











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkThe optimum harvest time can be defined as a balance between the production of dry matter, nutritional quality, and grass persistence. As the age of the pasture increases, the production of dry matter increases, but the nutritional quality declines due to a reduction in the content of crude protein and digestibility and to the rise of neutral detergent fiber and acid detergent fiber (Canseco et al., 2007). As the plant begins senescence, the accumulation of dead forage occurs, and in some cases, leaf-stem ratio decreases, reducing the proportion of green forage allowance from the animal (Anwandter et al., 2007). This balance between quality and yield is achieved with adequate management of defoliation frequency, determined by the optimum harvest time. Additionally, defoliation frequency infIuences the number of reserves that the plant has for regrowth. When the pasture is defoliated early, in phenological age, it is possible that carbohydrate reserves and post grazing leaf area index are not enough to hold the subsequent regrowth, delaying growth and decreasing the annual production of dry matter, and sometimes, it can generate degradation of the pasture. On the other hand, if the pasture is defoliated too late, nutritional quality decreases, and the rate of accumulation of green forage is reduced (Chapman et al., 2011).
The optimum harvest time, where adequate dry matter production, nutritional quality, and pasture persistence can be achieved, cannot be assigned based on the chronological age. The plants grow at different rates depending on environmental conditions, in particular on the environmental temperatures (Andrade et al., 2015; Herrero et al., 2000). Therefore, plant characteristics have been identified (phenological state, fIag leaf, among others) to make easy decision rules of the optimum harvest time to be used by the producer. One of these characteristics is the number of living leaves that remain in a stolon or a tiller, before the beginning of the senescence of the first leaf that emerged after grazing (Chapman et al., 2011). The number of green leaves that a tiller can support is constant and depends on the forage species (Fulkerson and Lowe, 2003). Leaf appearance rate is defined as the interval between the emergence of two leaves successive in the same tiller. Although this interval can be expressed in days, due to its close relation with environmental temperature, it has been calculated, preferably as thermal summation (phyllochron) (Wilhelm and McMaster, 1995). Knowing the phyllochron of a forage species allows making recommendations about defoliation frequency, considering the pasture phenological age and the mean temperature of a specific location.
Reeves and Fulkerson (1996) found for Kikuyu grass (Cenchrus clandestinus (Hochst. ex Chiov.) Morrone), the main forage species in high altitude tropics of Colombia (Mila and Corredor, 2004), that between 4 and 5 leaves per tiller, the grass is at the point of optimal nutritional quality and has enough carbohydrates reserves for the next regrowth. After this number of leaves, they found that the quality of Kikuyu began to decrease due to the senescence of older leaves. However, Fonseca et al. (2016) (in the Department of Boyacá, Colombia, at 2560 m.a.s.l.) concluded that the quality of Kikuyu did not change significantly between 4 and 6 leaves per tiller and the highest dry matter yield was reached in the stage of 6 leaves.
Herrero et al. (2000), using a growth model for this species, suggested that the optimal number of leaves per tiller to harvest could be infIuenced by the level of nitrogen fertilization. Without nitrogen fertilization, the model estimated 7 as the optimum number of leaves per tiller to harvest Kikuyu. At fewer leaves per tiller the forage mass required for grazing (>2500 kg dry matter ha-1) would not be reached. With N fertilization (100 kg N ha-1), the number of the leaves needed to have enough dry matter yield for grazing would be lower (4 to 6). These authors suggested that N fertilization increases the rate of leaf elongation and, at the same time, the rate of forage mass accumulation, reducing the time required to accumulate enough dry matter yield for grazing. A positive response in the growth rate induced by N fertilization has been proven in other studies with forage grasses (Borrajo and Alonso, 2014; Almeida et al., 2011). This response is more related to the effect of nitrogen on the production of cells rather than its elongation, although this mechanism is still under discussion (Gastal et al., 2015).
According to the above mentioned, the objective of this study was to determine the optimal harvest time of Kikuyu grass using as criteria the number of living leaves per tiller and to establish if the N fertilization affects this number.
MATERIALS AND METHODS
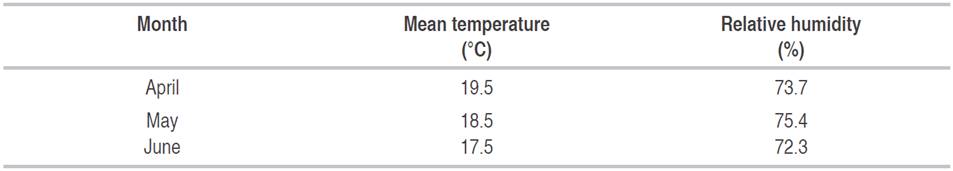
The experiment was carried out from December 2015 to June 2016 at the Facultad de Ciencias Agrarias of the Universidad Nacional de Colombia, Bogotá, at 2600 m.a.s.l. and an average daily temperature of 14 °C.
The experiment was conducted in pots in a greenhouse, with a permanent record of temperature and relative humidity (Table 1).
Kikuyu grass stolons were planted in 96 plastic pots (34 cm×27 cm×12 cm), with a density of 12 stolons per pot, using the same kind of soil (from high altitudes >2,800 m.a.s.l.) for all pots. This soil presented dark coloration, high content of organic matter (13.8%), strong acidity (pH 5.4), suitable Cation-Exchange Capacity (54.8 meq per 100 g), and clay loam texture, aspects similar to those reported for soils from the bleak Colombian uplands (IDEAM, 1999). Irrigation was applied to maintain soil moisture at field capacity. Kikuyu stolons were allowed to grow until they covered at least 80% of the pot. This period lasted 4 months. At this time, uniform defoliation was made at 3 cm from the ground.
Treatments and variables
Sixteen treatments were evaluated in a 4×4 factorial design, with three repetitions per treatment; each repetition consisted of 2 pots. Leaves per tiller at the time of harvest and the level of nitrogen fertilization were the factors. Four harvest treatments considering the leaves per tiller 3 (L3), 4 (L4), 5 (L5) and 6 (L6), and four levels of nitrogen fertilization, 0 (N0), 50 (N50), 100 (N100) and 150 (N150) kg N ha-1 year-1 were studied. The nitrogen source used was urea. Additionally, applications of phosphorus (69 kg ha-1 year-1), potassium (90 kg ha-1 year-1), and magnesium sulfate (25 kg ha-1 year-1) were made, according to the requirements of the Kikuyu (Bernal and Espinosa, 2003) in all treatments.
Dry matter (DM) yield (extrapolated to t ha-1) of leaves, stems, green forage (GF, leaves+stems), dead forage (DF), leaf-stem ratio (L:S), and green forage-dead forage ratios (GF:DF) were the variables evaluated. Also, the content of crude protein (CP), neutral detergent fiber (NDF), acid detergent fiber (ADF), lignin, ether extract (EE) and ash were determined for leaves and stems.
Additionally, eight pots were taken at random of each level of nitrogen to measure undisturbed height, tiller density, and leaf appearance rate and to evaluate the effect of N fertilization on these variables.
Sample measurements and analysis
After the uniform defoliation, the measurements of leaf appearance, undisturbed height, and tiller density were initiated. Undisturbed height was measured at ground level in five points of each pot, and the total tillers of each pot were counted. A tiller from each pot was marked, and measurements for leaf appearance rate were made each four days until the pot was harvest. The leaf appearance rate was determined, expressed as the degree days necessary for the appearance of a leaf (phyllochron) using equation 1.
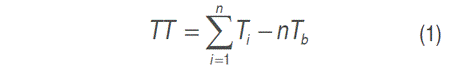
Where:
TT: Thermal time for the appearance of a leaf,
Ti: Mean daily temperature (°C) per day i,
Tb: Base temperature (°C) -this parameter was set at 8 °C, according to Ivory and Whiteman (1978).
Weekly, five tillers per pot were taken at random to count the number of leaves completely expanded by stolon. When the average of all pots within a treatment reached the number of leaves corresponding to their treatment, they were harvested, and the leaf, stem, and DF fractions were separated to perform the corresponding analyzes. Samples of the leaf, stem and dead material fractions were dried at 65 °C for 48 hours and then ground. Contents of CP, NDF, ADF, EE, ash, and lignin were estimated by Near-infrared spectroscopy (NIRS), according to the equations generated for Kikuyu grass by Ariza-Nieto et al. (2018).
Statistical analysis
Three different experimental models were used. A completely randomized model with factorial arrangement 4×4 (leaves per tiller × nitrogen fertilization) (Martínez et al., 2011) was used for the DM production from leaves, stems, GF:DF, stem-leaf ratio and for the nutritional composition of leaves and stems. A repeated measurement design (Martínez et al., 2011) was used for the analysis of the variable undisturbed height to determine the effect of nitrogen fertilization on the density of tillers and phyllochron. Homogeneity of the experimental material and normal distribution of the experimental error, with mean zero and common variance, were examined. Some variables were transformed to comply with the homogeneity of variances. The lack of normality was accepted when the data achieved the assumption of homogeneity of variances with the Levene test, which is considered robust to the lack of normality. The comparison of means for all the variables was made by the Tukey test. Hotelling test was used for the undisturbed height. The data were analyzed with InfoStat software, 2016I version.
RESULTS AND DISCUSSION
Dry matter yield
The interaction between numbers of leaves per tiller and nitrogen fertilization was not significant for DM yield of leaves per tiller (P=0.0751), stems (P=0.2616), GF (P=0.0790), L:S (P=0.7331), and GF:DF (P=0.8976). The interaction was only significant for DF (P=0.0451). DM yield of leaves and green forage increased when Kikuyu pasture was harvested with more leaves per tiller and/or fertilized with nitrogen. However, differences among harvest time (leaves per tiller) were small and only evident between L3 and L6. N fertilization differences were more evident at 50 kg N ha-1 year-1 (Figures 1 and 2). The highest leaf yield was reached at 150 kg N ha-1 year-1 (Figure 2). Stem yield increased with nitrogen fertilization, but the effect of leaves per tiller was not evident (Figures 1 and 2). Dead forage increased when the Kikuyu was harvested with more than 5 leaves per tiller. The magnitude of the response varied due to N fertilization (Interaction, P<0.0451), where differences among the leaves per tiller treatment were not evident for the treatment without N (Figure 3).
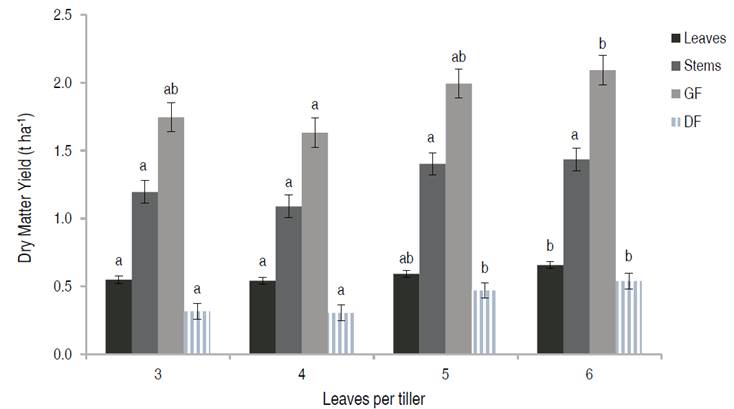
Figure 1 Dry Matter yield of leaves, stems, green forage (GF), and dead forage (DF) of kikuyo defoliated at a different number of leaves per tiller. Different letters within forage component (leaf, stem, GF, DF) mean significant difference (P<0.05).

Figure 2 Dry Matter yield of leaves, stems, green forage (GF), and dead forage (DF) of kikuyu fertilized with differen doses of nitrogen. Different letters within forage component (leaf, stem, GF, DF) mean significant difference (P<0.05).

Figure 3 Yield of dead forage (DF) in kikuyu according to nitrogen fertilization and defoliation at a different number of leaves per tiller. Different letters within the number of leaves per tiller mean significant difference (P<0.05).
Previous researches recommended harvesting Kikuyu at a maximum leaf stage of 4.5 since the L:S begins to decrease after 4 leaves per tiller (Fulkerson et al., 1999; Reeves and Fulkerson, 1996). In this experiment, the L:S remained constant regardless of the number of leaves per tiller, and it was not infIuenced by nitrogen fertilization (Figures 4 and 5). This result is similar to Fonseca's et al. (2016) study, which suggests that the changes in the proportion of stems in Kikuyu grasslands for the 4 to 6 leaf stages do not occur under the high altitude conditions in the tropics, maintaining a correlation between the number of leaves and stems within the pasture, given its stoloniferous growth habit. Fulkerson et al. (1999) found that the proportion of stems increased among leaf stages, with proportions of 9.5%, 13.1%, and 20% for 2, 4, and 6 leaves, respectively. However, this response varies by season. For summer, there were no differences in the proportion of stems among pasture harvested at different numbers of leaves per tiller. In autumn-winter, when the weather conditions are similar to those of the high altitude tropics of Colombia, L:S was similar between pasture harvested at 4 and 6 leaves per tiller and only differed to the grass harvested at 2 leaves per tiller.

Figure 4 Leaf-stem ratio (L:S) and green forage-dead forage ratio (GF:DF) according to the kikuyu defoliation at a different number of leaves per tiller. Different letters within a ratio mean significant difference (P<0.05).
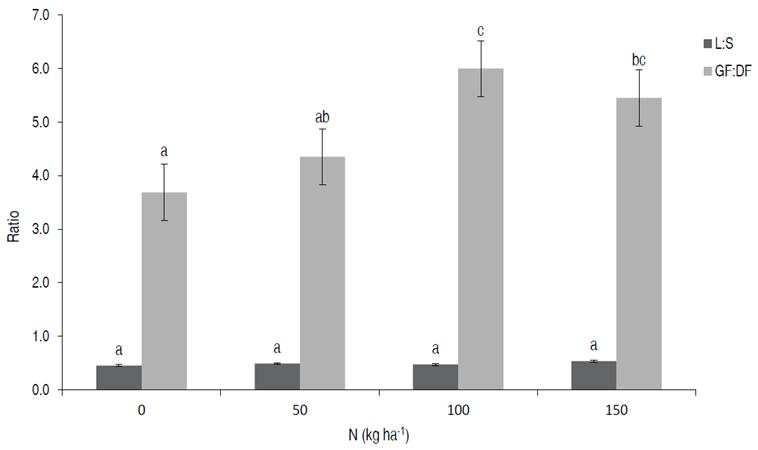
Figure 5 Effect of the nitrogen fertilization on leaf: stem ratio (L:S) and green forage-dead forage ratio (GF:DF). Different letters within a ratio mean significant difference (P<0.05).
In the present study, L:S was close to 0.5, similar to previous studies (Ivory and Whiteman, 1978), in which the whole plant was harvested, including the stolons at ground level, which may have generated a greater production of DM from stems than from leaves. The GF:DF was reduced when Kikuyu was harvested with more leaves per tiller (Figure 4) but increased by N fertilization (Figure 5). Changes in this ratio were insignificant between leaves 3 and 4 in a tiller that had similar ratios, but the changes were accelerated after 4 leaves (Figure 4). These results are comparable to the study reported by Reeves and Fulkerson (1996), who found the proportion of DF increased substantially after 4 leaves per tiller.
Different authors have reported the positive response of Kikuyu to nitrogen fertilization regarding yield and quality (García et al., 2014; Mejía-Taborda et al., 2014; Dugmore, 2011; Castillo et al., 1983). In this research, N fertilization resulted in a higher yield of DM of leaves, stems, and GF, reaching the highest production with N150 (Figure 2) compared to analogous studies. Although nitrogen fertilization also increased the yield of dead forage, N improved the GF:DF ratio, suggesting a positive net effect of N on pasture quality. The best results were obtained with N100 and N150 (Figure 5). Castillo et al. (1983) found a favorable response in the growth rate of Kikuyu with levels of up to 500 kg N ha-1 year-1, but with a lower economic efficiency because of the increase in the quantity of N applied. So the optimal level of application is a function of profitability, and it is not related to the maximum response of the species.
Herrero et al. (2000) suggested the best harvest time (leaves per tiller) for Kikuyu will depend on N fertilization. In the present study, the effects on yield and chemical composition of harvesting Kikuyu at different leaves per tiller were independent of the effects of N fertilization. Moreover, when Kikuyu was not fertilized or even with low levels of fertilization (N50), it was not possible to reach the recommended biomass to grazing (2,600 kg DM ha-1) (García et al., 2014). Therefore, the leaf stage is not a unique criterion for harvesting time. It should be considered the undisturbed height and pre-grazing cover to define the optimum grazing time. Finally, N fertilization could reduce pasture rest period (lower leaf stages), to reach higher annual forage yields of better quality.
Chemical composition
The interaction between numbers of leaves per tiller and nitrogen fertilization was not significant for CP (P=0.4052), NDF (P=0.9148), ADF (P =0.9462), lignin (P=0.6827), EE (P=0.4868) and ashes (P=0.1000).
By increasing the number of leaves per tiller at harvest, the level of crude protein in both leaves and stems was slightly reduced. This difference was only significant between the Kikuyu harvested at L3 and L6 (Table 2). NDF content in leaves was higher as tillers had more leaves, but not on stems (Table 2). In the leaves, the ADF and the ash were not modified regarding the increase in the number of leaves per tiller at harvest. However, in the stems, the contents of ADF and ash were reduced, being lower at L6 (Table 2).
Table 2 Chemical composition of Kikuyu leaves and stems defoliated at a different number of leaves per tiller.
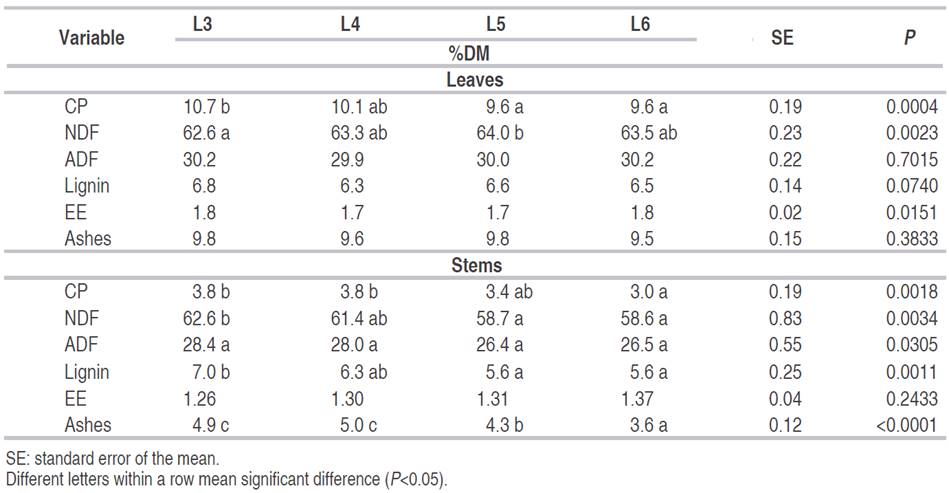
Quality of kikuyo is already changing at L5, reducing the content of CP and increasing the content of NDF with respect to L3. However, no significant differences were found between L4, L5, and L6, similar to what was found by Fonseca et al. (2016) and contrary to the study reported by Reeves and Fulkerson (1996). The content of NDF and lignin of L5 and L6 respect to L3 and L4 were inferior. This could be explained by the appearance of a new secondary tiller in L5 and L6, which have lower cell wall content, while in L3 and L4, primary tillers were dominant.
The results of this experiment show that the best quality of the Kikuyu is found in L3. Yet, at this stage, it may be challenging to reach enough biomass for grazing. So to harvest the pasture may be recommended between L4 and L5 stages. Although in the L6 stage, there were no significant differences in nutritional quality with respect to L4 and L5, senescence losses began in the pasture, with a negative impact on the efficiency of use. Escobar et al. (2020) reported the highest nutrient yields (CP, Net Energy for Lactation [NEL]) for L5 in a ten-month on-farm trial without N fertilization. Harvesting Kikuyu with fewer leaves per tiller (<5) may be feasible if nitrogen fertilization is applied or cultivated in high fertile soils (García et al., 2014; Dugmore, 2011; Mejía-Taborda et al., 2014; Castillo et al., 1983).
Nitrogen fertilization had a positive impact on the chemical composition of Kikuyu. Similar results were found by Dugmore (2011) and Castillo et al. (1983). The content of CP in leaves increased by N fertilization, while the content of NDF in the leaves and stems decreased (Table 3). The high content of NDF in Kikuyu is considered limiting in terms of nutritional quality, which can restrict its intake. (García et al., 2014; Correa et al., 2008). The reduction in NDF by nitrogen fertilization was reached with levels from 0 to 500 kg ha-1 year-1, regardless of the source of nitrogen used, achieving 53.5% as the minimum content of NDF (Castillo et al., 1983). Therefore, it is possible to reduce the NDF content by nitrogen fertilization, within the ranges of the species and counteract the negative effect on DM intake.
Table 3 Chemical composition of Kikuyu leaves and stems with different levels of nitrogen fertilization.

The values obtained for CP and NDF differ significantly from those found in field conditions for Kikuyu. In general, CP values were lower, and NDF values were higher than those (17.8±0.9% and 58.3±1.5%, respectively) reported by García et al. (2014). It could be due to the characteristics of the soil used, which was highly restrictive in the supply of nitrogen (Yutaro and Osamu, 2018). The high organic matter content in soils from bleak uplands has been related to low mineralization due to the low temperatures present at high altitudes that limit biological activity (IDEAM, 1999). These characteristics of the soil led to the N0 treatment being very limiting in nitrogen for the plant, an aspect refIected in the poor growth obtained and in difficulty for the establishment of the Kikuyu in the pots of this treatment.
Structural variables and phyllochron
The undisturbed pasture height and density of Kikuyu tillers were increased by N fertilization, while the phyllochron was reduced. The effects of N on tiller density was only evident for the highest level of N fertilization (Table 4). The increase of N fertilization has been related to a higher foliar elongation (Borrajo and Alonso, 2014; Da Silva et al., 2012; Almeida et al., 2011). Therefore, nitrogen deficiency may be related to the slow elongation of Kikuyu pastures observed in low fertility soil used in this experiment, and greater height of the pasture in response to N. Previous studies also had reported N fertilization has a positive effect on tilling (Dourado et al., 2015, Da Silva et al., 2012). This effect may be greater in plants that grow distant from each other than in dense stoloniferous growth species such as Kikuyu. These pastures are usually denser, and the light competition may limit the increments in tiller density (Lemaire et al., 2009).
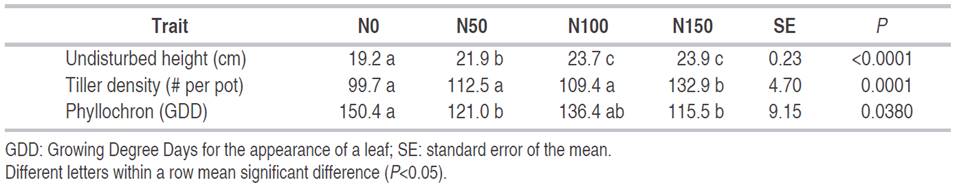
The leaf appearance rate is primarily infIuenced by temperature and then by soil moisture and nutrient content (Wilhelm and McMaster, 1995). It was observed in this experiment, without N fertilization (N0) since Kikuyu had a higher phyllochron while in the other N fertilization levels were similar (Table 4). Wilhelm and McMaster (1995) concluded that under extreme conditions of soil fertility, like N0 treatment, the phyllochron may be increased due to a longer time required for the appearance of a leaf. Therefore, it limits the accumulation of DM.
CONCLUSIONS
Harvesting Kikuyu with higher leaves per tiller (>5) or fertilized with N increased DM yield. However, the best nutritional quality was obtained harvesting with fewer leaves (<4), and fertilizing with higher concentrations of N. DM yield was 1.7 t ha-1 harvesting at a lower number of leaves per tiller, which is not suitable to grazing regardless of the positive effect of N fertilization. It could be recommended to harvest the Kikuyu between 4 and 5 leaves per tiller to maintain the balance between quantity and quality. Harvesting Kikuyu, with fewer leaves per tiller than 4, would be feasible if pastures are fertilized with higher levels of N (>100 kg ha-1). Regardless of the number of leaves per tiller, fertilization can maximize DM production and contributes to the improvement of nutritional quality by increasing the contents of CP and decreasing NDF contents in leaves and stems. Likewise, it improves GF:DF, the production of DM such as leaves, and the undisturbed height of the pasture. Therefore, N fertilization is a key element to improve DM yield, the nutritional quality of the pasture, and to increase defoliation frequency. Additionally, this study suggests that the number of leaves per tiller cannot be used as the only criteria to define the moment to graze. Also, pre-grazing biomass available and the undisturbed height of the pasture should be considered.

