











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Sourgrass (Digitaria insularis [L.] Fedde), a Poaceae perennial weed, is infesting large areas across South America. Its dissemination is occurring because of the plant characteristic, such as tufted formation, rhizome structures, high seed production, and because of the pressure selection of plant biotypes with resistance to glyphosate and ACCase herbicides (Machado et al., 2008; Veldman and Putz, 2011; Melo et al., 2012; Gemelli et al., 2013; Gazola et al., 2019). All these characteristics avoid the easy control of this plant, genarating interference in crops. For instance, the coexistence of six plants m-2 of D. insularis with soybean crop is enough to reduce its yield in 40% (Gazziero et al., 2019).
The glyphosate has been used as the main herbicide in weeds management for many years, but its intensified use in weed pre-sowing control with non-tillage system and post-emergence generates glyphosate-tolerant transgenic crops (Green, 2018). D. insularis has a great vegetative propagation and a high seed production in a short time with seed germination across the year. Therefore, it is required to increase the rates of herbicides for effective control.
This grass presents cases of resistance to herbicides in Brazil, with resistance to glyphosate (Adegas et al., 2010; Carvalho et al., 2011) and ACCase inhibitors (haloxyfop and pinoxaden) (Takano et al., 2020).
There are many ways to manage the D. insularis resistant to glyphosate, including the herbicide application in the early stage of development, avoiding seed production, rotation of herbicides with different mechanisms of action or chemical groups, among other cultural practices including the burndown. It could be important for the effective management of D. insularis; its use must be implemented in advance of sowing and as complementary application to other products (Oliveira-Júnior et al., 2006; Canedo et al., 2019). The burndown immediately before sowing involves the application of one or more herbicides (usually systemic action), its choosing depends on the floristic composition of the site and infestation density (Oliveira-Júnior et al., 2006; Frisvold et al., 2020).
Particularly for D. insularis, there are few herbicides for chemical control. Paraquat, for example, with a single application is not enough to eradicate the whole plant causing re-growth (Zobiole et al., 2016). Besides, paraquat is being taken off the market in Brazil in September 2020 (ANVISA, 2020). Diquat is neither a great alternative in the control of grasses, generally with low efficacy improving when is associated with adjuvants; however still being unsatisfactory option to control it (Gitsopoulos et al., 2014). On the other hand, the use of ACCase herbicides, especially "fop" herbicides could lead to a rapid herbicide-resistant biotypes selection (Takano et al., 2020).
The use of herbicides of different mechanisms of action and with the same control spectrum is one strategy that must be used. Because of the presence of other weeds in the field, with infestation of grasses and broad-leaved, it is common the spray of glyphosate or ACCase mixed with synthetic herbicides. There are reports of the antagonist effect of 2,4-D on the action of ACCase inhibitors graminicides (Gomes et al., 2020), due to the reduction of translocation and increase of herbicides metabolism from the ariloxifenoxipropionics group (Trezzi et al., 2007). Pereira et al. (2018) observed that synthetic auxinic (2,4-D and dicamba) associated with haloxyfop interfered negatively on D. insularis control.
There is still contrasting information that considers the antagonist action of 2,4-D and other synthetic auxinic herbicides (triclopyr and dicamba) mixtures with glyphosate and ACCase inhibitors. As D. insularis has few options of herbicides for chemical control, this study aimed to evaluate the control of D. insularis, with glyphosate plus ACCase inhibitors and latifolicides applied in pre-emergence and verify possible antagonism between ACCase inhibitors herbicides with synthetic auxins and other latifolicides.
MATERIALS AND METHODS
Design and experimental conditions
Two experiments were installed in a commercial area in 2018-2019 season. Palotina, Paraná (PR), Brazil, (24°2326.93"S 53°84'51.36"W) and Corpus Christi, Canindeyú, Paraguay (24°3'37.24"S 55°0'22.22"W) were the locations selected. The climate of both regions is classified as Cfa (humid subtropical with abundant rainfall, well distributed throughout the year), according to Köppen classification. Figure 1 (A,B) presents climate data during the period of experiment conduction.
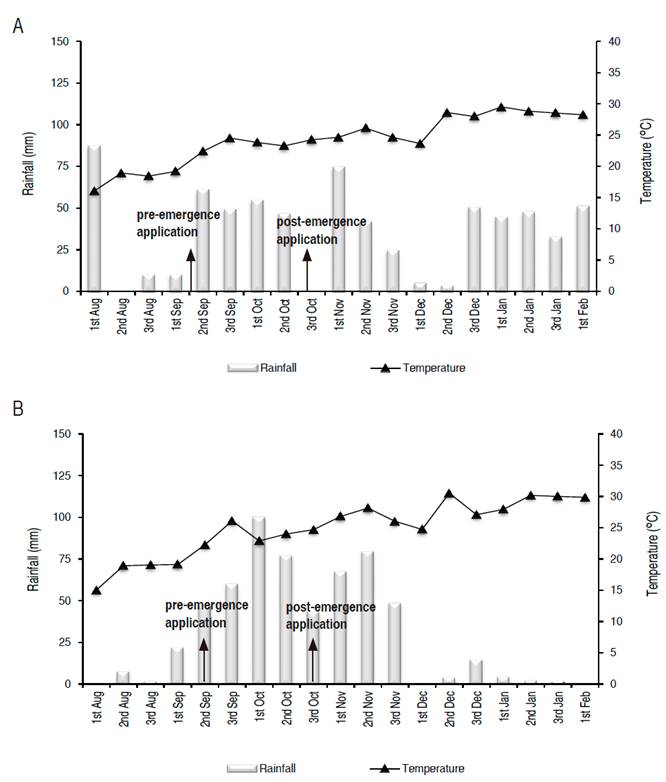
Figure 1 Rainfall representation average temperature for the experiment site. Aug - Nov 2018. A. Palotina, PR, Brazil. B. Corpus Christi, Canindeyú, Paraguay.
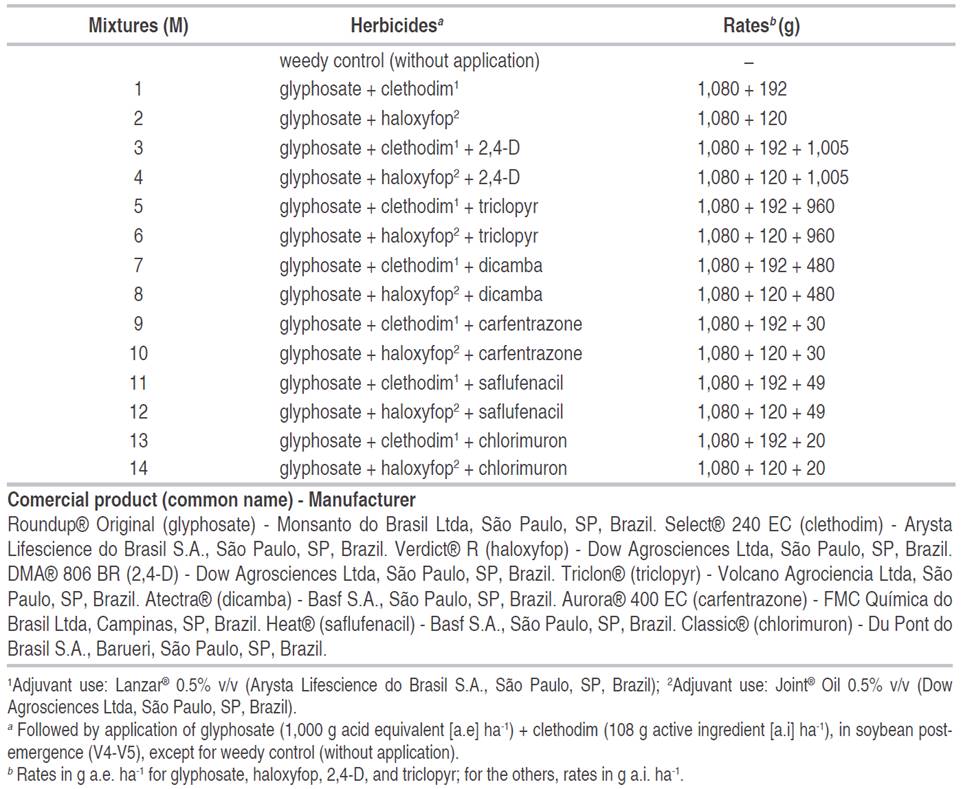
Both areas had a high population of perennial sourgrass at flowering stage, with records of use of glyphosate, and its loss of efficacy in recent years. Palotina assay had a populational density average of 1 to 2 tufts m-2, meanwhile, in the Paraguay area, the populational density was higher, 2 to 4 tufts m-2. The experiments were installed in a fallow field, prior to soybean sowing. Previously, the Palotina area had been cultivated with maize crop, and Corpus Christi was a fallow area since the soybean harvest in summer (without second season crop). A randomized block with four replications was used in the experimental design. The experimental plots were composed of 3x5 m-2. The treatments are described in Table 1.
The treatment applications were performed two days before soybean sowing. In Palotina the application occurred on September 11, 2018, at 29 °C, relative humidity of 60%, and wind speed of 6.5 km h-1. While in Corpus Christi on September 15, 2018, the temperature was 23.9 °C, relative humidity 72.3%, and wind speed 6.8 km h-1. The application of M1 in soybean post-emergence was performed 42 days after emergence (DAE) in both locations, with soybean plants at the V4-V5 stage (BBCH, 2001). This application was carried out in all treatments, except in the weedy control (without any application). In Palotina the application occurred on Octuber 30, 2018, at temperature of 30 °C, relative humidity of 58%, and wind speed of 5.1 km h-1. In Corpus Christi on November 03, 2018, the temperature was 26.9 °C, relative humidity 78%, and wind speed 6 km h-1. CO2-pressurized sprayer was used for all herbicide applications. It was equipped with six AIXR 110 015 spray nozzles spaced 0.5 m from each other, 2.5 kgf cm−2 calibrated pressure and speed of 1 m s-1, resulting in a spray volume of 150 L ha-1.
Evaluations and statistical analysis
At soybean pre-emergence, the visual evaluations of control were done at 7, 14, 21, 28, and 35 days after application (DAA) for both experiments. The control also was evaluated at 7, 14, and 21 DAA, in Palotina, and at 7 and 14 DAA in Corpus Christi, at soybean post-emergence. Percentage values from 0 up to 100% were assigned for the evaluation (0 no injuries, 100% plant death) with regard to weedy control (Velini et al., 1995).
The data were submitted to analysis of variance (ANOVA) by F-test (P<0.05), according to Pimentel-Gomes and Garcia (2002). The means of treatments were grouped by Scott and Knott (1974) test (P<0.05). Sisvar 5.6 software was used for the analysis (Ferreira, 2011).
RESULTS AND DISCUSSION
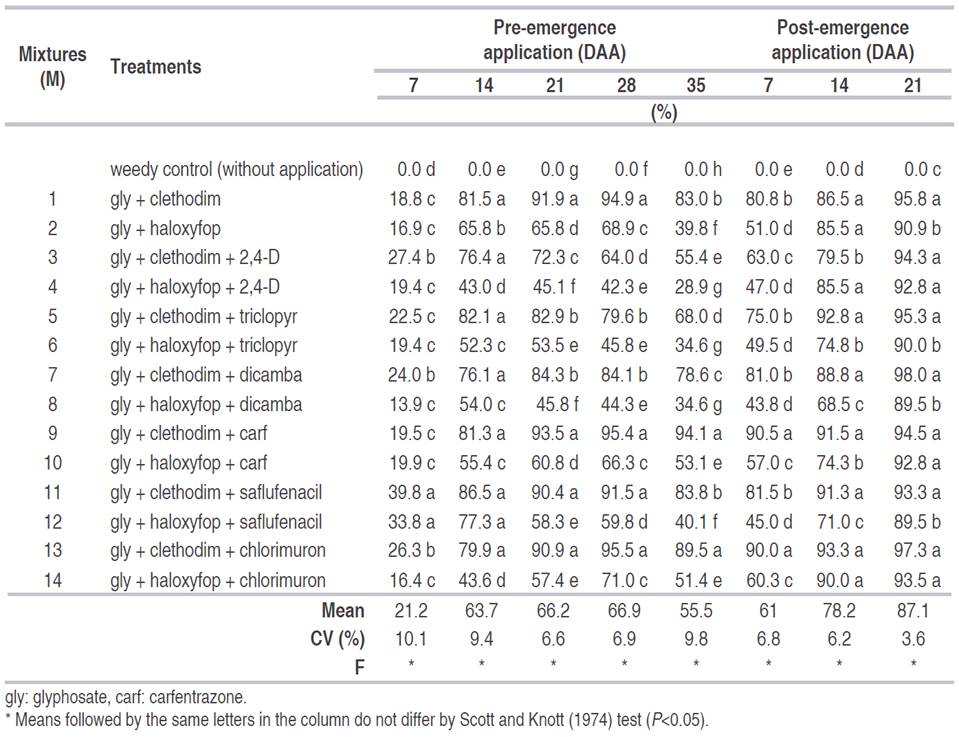
All treatments with clethodim were more effective compared with haloxyfop treatments from 21 to 35 DAA (Palotina). At 7 DAA, there were not high levels of control, at most 39.8% (Table 2). On the other hand, the treatments with saflufenacil provided greater control, even 39.75% higher than other treatments. For 35 DAA, stands out that the best results were observed for treatments M13 or M9, with values up to 94.1%. After the post-emergence application of M1, it was observed some differences between the treatments, there were lower values for some treatments with haloxyfop application, but all mixtures provided minimum control of 89.5%, at 21 DAA (Table 2).
Table 2 Control treatments to D. insularis at 7, 14, 21, 28, and 35 days after the application (DAA) of herbicides, at soybean pre-emergence, and at 7, 14, and 21 DAA at soybean post-emergence. 2018-2019 season, Palotina, PR, Brazil.
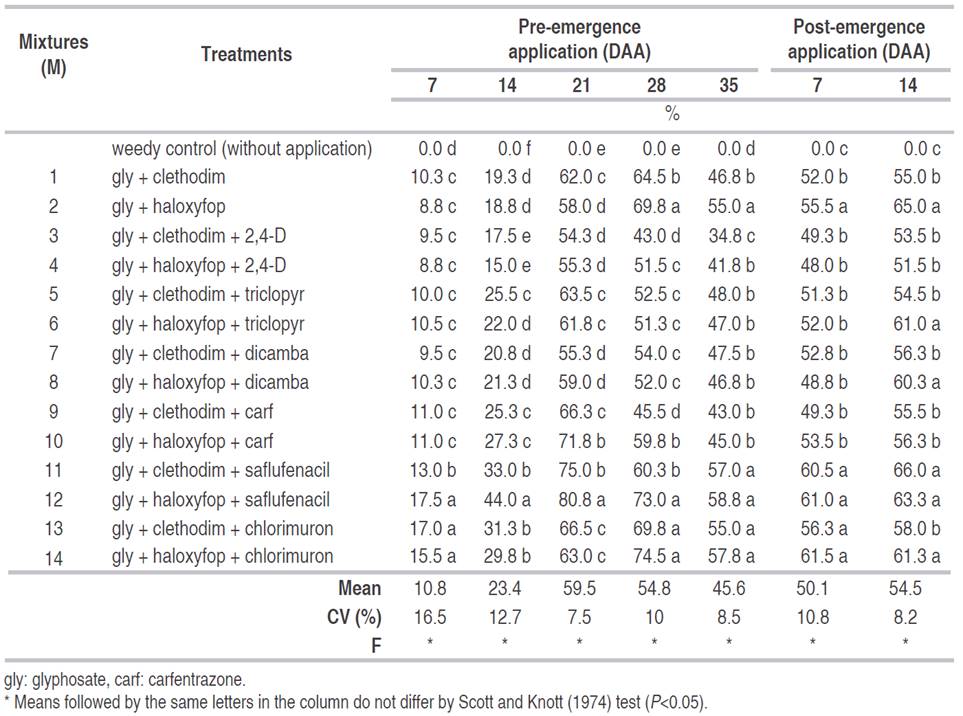
In Corpus Christi, the control was lower in general, probably due to the higher infestation of perennial plants. The higher averages on 28 DAA were reached by the treatment's M2, M12, M13, and M14 up 74.5%. Even after the post-emergence application at 14 DAA using M11, a D. insularis control up 66% was observed (Table 3).
Table 3 Control treatments to D. insularis at 7, 14, 21, 28, and 35 days after the application (DAA) of herbicides, at soybean pre-emergence, and at 7, 14, and 21 DAA at soybean post-emergence. 2018-2019 season, Corpus Christi, Canindeyú, Paraguay.
M11 was effective in the control, especially in the first evaluations in Palotina. The application of glyphosate+saflufenacil has been reported effective especially to control eudicotyledon weeds, for example, Conyza spp. (Mahoney et al., 2016). However, saflufenacil does not present high control on Digitaria spp. (Soltani et al., 2014). Nevertheless, in this study, in Palotina, it could be an adjuvant on D. insularis control, when was mixed with clethodim (M11) or haloxyfop (M12). The addition of saflufenacil in the mixtures did not increase the control promoted by glyphosate+clethodim; however it did not have an antagonistic effect. Mixtures like this have a broad spectrum of action, being important in weed management in infested areas with D. insularis and other weeds (Roskamp et al., 2012).
In Palotina, all treatments with clethodim were more effective compared with haloxyfop treatments from 21 to 35 DAA. Zobiole et al. (2016) did not observe differences on D. insularis control between clethodim and haloxyfop mixed with glyphosate. Cassol et al. (2019) found similar efficacy of clethodim and haloxyfop in association with glyphosate on perennial D. insularis control and at soybean post-emergence weed control, but in the control of plants in the off-season, clethodim+glyphosate was more powerful than haloxyfop+glyphosate.
Other studies highlight the efficacy of clethodim and haloxyfop, at different mixtures, on D. insularis control (Barroso et al., 2014; Gilo et al., 2016). Nonetheless, it is not possible to determine which one is more effective on D. insularis control; their choice should consider several factors including the background of the use of these herbicides in the area. According to López-Ovejero et al. (2017), Takano et al. (2018), and Lucio et al. (2019) it is crucial to rotate the chemical groups in order to avoid herbicide-resistant D. insularis biotypes.
A reduction in weed control was found in both locations from 14 DAA regarding the synthetic auxins in association with ACCase inhibitors. In Palotina, at 28 DAA, 30.93, 15.30, and 10.85% of control losses were observed when 2,4-D (M3), triclopyr (M5), and dicamba (M7) were mixture with glyphosate+clethodim, respectively. In Corpus Christi at 28 DAA, the losses of effectiveness were 21.5, 17.25, and 10.5% for treatments with 2,4-D (M3), triclopyr (M5), and dicamba (M7) respectively, when compared with glyphosate+clethodim treatment. In relation to glyphosate+haloxyfop associations, the auxins 2,4-D, triclopyr and dicamba provided a reduction of 18.25, 18.5, and 17.75% in control effectiveness. At the same time, the treatments with auxinics added to haloxyfop provided 42.25% (M3), 45.25% (M6), and 45.75% (M8) of weed control.
In this study, antagonism and reductions were verified by the association of ACCase inhibitors, as haloxyfop with 2,4-D (M4) and dicamba (M8). By Pereira et al. (2018), some cases more than 40% was observed for both. Clethodim+dicamba is also reported as an antagonist, with losses of 6 to 15%, in volunteer maize control in soybean (Underwood et al., 2016). The losses due to antagonism were of 11.85% in Palotina and 10.5% in Corpus Christi treated with M7.
In the case of haloxyfop, the antagonism on these mixtures is possibly explained by the reduction of translocation.(Olson and Nalewaja, 1981). The clethodim+2,4-D antagonism was detected due to leaf necrosis in a few days after application (Gomes et al., 2020). This can result in less absorption and translocation triggering the less weed control. Mixtures of triclopyr with ACCase inhibitors also reduce its effectiveness on grasses control (Scherder et al., 2005). About dicamba, it can be also explained for the possible reduction of translocation of the graminicide to the roots and the plant's rhizome (Aguero-Alvarado et al., 1991).
The D. insularis control varied between the two locations, which could be explained by higher population density in Corpus Christi. While the lower efficacy of haloxyfop treatments (compared to clethodim) in Palotina, probably it is a risk warning for the selection of haloxyfop resistant biotypes. There are records of the D. insularis resistence to this herbicide and pinoxaden in Brazil (Takano et al., 2020).
In a general context, the D. insularis control was satisfactory for treatments with triple combinations, only in the Palotina area. There, even with ineffective control for treatments composed with synthetic auxins, the post-emergence application in soybean increased the control level, with satisfactory final controls for all treatments. However, antagonism was observed for all synthetic auxins in both locations.
The coexistence of six plants m-2 of D. insularis with soybean crop is enough to reduce yield in 40% (Gazziero et al., 2019). In Corpus Christi with a population of 2 to 4 tufts per m2, the application was not effective. The anticipated control of D. insularis populations should be a priority to avoid crop losses due to weeds competition. The combination of ACCase inhibitors (haloxyfop, clethodim) and synthetic auxins (2,4-D, dicamba, and triclopyr) is not recommended. Another option is to use the auxin herbicides in sequential application. For instance, by Leal et al. (2020) haloxyfop must be applied at least 6 days before 2,4‐D to control Conyza spp. and D. insularis when they are present simultaneously.
CONCLUSION
In Palotina, the perennial D. insularis control at soybean pre-emergence burndown was effective in some mixtures that presented glyphosate+ACCase inhibitor added to carfentrazone, saflufenacil, or chlorimuron, demonstrating the potential use of these associations for weed control. Even with ineffective control for treatments composed with synthetic auxins, the post-emergence application in soybean increased the control level, with satisfactory final controls for all treatments
In Corpus Christi, the herbicides combinations were not effective, even after the post-emergence application of glyphosate+clethodim.
Clethodim and haloxyfop had a reduction on the efficiency in combination with the synthetic auxins 2,4-D, triclopyr, and dicamba. Among synthetic auxins, dicamba showed the lowest antagonism.

