fertilizer was added to those substrates, which required it as instructed in the experimental design. The fertilized substrates were then taken to the humidity chambers to initiate the process of sowing the plantlets. The humidity chambers built for this investigation are closed containers where the process of acclimation happens due to its ability to maintain high relative humidity, which helps the plant to tolerate the transition to in vivo conditions (Read and Fellman, 1985). Each of the chambers consists of two 16-oz plastic cups (Figure 2C) that were previously sterilized using sodium hypochlorite at 2% (w/v) for 10 min. The plantlets were cultivated for 80 days, in which there was measured the growth and development of the plantlets. In days 0 and 80, the length of its leaves and roots, as well as its wet weight, were measured. The chambers were kept at room temperature in greenhouse conditions.

RESULTS AND DISCUSSION
Establishment of in vitro culture
Plantlets were established after the in vitro germination of the seeds. The in vitro germination rate was 87.9±3.03%. In Colombia, this species is exclusively asexually propagated from rhizomes (Córdoba and Portilla, 2005), mostly due to the low germination rate of the seeds in vivo. Unlike in vitro cultures, in vivo cultures in soil do not have controlled conditions of pH, temperature, availability of nutrients, or humidity. The use of growth factors presents in the semisolid media (IAA, BAP, and GA3) may have infIuenced some phenomena, such as the physiological latency of the seeds (Baskin and Baskin, 2014). These events could be successfully established in an in vitro culture to develop a product that allows more efficient multiplication of C. palmata.
In vitro propagation
The resulting plantlets of the in vitro propagation are shown in Figure 1A. It is worth noticing that the plantlets were vigorous and did not exhibit the presence of necrotic tissue in their roots or stems.
Table 2 shows that a BAP concentration lower than 1 mg L-1, the NSE was low, whereas, in higher concentrations, the multiplication rates increased, with more than 10 shoots per explant. The highest NSE was obtained (17±3 shoots per explant), and it was achieved with 2 mg L-1 of BAP. The large proportion of new regenerated shoots might be due to the ease that some plants have to respond to hormonal and environmental conditions, as in the case of the iraca palm.
Table 2 Effect of the concentration of BAP in the formation and development of new shoots from seedlings of C. palmata.
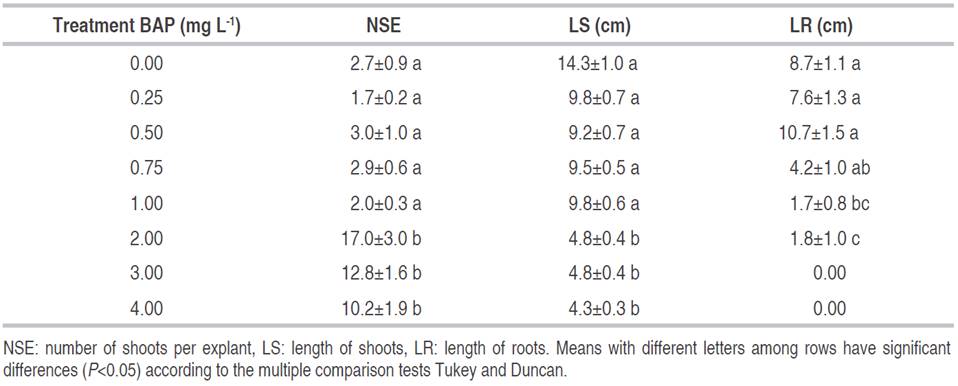
It was found that the highest LS with a value of 14.3±1.0 cm was the control (without hormones), as it is shown in Table 3. Similar results were observed when BAP was used as a growth regulator for the induction of shoot growth when multiplying the medicinal species Zhumeria majdae (Fallah et al., 2019) and other plants of industrial interests, such as bamboo and Hibiscus cannabinus L. (Herath et al., 2004; Ayadi et al., 2011). Furthermore, it can be noticed that the length of the shoots decreases as the concentration of BAP increases. This response would be associated with a physiological factor of the plants related to the redirection of energy to generate biomass: at a greater number of shoots, the shorter those are, and vice-versa.
Currently, there are no reports about the in vitro propagation of C. palmata. For this reason, there was propose to add cytokinin BAP to achieve an efficient multiplication of this plant. This phytohormone has been used to generate buds in an important amount of commercial vegetal species and forest trees. When observing the effects of BAP in the multiplication and elongation of shoots, it can be noticed that while the maximum number of shoots is achieved, their length diminishes. This behavior might happen because lower concentrations of BAP in the medium cause the biomass production to be more directed towards elongating the shoots rather than generating new ones, whereas, in the presence of concentrations of BAP higher than 2.0 mg L-1, the biomass production would be directed to the generation of new shoots. This can be noticed when observing the smaller shoots in the presence of higher concentrations of BAP and the larger shoots in lower concentrations of BAP, as described by George et al. (2008). This phenomenon could be related to the ability of the cytokinins of intervening in the cellular cycle, encouraging the synthesis of cyclin-dependent kinases, which act upon the transition G2/mitosis. In turn, this encourages cellular division, and all reactions that tend towards said division require a certain amount of energy to be spent (Azcón-Bieto and Talón, 2013) to generate new vegetative shoots (Jurado, 2010; Miller et al., 1955).
Root formation
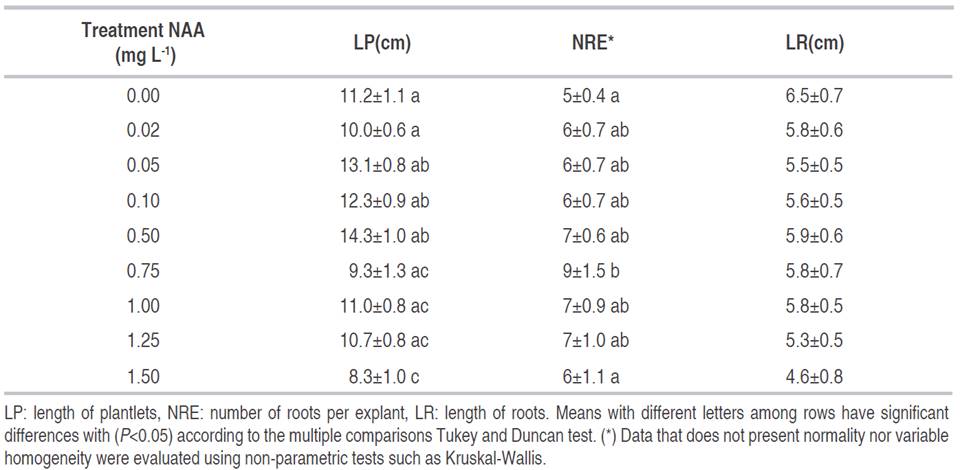
The LR did not present any significant difference for the treatments with NAA. Roots were present in every treatment, including the control subject. On the other hand, the treatment of NAA at 0.75 mg L-1 was the one that presented the highest NRE with a value of 9.0, as shown in Table 3.
The highest LR was observed in the treatment of 0.5 mg L-1 of NAA (5.9 cm) as well as in the control subject (6.5 cm). These results show that in the experiments of clonal multiplication of iraca palm, root generation would not be a problem. Lastly, as shown in Table 3, it can be observed that in shoot formation, there were no significant differences across treatments. These events suggest that C. palmata does not present resistance to rooting and might not require hormones due to the presence of endogen auxins (Figures 2A and 2B). This behavior of iraca palm clones when rooting facilitates the process of hardening the plant for in vivo conditions. It was also found that in the presence of high concentrations of auxins, there is no effect over the growth and development of the root system (Overvoorde et al., 2010).
Adaptation to ex vitro conditions
One hundred percent of the plantlets that were transferred to the humidity chambers survived after being planted for four weeks (Figure 2C). The plants were planted in bags of soil and continued to grow on free exposition without presenting signs of deterioration or death (Figure 2D). This could be due to all in vitro grown plants had an adequate growth of shoots and mainly of roots, which facilitated the acclimation process (Asadi et al., 2009; Shekhawat et al., 2015).
The data, corresponding to the effect of different substrates in the adaptation of the plantlets, are reported in greenhouse conditions (Table 4). In the case of the growth of the aerial part, the use of triple-15 fertilizer aided the growth of the foliage considerably, unlike the substrate soil without fertilizer registered the lowest values of the growth of the aerial portion. The mixture (soil-peat-rice husk) in two presentations (with and without fertilizer) proved to be better than soil-rice husk or the peat substrate on their own, although no differences significant enough to establish the presence of the fertilizer as a determining factor in foliage growth were found.
Table 4 Effect of different substrates over the adaptation of plantlets of C. palmata between day 0 and day 80 of being in greenhouse conditions.
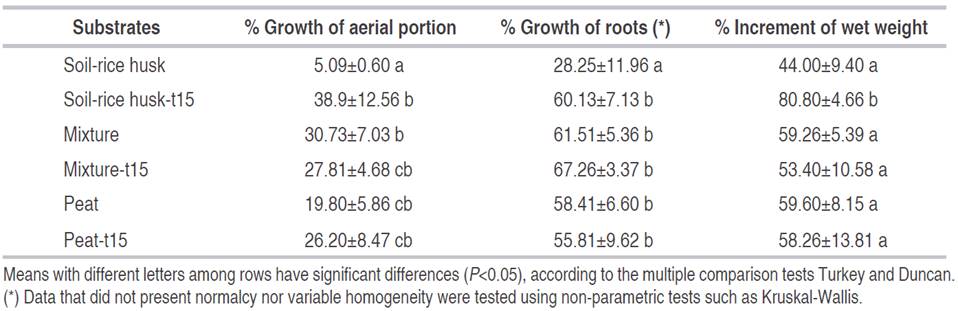
In terms of root growth percentage, it was found that the substrate soil-rice husk without fertilizer exhibited the lowest root production with 28.25±11.96%, being significantly different compared with other treatments (P<0.05). For all other treatments, it was found that, although an increment in the percentage of growth was found with triple-15 fertilizer, the results were not significantly different. As for the wet weight, an increment of 80.80±4.66% was found in the plantlets that were planted in the substrate soil-rice husk-t15, which was higher than other treatments (P<0.05). This difference could be explained by the contribution of macronutrients present in the fertilizer. Similarly, the physical structure of the soil substrate, which allowed better adaptation of the plantlets; however, it could be rather poor in regard to the concentration of the nutrients present in it, nutrients necessary to generate a strong vascular system, and greater foliage.
It is noteworthy that synthetic fertilizers, such as triple-15, are a relation of macronutrients that contain 15% (w/w) of nitrogen, phosphorus, and potassium, respectively. Nitrogen intervenes in processes of synthesis of proteins, chlorophyll, sugar conversion, and facilitates the absorption of nutrients. Phosphorous is related to the metabolism of coenzymes (NAD+, NADP+, FAD+, thiamine pyrophosphate), ATP synthesis, promotes root development and cellular division and growth. Lastly, potassium is related to enzyme action and therefore intervenes in processes of protein synthesis participating in the formation of roots and in the ability to open and close stomas (Rozov et al., 2019; Zahoor et al., 2017). It also regulates the transpiration of water in periods of drought - this translates into the growth of foliage and the plant's vigor, incrementing its biomass (Zahoor et al., 2017).
On the other part, the sample of soil taken for this experiment is similar to the soil in which the plant would develop naturally, so the relation between plants and microorganisms could be considered. In turn, this might have manifested in the increment of the biomass in the substrate soil-rice husk-t15. These relationships are beneficial for the absorption of minerals and nutrients, such as phosphorus, nitrogen, zinc, calcium, and potassium (Gonçalves de Oliveira et al., 2011; Smith and Read, 2008), and that increments tolerance to environmental stress. Therefore, those macro- and micronutrients are beneficial because they are absorbed, and the growth and vigor of the plantlets improve. The increase of root formation and the strengthening of stomas increases water absorption and nutrients and regulate water loss via transpiration. This process might increase the plant's biomass because the presence of water that has been retained increases its wet weight.
CONCLUSIONS
In this study, it has been developed an efficient in vitro micropropagation system for the multiplication and root formation of the iraca palm. The use of BAP and NAA had an important effect on this purpose. In vitro grown plants successfully adapted to ex vitro conditions; thus, the first report of the in vitro micropropagation of the iraca palm (Carludovica palmata) has been informed. This multiplication protocol through biotechnological tools might contribute as the base of a future in vitro germplasm bank of iraca palm.

