











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Vegetative propagation is typically an efficient method of maintaining specific traits of plant genetic resources. Considerable amount of superior quality planting materials can be produced through clonal propagation within short time (Baul et al., 2010; Hartmann et al., 2011; Wetzstein et al., 2018). Adventitious rooting, as one of the most important methods of plant vegetative propagation, is a requirement for successful production of practical clones. It is also one of the most efficient methods for fast commercial production of horticultural plants worldwide (Shi-Weng et al., 2009b). Hardwood cuttings are one of the least expensive and easiest methods of vegetative propagation. They are not easily perishable, may be shipped safely over long distances and require little or no special equipment during rooting (Hartmann et al., 2011).
The GF677 (Prunus amygdalus × Prunus persica) is a valuable rootstock, which has a wide range of compatibility with various cultivars and species (Karimi and Yadollahi, 2012; Gainza et al., 2015). It is tolerant against donor Fe deficiency and suitable for semi-drought conditions, calcareous and poor fertility soils (Legua et al., 2012; Bagheri et al., 2016; Ranjbar et al., 2019). Therefore, to find the appropriate methods to improve the GF677 propagation is highly important (Tsipouridis and Thomidis, 2004; Tsipouridis et al., 2005; Karimi and Yadollahi, 2012; Sarikhani et al., 2017).
Environmental and endogenous factors such as temperature, light conditions, plant growth regulators (PGRs), carbohydrate, mineral elements and other molecules, may act as signals and induce groups of cells to alter the result in adventitious rooting (Shi-Weng et al., 2009a; Hartmann et al., 2011). In addition to the genotype of the plant, the nutritional status, the phenological stage, the environmental factors, and the climatic conditions cause seasonal variations in the rooting capability of the woody cuttings (Hartmann et al., 2011). Application of plant growth regulators (PGRs) is one of the most effective procedures to increase root initiation, rooting percentages, and quality and uniformity of roots. The most widely used PGRs for this purpose is indole butyric acid (IBA) (Ozelbaykal and Gezerel, 2005; Asl moshtaghi and Shahsavar, 2011; Nazary and Yadollahi, 2012; Nag et al., 2013; Caplan et al., 2018). Furthermore, it has been reported that polyamines (PAs) are able to promote root formation and root development in difficult-to-root plants; particularly, Put has displayed a better response in comparison with other PAs (Rey et al., 1994; Wu et al., 2010; Sivanandhan et al., 2011; Silvestri et al., 2018). Rey et al. (1994) found a powerful enhancing effect of PAs on microshoots rooting of hazelnut. PAs improved rooting when microshoots were treated with IBA in a synergistic mode; probably generated a better induction of roots, while PAs had only a limited beneficial impact on rooting when applied without IBA. Additionally, Put in combination with IBA promoted early rooting and increased the rooting percentage in hazelnut (Cristofori et al., 2010), olive (Rugini et al., 2016) and in Ficus (Ghasemi and Kosh-Khui, 2019). Some studies demonstrated a possible improvement of the auxin stimulation on adventitious root formation when it is mixed with co-factors such as phenolics (Bartolini and Tattini, 1986; De-Klerk et al., 1999), flavonoids (Lewis et al., 2011) and hydrogen peroxide (H2O2) (Sebastiani and Tognetti, 2004; Shi-Weng et al., 2009a and b). Moreover, there is a connection between PAs effects on rooting enhancement of cuttings and increase of peroxidase activity at the basal end of cuttings (Rugini et al., 1997).
The relationship between tissue carbohydrate contents and rooting has remained controversial for many years (Tsipouridis and Thomidis, 2004). Most of the tree species can be rooted with leafy cuttings rather than leafless cuttings (Ky-Dembele et al., 2011). Indeed, further carbohydrate substances, seasons and cutting date are critically important elements. For instance, some cuttings as quince can root at any season of the year, whereas cuttings obtained from another species such as cherry and olive root are only successful at a certain time of the year (Hartmann and Loreti, 1965; Hartmann et al., 2011). Ucler and Parlak (2004) examined the influence of IBA and cutting date on rooting of semi-hardwood cuttings of kiwifruit and reported that the cuttings which were taken in August had better rooting compared to those taken in July. In addition, they concluded that cutting date had a significant influence on rooting potential.
In this context, the aim of this research was to evaluate the effects of IBA, Put and H2O2 in different concentrations and seasons, separately or in combination with each other, on rooting of GF677 rootstock.
MATERIALS AND METHODS
Location and plant material
This experiment was conducted in the research greenhouse of Bu-Ali Sina University, Hamedan, Iran (1741.5 masl, 34°47'N, 48°30'E), as a completely randomized design with three replications and 12 cuttings in each replication. Uniform cuttings of GF677 (Prunus amygdalus × Prunus persica) were collected from Paradise Nursery in Hamedan province and Sanaz Nursery in Zanjan province. At first, the semi-hardwood cuttings with about 30 cm length were immersed in Put or H2O2 solutions for 30 s, and then treated with IBA for 10 s. Subsequently, cuttings were planted diagonally in a medium involving sand and perlite with a ratio of 80 and 20%, respectively. Furthermore, the rooting bed was covered with covering film to maintain the moisture content. The research included three experiments as following:
First experiment
The first experiment was done on July 6th, and semi-hardwood leafy cuttings were taken from the middle third of the current season branches of the 3-year-old GF677 donor plants. The cuttings were treated with Put and H2O2 in seven levels including distilled water (control), Put at three concentrations of 800, 1600 and 3200 mg L-1 and H2O2 at three concentrations of 1.5, 3 and 6 % w/v, as one factor. Another factor was IBA in four concentrations of 0, 1000, 2000 and 3000 mg L-1.
Second experiment
The experiment was prepared on March 6th and hardwood cuttings without leaf were taken from the current season branches of the GF677 donor plants. Similar to the first experiment, the cuttings were treated with IBA, Put and H2O2.
Third experiment
This experiment was performed on October 16th and semi-hardwood leafy cuttings were taken from the current season branches of the GF677 donor plants. Based on results of first and second experiments, the desired levels of IBA (0, 1000 and 2000 mg L-1), Put (800 and 1600 mg L-1) and H2O2 (1.5 and 3 % w/v) were used at this stage.
In all three experiments, cuttings were taken in early morning. Immediately after treatment, the cuttings were cultured on the medium mentioned previously. The moisture content was monitored daily. After three months, cuttings were removed from the culture medium and some traits were analyzed such as callus proliferation, rooting of cuttings, fresh and dry weight of rooted and/or callused cuttings.
Statistical analysis
All data were normalized using arc-sin transformation method, and later statistically analyzed based on full factorial experiment in a completely randomized design by using the SAS software (version 9.1). Analysis of variance and Duncan's multiple-range tests (P≤0.05) were performed to assess possible significant differences among treatments.
RESULTS AND DISCUSSION
First experiment
The analysis of variances (P≤0.01) showed significant effects of IBA, Put and H2O2 and their interactions on callus proliferation percentage (Data of analysis of variance not shown). The callus proliferation was from 0.00 to 42.33% of treated cuttings taken in July. The highest percentage of callus proliferation (42.33%) was observed in cuttings treated with IBA 1000 mg L-1, which showed no significant differences with some other treatments. However, the lowest percentage (0.00%) corresponded to Put treated cuttings. Furthermore, no callus proliferations were noticed using 3000 mg L-1 IBA+1600 mg L-1 Put (Table 1).
Table 1 Effect of IBA, Put and H2O2 on callus proliferation and rooting of GF677 cuttings taken in July.
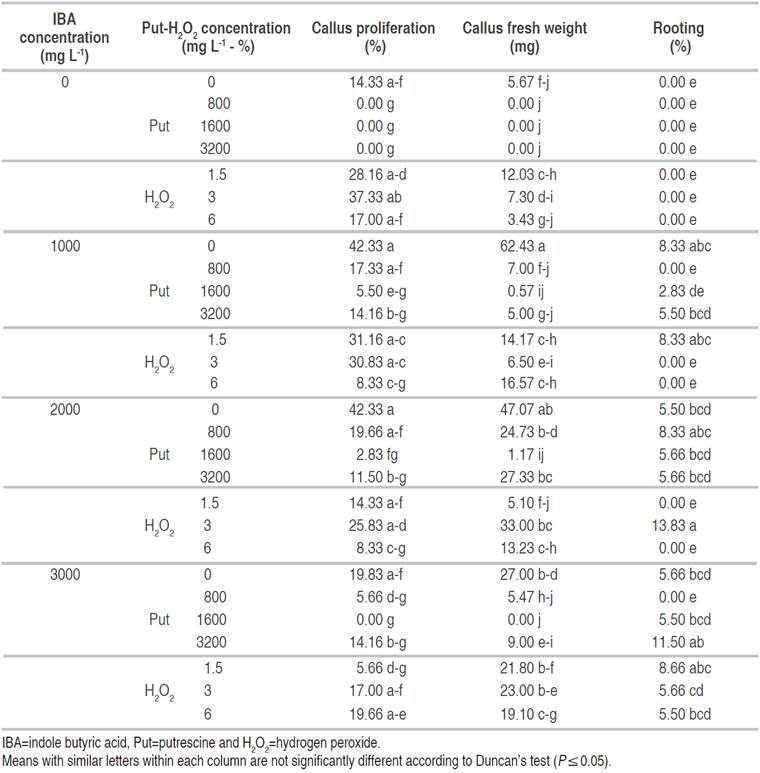
The effect of IBA, Put and H2O2 and their interactions on callus fresh weight were significant at P≤0.01. Application of IBA at 1000 mg L-1 led to the highest callus fresh weight in comparison with other treatments. The lowest callus fresh weight was obtained by Put at 800 mg L-1, 1600 mg L-1, 3200 mg L-1, and also 3000 mg L-1 IBA+1600 mg L-1 Put with no significant difference from control (Table 1).
Regarding rooting response, the application of IBA, Put and H2O2 and their interactions were significant (P≤0.01) on rooting percentage. Rooting percentage in this experiment was ranged between 0.00 to 13.83%. The highest rooting percentage (13.83%) was obtained by the application of 2000 mg L-1 IBA+ 3% H2O2. It did not have a significant difference when the treatments: 3000 mg L-1 IBA+3200 mg L-1 Put, 3000 mg L-1 IBA+1.5% H2O2 and 1000 mg L-1 IBA+1.5% H2O2 were used. The results of this study showed that the effect of cuttings preparation time as well as auxin, Put and H2O2 treatments were effective on callus formation and rooting of GF677 cuttings. Auxin has been proven to be an important factor in the rooting of many cuttings. Therefore, the internal auxin level is vital in callus production and rooting of cuttings in various plants. In the case of high levels of endogenous auxin, rooting can easily occur in many cases (Hartmann et al., 2011). In contrast, in cases where external auxin treatment is required, the use of IBA as a commercial auxin to enhance rooting of many plants has been demonstrated (Shiozaki et al., 2013). The lowest rooting percentage was gained in all treatments without IBA such as those of control and different concentrations of Put-H2O2 (Table 1).
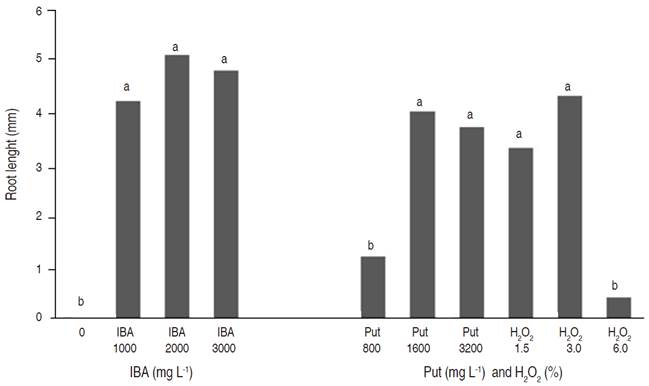
The effect of IBA treatment on root length (P≤0.05) was significant. The amount of Put at 1600 and 3200 mg L-1, and also H2O2 at 1.5 and 3% had no significant effects on root length in comparison with IBA in all concentrations. The highest root length (5.1 mm) was observed in cuttings treated with 2000 mg L-1 IBA with no significant difference between 1000 mg L-1 and 3000 mg L-1 IBA (Figure 1).
Second experiment
Analysis of variance showed significant effects of IBA, Put and H2O2 treatments and their interactions on percentage of callus proliferation (P≤0.01). The highest callus proliferation was obtained at 1000 mg L-1 IBA+1600 mg L-1 Put, with no significant difference at 1000 mg L-1 IBA+800 mg L-1 Put, 1000 mg L-1 IBA+3200 mg L-1 Put, 1000 mg L-1 IBA+ 3% H2O2, 2000 mg L-1 IBA+800 mg L-1 Put and 3000 mg L-1 IBA+800 mg L-1 Put. The callus proliferation percentages for control and IBA 2000 mg L-1 treatments were zero. Also, the IBA at 3000 mg L-1 had no significant difference with regard to control (Table 2). These results showed that rooting was gained after callus production. Callus formation of GF677 was coincided with data that was obtained from rooting percentage and number of roots. Researchers have suggested that basal callus formation generally tracks rooting of GF677 with young hardwood trees (Tsipouridis and Thomidis., 2004; Tsipouridis et al., 2005; Karimi and Yadollahi, 2012). Other authors have claimed that callus formation might help the rooting of some plant species with hardwood cuttings (Lodama et al., 2016; Zhou et al., 2018).
Table 2 The interaction effect of IBA, Put and H2O2 on callus proliferation and rooting of GF677 hardwood cuttings in March.
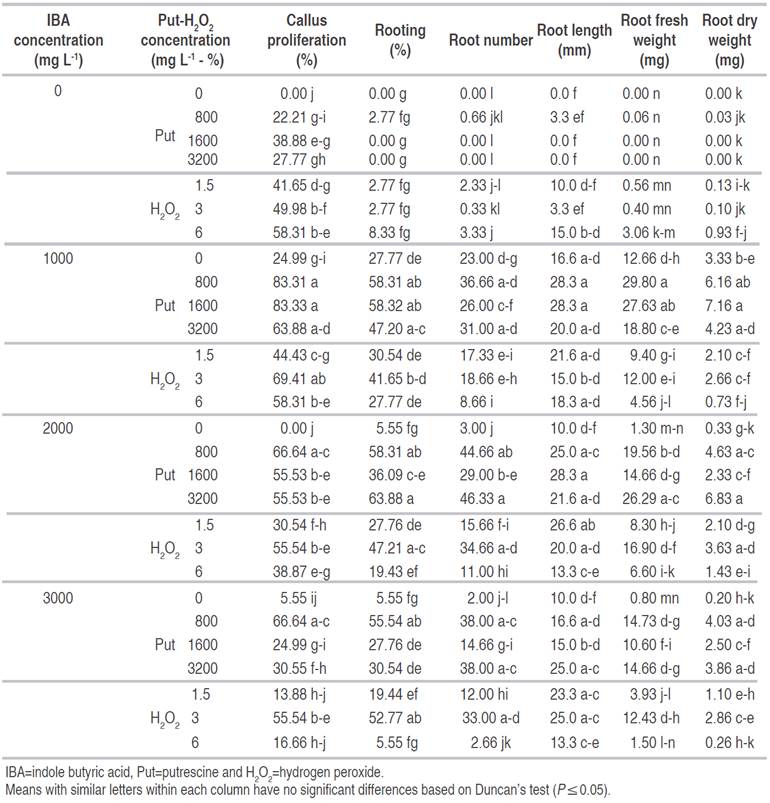
Application of IBA, Put and H2O2 and their interactions on rooting percentage were significant (P≤0.01) in cuttings taken in March. The highest rooting percentage (63.88%) was attained from cuttings treated by 2000 mg L-1 IBA+3200 mg L-1 Put (Figure 2), which did not have a significant difference with the treatments:1000 mg L-1 IBA+1600 mg L-1 Put (58.32%), 2000 mg L-1 IBA+ 800 mg L-1 Put (58.31%), 3000 mg L-1 IBA+3% H2O2 (52.77%), different levels of IBA with 800 mg L-1 Put, 2000 mg L-1 IBA+3% H2O2 (47.21%) (Figure 3), and 3200 mg L-1 Put+1000 mg L-1 IBA (47.20%). The lowest rooting percentage was resulted from control and treatments without IBA (Table 2).

Figure 2 GF677 semi-hardwood cuttings three months after treatment of 2000 mg L-1 IBA+3200 mg L-1 Put in March.

Figure 3 GF677 semi-hardwood cuttings three months after treatment of 2000 mg L-1 IBA+3% H2O2 in March.
The application of IBA, Put and H2O2 treatments and their interactions on root length were significant at (P≤0.01). The highest root length was obtained at 2000 mg L-1 IBA+1600 mg L-1 Put, 1000 mg L-1 IBA+1600 mg L-1 Put and 1000 mg L-1 IBA+800 mg L-1 Put. The root length for control and Put treatments with 1600 mg L-1 and 3200 mg L-1 was zero and had no significant difference with 800 mg L-1 Put, H2O2 (1.5 and 3%), IBA (2000 and 3000 mg L-1) treatments (Table 2).
The application of IBA, Put and H2O2 treatments and their interactions on root number were highly significant (P≤0.01). The highest root number (46.3) was observed in 2000 mg L-1 IBA+3200 mg L-1 Put, 2000 mg L-1 IBA+800 mg L-1 Put, 3000 mg L-1 IBA+800 mg L-1 Put, 1000 mg L-1 IBA+800 mg L-1 Put and 1000 mg L-1 IBA+3200 mg L-1 Put treatments. The lowest number of roots was recorded for control and 1600 and 3200 mg L-1 Put, while they did not have significant differences with 1.5 and 3% H2O2 and 3000 mg L-1 IBA treatments (Table 2).
The study showed that Put had some effects on root formation and its growth. Data on length and dry weight of roots illustrated that application of Put in the rooting of GF677 resulted in better quality roots compared with IBA treated cuttings. Moreover, concentrations of 800 and 1600 mg L-1 Put were better than its higher concentration (3200 mg L-1) on the above-mentioned traits. PAs have been showed to increase root elongation and growth by increasing cell division. The amount of Put increases during elongation in the differentiation zone, indicating that PAs are involved in root development (Tang and Newton, 2005). In fact, application of PAs increases the synthesis of internal PAs in plant tissue (Vondrakova et al., 2015). Increasingly, PAs are associated with increased mitotic activation and increased primary and lateral roots. The presence and involvement of genes are also related to the synthesis of Pas, which play a role in root development in the presence of PAs (Mahdavian et al., 2020). Supplementary using of PAs as a contemporary group of plant hormones considerably enhanced the number of rooting saplings of GF677.
This study revealed that Put by-itself did not have much effect on rooting of GF677 cuttings; however, a greatest effect was seen when IBA was used. IBA generates the root induction and Put stimulates the root growth, increasing synergism between both. Similar results also were obtained by Karimi and Yadollahi (2012). They showed that the highest weight of dry roots was attained with 2 mM Put treatments, and the lowest weight of roots was observed with treatment of 3000 mg L-1 IBA. The rate of saplings along callus was substantially more exclusive in 2 and 4 mM Put and 1500 mg L-1 IBA treatment and the lowest number of the saplings with callus was noticed with a treatment of 3000 mg L-1 IBA (Karimi and Yadollahi, 2012).
Third experiment
The application of IBA, Put and H2O2 and their interactions on callus proliferation percentage were significant (P≤0.01). The highest percentage of callus proliferation was observed under the treatments: 800 mg L-1 Put (55.55%), 1000 mg L-1 IBA+800 mg L-1 Put (41.65%), 3% H2O2 (36.1%), 1000 mg L-1 IBA (30.54%) and 2000 mg L-1 IBA (24.99%) (Table 3).
Table 3 The interaction effect of IBA, Put and H2O2 on rooting and callus proliferation of GF677 semi hardwood cuttings in October.
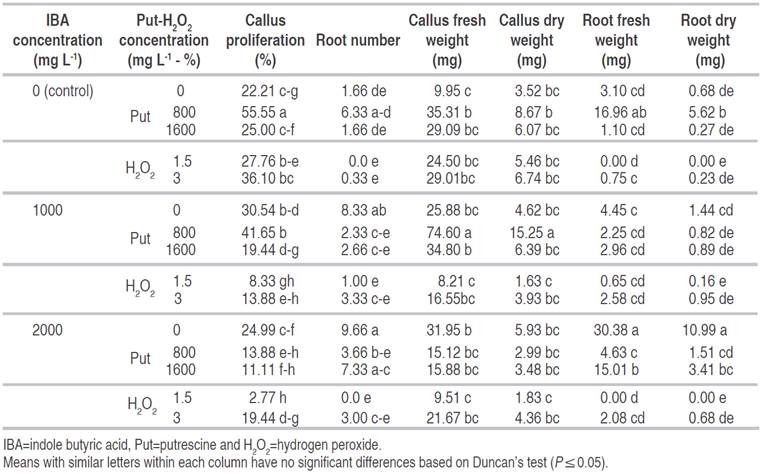
Data analysis showed significant effects of IBA (P≤0.05), Put and H2O2 (P≤0.01) and their interactions (P≤0.01) on callus fresh weight. The highest callus fresh weight was observed at 1000 mg L-1 IBA+800 mg L-1 Put (74.6 mg) and the lowest amount of callus fresh weight was recorded for control (9.95 mg), 2000 mg L-1 IBA+1.5% H2O2 (9.51 mg) and 1000 mg L-1 IBA+1.5% H2O2 (8.21 mg) (Table 3).
The effect of IBA (P≤0.05), Put and H2O2 (P≤0.01) and their interactions (P≤0.01) on callus dry weight was significant. Highest callus dry weight (15.25 mg) was obtained in cutting treated by 1000 mg L-1 IBA+800 mg L-1 Put. Also, the least callus dry weight observed from 2000 mg L-1 IBA+1.5% H2O2 (1.83 mg), and 1000 mg L-1 IBA+1.5% H2O2 (1.63 mg), while they did not have a significant difference with control (Table 3).
The IBA treatment had no significant difference with the interaction treatments on rooting percentage. There were no significant differences among different amounts of IBA, Put and H2O2 with control. The rooting percentage in 1.5% H2O2 treatment (0.92) decreased in comparison with control (Table 4). The activity of the enzymes at the rooting zone of stem cuttings may facilitate easy and rapid cell differentiation toward rooting. Peroxidase is an enzyme that initially increases its activity in root initiation and root development and has a positive effect on rooting. In addition, it is used as a marker in rooting to improve and enhance rooting (Hartmann et al., 2011). H2O2 is a co-enzyme that catalyzes the oxidation of a variety of organic compounds. Numerous rooting studies have shown that H2O2 plays an essential role in rooting of cuttings (Sebastiani and Tognetti, 2004; Shi-Weng et al., 2009a and b). Shi-Weng et al. (2009a) demonstrated that H2O2 may act as a signal molecule, involved in the auxin-induced formation of adventitious root formation. Their results showed that higher concentrations of H2O2 were required during the induction of adventitious root formation. Moreover, applying of IBA with H2O2 significantly refined the rooting of saplings compare to untreated ones; but the rooting percentage was low compared when only IBA was used. According to Sebastiani and Tognetti (2004), IBA+H2O2 caused considerable higher root number comparing with only IBA treatment on olive cultivars. The effect of Put and H2O2 treatments on root length was significant at (P≤0.01), while IBA treatment and their interactions were not significant. The highest root length was observed with 800 mg L-1 Put treatment (23.4 mm) and the least root length was obtained at 3% H2O2 treatment with (4.8 mm) (Table 4).
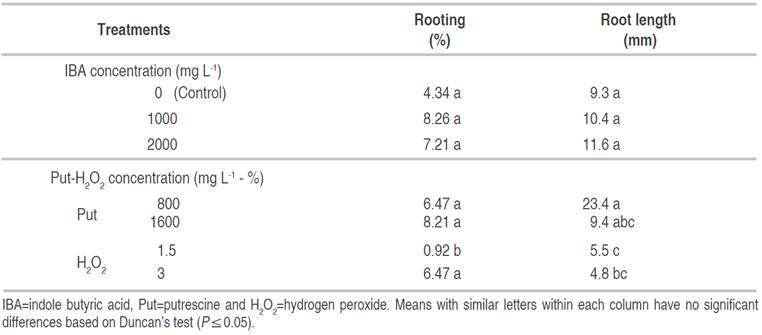
Analysis of variance showed significant effect of Put and H2O2 (P≤0.01), IBA (P≤0.05) and their interactions (P≤0.05) on root number. The Highest root number was obtained at 2000 mg L-1 IBA only, 1000 mg L-1 IBA only and 2000 mg L-1 IBA+1600 mg L-1 Put. The least root number resulted under treatments of: 1.5 and 3% H2O2, 1000 mg L-1 IBA+1.5% H2O2 and 2000 mg L-1 IBA+1.5% H2O2, which had no difference with control, 1600 mg L-1 Put, 1000 mg L-1 IBA+800 mg L-1 Put, 1000 mg L-1 IBA+1600 mg L-1 Put, 1000 mg L-1 IBA+3% H2O2, 2000 mg L-1 IBA+800 mg L-1 Put and 2000 mg L-1 IBA+3% H2O2 treatments (Table 3).
The application of IBA, Put and H2O2 treatments and their interactions on fresh and dry weight of roots were highly significant (P≤0.01). The most root fresh weight (30.38 mg) was observed by 2000 mg L-1 IBA treatment, whereas the least root fresh weight was obtained using the treatments: 1.5% H2O2 (0 mg), 2000 mg L-1 IBA+ 1.5% H2O2 (0 mg), 1600 mg L-1 Put (1.1 mg), 1000 mg L-1 IBA+1.5% H2O2 (0.65 mg) and 3% H2O2 (0.75 mg) (Table 3). The effects of IBA, Put and H2O2 treatments and their interactions on root dry weight were significant at (P≤0.01). The highest root dry weight resulted using 2000 mg L-1 IBA. Moreover, root dry weight in 1.5% H2O2 and 2000 mg L-1 IBA+1.5% H2O2 were zero, which had no significant difference with control and other treatments (Table 3). In the present study, the use of the only auxin (IBA) produced callus and rooting in the three times of cutting collection. The need of using auxin for rooting of GF677 cuttings has been reported in previous studies (Tsipouridis and Thomidis. 2004; Tsipouridis et al., 2005; Karimi and Yadollahi, 2012). Based on Tsipouridis et al. (2005) the appropriate concentration is between 500 and 2500 mg L-1 IBA. Similar results were obtained by Karimi and Yadollahi (2012) who reported rooting of 57% of GF677 cuttings when treated by 1500 mg L-1 IBA using a quick-dip method. The rooting decreased at 3000 mg L-1 IBA. High concentrations of IBA may have toxic effects on cuttings, thereby reducing the rooting (Karimi and Yadollahi, 2012). In addition, in this study, it was observed that time of cuttings preparation was very effective on callus formation and rooting of GF677 cuttings. Little rooting was observed in cuttings prepared in July and October. In contrast, rooting was much higher in those cuttings that were taken in March. The rooting success of peach cuttings has been reported to be affected by the date of collection of cuttings (Tworkoski and Takeda, 2007). The effect of cuttings collection date on the rooting of many cuttings, including olives (Hartmann and Loreti, 1965; Khajehpour et al., 2014) and peach (Tofanelli et al., 2003), has been demonstrated. It seems that several factors related to the time of preparation of cuttings can be effective on rooting of cuttings (Hartmann et al., 2011). Khajehpour et al. (2014) observed no rooting on the cuttings collected in October, but the cuttings collected in August rooted well. They also reported better rooting in cuttings taken in March compared to those taken in late summer or early autumn (Khajehpour et al., 2014). However, conflicting results have been reported by other researchers. Tofanelli et al. (2003) for peach cuttings reported that the most favorable time for collecting of cuttings is between October 25 and November 13.
CONCLUSION
The best results of callus formation and rooting were observed in cuttings prepared in March. The current experiment showed that the application of IBA, Put and H2O2 in comparison with control, increased the rooting of hard and semi-hardwood cuttings of GF677 rootstock. Treatment of cuttings by only IBA was more effective than only Put or H2O2. Additionally, IBA at concentrations of 1000 and 2000 mg L-1 indicated better influence on rooting. Also, the use of inexpensive chemicals such as Put or H2O2 together with IBA had a positive effect on increasing the rooting of GF677 cuttings, and the use of Put had a greater effect on root growth. The high concentration of only Put at 3200 mg L-1 and H2O2 at 6% caused the least effect on measured traits while these dosages combined with IBA showed a favorable effect on mentioned factors.

