











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
Permalink
Lulo (Solanum quitoense Lam.) is a crop that belongs to the section Lasiocarpa, family Solanaceae, and genus Solanum. Its center of origin is located between the Colombian and Ecuadorian Andes (Heiser and Anderson, 1999; Lobo and Medina, 2000; Ramírez and Davenport, 2020; Ramírez 2021). Several authors, including to Ramírez et al. (2018); Arias and Rendón (2014); Muñoz et al. (2013); Paull and Duarte (2012) and Medina et al. (2009), mention various factors that make lulo a promising fruit-tree with high productive potential, including a high genetic variability with related species in the Andean region, proper niches for planting, consumer acceptance of the fruit and agroindustrial potential.
Despite the productive potential of lulo, there is a lack of development of this crop. According to Agronet (2019), Colombia has a planted area of 8,821.35 ha with a production of 89,050.41 t and an average yield of 10.1 t ha-1. The main producers are the departments of Huila, Valle del Cauca, Antioquia and Boyacá, with production ranging from 8,424 to 14,339.8 t. The cultivated area, production and yield in the department of Nariño have decreased considerably from 2007 to 2016 about of 6, 17 and 12%, respectively. This behavior may be caused, among others, by the dynamics of production systems based mainly on a scheme lacking advanced technologies and a shortage of planting materials (Almanza et al., 2016), since genetic improvement programs in the region have been directed primarily to industrial crops and crops of importance in food security (Pareja et al., 2010). For this reason, despite having suitable soil and climatic conditions for the crop, it does not reach its potential under competitive growing conditions (Casierra et al., 2013).
One of the most important factors that has reduced the productivity of lulo crops in the department of Nariño, especially in cultivars (e.g. Castilla and Larga Vida) planted by growers, is the susceptibility to vascular and root wilt caused by Fusarium oxysporum. In most cases, this problem reduces productivity, delays the crop cycle, and leads the death of many plants. Therefore, genotypes with resistance genes that allow counteracting the disease must be identified and selected (Díaz et al. 2011). F. oxysporum has caused economic losses ranging from 50 to 90% in commercial crops (Lagos et al. 2015), and it is characterized by visible symptoms, such as chlorosis and loss of turgor, causing complete wilting of the plant that develops from the germination of chlamydospores that come into contact with the roots, generate an appressoria that penetrates the root cortex and develops an internal mycelium in the root and advances until it reaches the vascular system (xylem ducts) (Cruz et al. 2011). Internally, the vascular bundles display discoloration or ascending necrosis that is evident through a cross-section of the main stem (Ochoa, 2002). This disease is considered difficult to manage since F. oxysporum can survive in plant residues and soil due to resistant structures known as chlamydospores (Agrios, 2005). The survival period of this fungus can be up to 20 years, triggering the loss of soil productivity for new plantings (Estupiñan and Ossa, 2007).
The control of this pathogen generally involves the excessive use of chemical fertilizers that affect human health and cause further environmental degradation (Villa-Martínez et al. 2014). Therefore, in an integrated crop management approach, disease management, environmental protection and reduction of the use of agrochemicals are prioritized actions, requiring strategies that include agronomic and cultivar improvement practices that take advantage of the diversity associated with the species. However, these practices require the adoption of processes and technology transfer, which are often difficult to implement (Ochoa, 2002; Ochoa and Gallardo, 2004; Clavijo, 2014; Arizala et al., 2011; Cardona, 2013).
A viable option for disease management is genetic resistance, in which the assessment of genotypes focuses on the search for sources of resistance. This alternative provides many advantages, for instance, it is environmentally friendly, reduces the possibility of developing pathogen resistance to agrochemicals, and provides low production costs due to the reduced use of agrochemicals (Clavijo, 2014).
Several studies on different commercial species have studied the sources of resistance to F. oxysporum obtaining a great contribution to the genetic improvement processes, such as García-Velasco et al. (2020), who evaluated the resistance or susceptibility of Musa sp. cultivars in Cuba, using different filtrates of F. oxysporum, allowing the establishment of an efficient and non-destructive method for the identification of races 1 and 2 of this pathogen, Carvalho et al. (2021) identified Passiflora nitida, P. foetida and P. mucronatacan species as sources of resistance to F. oxysporum and F. solani complex and recommended them for use in Passiflora breeding programs of Brazil and Shaw et al. (2017) identified a dominant nature of resistance in Ricinus communis inbred line AP42 to F. oxysporum f. sp. ricini, which is of great interest in hybridization programs in India, among others.
At national level, studies carried out by Tamayo et al. (2002) assessed the genetic resistance of the interspecific lulo hybrid "La Selva" (S. quitoense x S. hirtum); in particular, the authors recommend this material as a control alternative, with economic impact, in areas where the development of this crop is hindered by the pathogen. In other Solanaceae, such as Capsicum spp., Clavijo (2014) identified six accessions with resistance genes, which are recommended as parental plants in chili pepper improvement programs. Morales et al. (2014) assessed wild and cultivated accessions of Solanum lycopersicum and found sources of resistance in Solanum neorickii; hence, the authors recommended this species as a parent in interspecific breeding programs. Mayorga-Cubillos et al. (2019) identified six genotypes of cape gooseberry (Physalis peruviana) that are promising against F. oxysporum and can be used in subsequent breeding schemes.
In this context, the study hypothesis is based on the fact that within the working collection of lulo there are no genotypes with characteristics of tolerance or resistance to the artificial inoculation of F. oxysporum and the objective of this work was to evaluate the response of 22 lulo genotypes, from the GPFA (Grupo de Investigación en Produccion de Frutales Andinos) collection, to the artificial inoculation of Fusarium oxysporum Schelcht, the causal agent vascular and root wilt and to select sources of resistance that can be useful in lulo breeding programs.
MATERIALS AND METHODS
Location
The experiment was carried out at the greenhouse of Botana Experimental Farm of Universidad de Nariño at 2,670 masl (01°09'30.62"NL, 77°16'31.79"WL), with an average outdoor temperature of 12°C, average indoor greenhouse temperature of 22°C and 80% relative humidity.
Plant material
As plant material, 22 lulo genotypes were used, including 10 selected based on field-resistance to F. oxysporum and good agronomic traits obtained by sexual seed propagation (Table 1), as well as 12 genotypes from half-sibling families (HSF) propagated by cuttings were evaluated. The 22 genotypes belonged to the GPFA collection of Universidad de Nariño (Table 2).
Table 1 Field-resistance lulo genotypes to Fusarium oxysporum derived from sexual seed, under natural conditions of the region of the department of Nariño, Colombia.
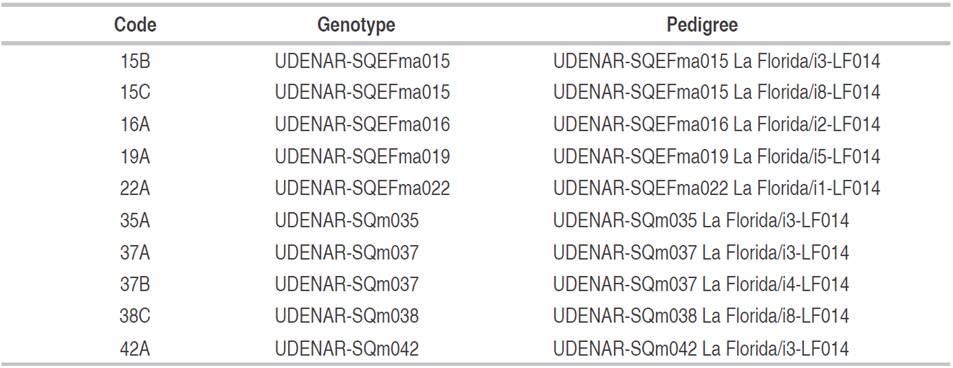
Table 2 Half-sibling families of lulo derived from cuttings, obtained through individual stratified selection of the region of the department of Nariño, Colombia.

The 12 HSF (Table 2) are derived from two selection stages. In the first stage, a commercial crop plantation was established, using seeds of the "La Selva" cultivar, located in Nariño (01°07'24.28"N,77°26'19.66"W), at 2,237 masl. A stratified selection was made from this plantation; the best plant was selected from each stratum according to yield, fruit quality, health and architecture. To create 50 HSF, 50 plants were selected.
In the second stage, 50 HSF were planted in experimental trials in four localities of the department of Nariño, namely La Unión, San Pedro de Cartago, Arboleda and Tangua, which are located between 1,700 and 2,100 masl. The experiment was a completely randomized block design with three repetitions and the families as treatments. At each locality, growth, yield, and fruit quality were assessed as the response variables. The data were analyzed through an Analysis of Variance and a selection index was applied to the most important variables. Based on this, 12 HSF were selected and propagated taking 15 cm long, healthy cuttings with at least two axillary buds that were planted in sterile substrate for rooting and then, evaluated by their response to artificial inoculation by F. oxysporum.
Inoculation of the lulo genotypes
Selected lulo genotypes were inoculated with F. oxysporum (Fo) isolate Fo15, collected in the municipality of Buesaco, Nariño, Colombia and evaluated for its pathogenicity on S. quitoense and S. hirtum, generating severity of 96% and 84%, respectively (Duarte-Alvarado et al., 2020). The five inoculated plants per genotype showed at least four true leaves and a root growth without phytosanitary problems. The inoculation method based on wounded roots was applied (Clavijo, 2014), which consisted in washing the roots with distilled water, making small cuts in the apices, and submerging the roots in previously prepared inoculum for 3 h, then, the plants were sowed in a sterile substrate. The control was subjected under the same treatment, except the immersion step in sterile distilled water (Betancourth, 2005; Clavijo, 2014).
For the preparation of the inoculum, a spore suspension of pure monospore cultures grown in PDA at 28°C for 7 to 10 days was used. Subsequently, 20 mL of sterile distilled water was added and a superficial scraping with a sterile spatula to remove conidia and mycelium. Afterwards, the solution was passed through a filter to separate the conidia from the mycelium and adjusted to a concentration of 1x106 conidia mL-1 by a haematocytometer with a Neubauer chamber (Clavijo, 2014).
The plants with symptoms of vascular and root wilt caused by F. oxysporum were taken to the laboratory to confirm Koch's postulates (Koch, 1876). The genotypes that did not show disease symptoms and survived during the trial period were considered to be resistant. Also, to confirm these results, re-inoculations with a mix of four isolates, namely Fo1, Fo2, Fo16, and Fo19, which cause 100% severity in S. quitoense were performed according to the study carried out by Duarte-Alvarado et al. (2020).
Experimental design
A completely randomized design with five repetitions with the 22 lulo genotypes (Tables 1 and 2) as treatments was implemented. Each repetition comprised five plants. The experimental unit corresponds to a plant grown in a pot with sterile substrate (pH of 5.5, 88.7% organic matter, 758.29 ppm of N, 499.35 ppm of P, 2,371.25 ppm of K), 110 experimental units in total. Two controls were used; the inoculation control was Castilla cultivar inoculated with distilled water and Fo15 and the resistant control was "La Selva" hybrid inoculated with isolate Fo15 and sterile distilled water (Tamayo et al., 2002).
Disease assessment
The response of the genotypes to the inoculation by measuring the following variables:
Plant height (PH) (cm) was measured for the genotypes propagated by seeds (Table 1). Following inoculation, the PH was recorded every seven days for three months. After the increase in PH, the ΔPH was calculated through the Equation 1:
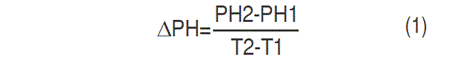
Where: PH1=plant height in cm at T1, AP2=plant height (cm) at T2, T1=initial time of PH measurement, T2=final time of PH measurement.
Stem diameter (SD), similar to PH, SD (cm) was assessed for the genotypes propagated by seeds (Table 1). Following inoculation, the SD was recorded every seven days for three months. The increase in SD (ΔSD) was calculated through the Equation 2:
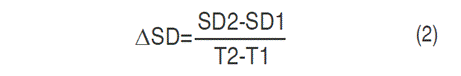
Where: SD1=stem diameter in cm at T1, SD2=stem diameter in cm at T2, T1=initial time of SD measurement, T2=final time of SD measurement.
Considering the nature of the reproduction of the genotypes evaluated, the variables PH, SD were taken only in those genotypes multiplied by sexual seed (Table 1), given that the root system of plants propagated by cuttings does not allow these two variables to be taken with precision.
The following variables were evaluated for the 22 genotypes, including 10 derived from sexual reproduction (Table 1) and 12 from asexual reproduction (Table 2):
Days to the onset of symptoms (DOS), number of days from the moment of inoculation to the onset of disease symptoms.
Incidence (I) expressed as percentage, based on the number of diseased plants divided by the total number of plants assessed.
Severity (S) developed by each plant during 84 days of evaluation was calculated based on the scale proposed by Elmer and Robert (2004) (Table 3).

The average S(%) was calculated according to the Equation 3 (Song et al., 2004), which is estimated according to the scale in Table 3 and expresses the severity as an index.
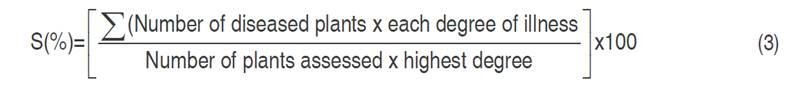
Re-isolation of the fungus
The fungus from the diseased plant material was isolated to determine if the symptoms observed were caused by F. oxysporum. Likewise, the isolation from the vascular region of asymptomatic plants to determine if there was a slight infection in the basal part of the stem in any of the genotypes was performed; to demonstrate possible asymptomatic carriers (Estupiñan and Ossa, 2007).
Data analysis
An analysis of variance on the data using S.A.S 9.3 software (Statistical Analysis System, Institute Inc.) was performed. When the null hypothesis was rejected, there were differences in the variables between treatments. This was determined according to De la Cruz et al. (2010), who established significant differences between treatments when the upper values exceed the mean plus one standard error (μ+σ) and highly significant when the upper values exceed the mean plus two standard errors (μ+2σ). In this case, for disease I and S, the most outstanding genetic materials were those found below μ-σ or μ-2σ, with significant or highly significant differences compared to the other genotypes.
RESULTS AND DISCUSSION
There were significant differences for plant height and stem diameter, between the genotypes assessed through sexual seed (Table 4). Genotypes 22A, 15B, 15C and Castilla showed the highest ΔPH with values between 0.34 and 0.41 cm day-1 in comparison with the control, indicating that the infection process caused by the fungus within the vascular system of these genotypes, generated a relatively low decrease in apical growth, which may be due to a mild or first-stage infection process corresponding to epidermal fixation or penetration and does not progress to a second stage, which corresponds to intravascular colonization (Gonzales et al., 2012). Similarly, it can be considered that as a mechanism of infection, the pathogen can take two routes, initially hemibiotrophic and then necrotrophic (Perfect and Green, 2001), considering in this case that only the first one occurred.
Table 4 ANOVA mean squares for days to onset of symptoms (DOS), plant height (ΔPH) and stem diameter (ΔSD), evaluated in 10 lulo genotypes derived from sexual seeds inoculated with F. oxysporum under greenhouse conditions.
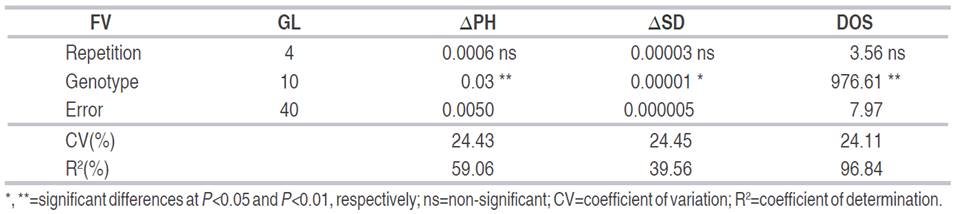
Genotype 42A showed the lowest average ΔPH with 0.14±0.06 cm day-1 with a more severe intravascular colonization process and affecting the apical growth (Table 5).
Table 5 Averages of the variables plant height (ΔPH) and stem diameter (ΔSD) in 10 genotypes of lulo obtained through sexual seed.
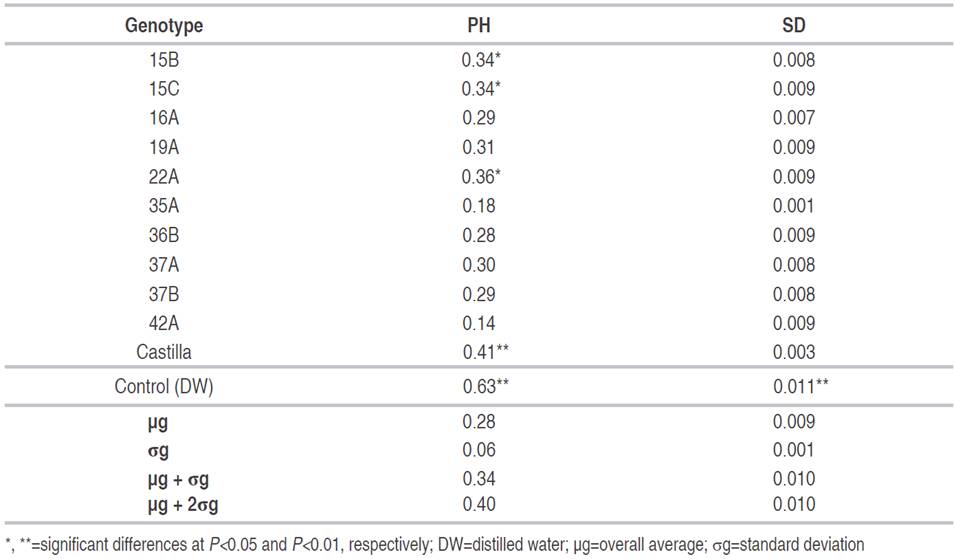
For ΔSD, no genotype showed a significantly different mean, except for the control that showed a highly significant difference compared to the other genotypes. The ΔSD ranged from 0.001 cm day-1 for 35A to 0.09 cm day-1 for 15C, 19A, 22A, 36B, and 42A (Table 5). In 35A and Castilla, the values of ΔSD indicates that the infection process is compromising the normal development of the stem by blocking vascular ducts. According to González et al. (2012), these results showed that exist a colonization of the mycelium into the xylem and the establishment of the fungus in the plant via this tissue. This invasion through the vascular bundles causes symptoms such as dwarfism, wilt, and loss of turgor since nutrient and water transport are affected by the colonization of this pathogen, This coincides with the statements of Chekali et al. (2011) and Cruz et al. (2011), who affirmed that as a typical response to the fungus, metabolic and growth functions are altered in plants and that according to their evolution and aggressiveness, which vary by their age, their susceptibility to the pathogen and environmental conditions, they can even cause their death. The results obtained are also corroborated with similar works carried out by Clavijo (2014), which by means of pathogenicity tests with Fusarium oxysporum in the chili bell pepper crop was demonstrated that in addition to the susceptibility of this crop to the pathogen, it influenced in the growth variables such as height and stem diameter at the end of its evaluation.
For DOS variable, there were highly significant differences between genotypes (Table 4). Among the 22 genotypes, 44% showed a disease incubation period between 7 and 14 days. These results differ from those reported by Manangón et al. (2015), Maya and Lagos (2011) and Narváez and Zambrano (2006), who obtained incubation averages between 21 and 46 days in different accessions of the Lasiocarpa section under controlled conditions and can be explained by the difference in the infective capacity of F. oxysporum isolates and the resistance or susceptibility of the genetic materials evaluated (González et al., 2012).
Genotypes 15C, 36B, HSF1, HSF10, HSF36 and control did not show signs of vascular and root wilt during the trial period (Table 6). However, the intensity of infection caused by F. oxysporum is often visually diagnosed by the typical symptoms of the disease, but it is not proportional with internal infection processes such as vascular colonization and it is necessary to evaluate organs such as root and stem to determine if an infectious process exists (Ríos et al., 2018). To confirm these findings, we re-inoculated with isolates Fo1, Fo2, Fo16, and Fo19 after three months of the assessment; there was not visible disease symptoms, vascular colonization processes or any type of root necrosis.
Table 6 Average days to onset of symptoms (DOS) in 22 lulo genotypes inoculated with Fusarium oxysporum under greenhouse conditions.
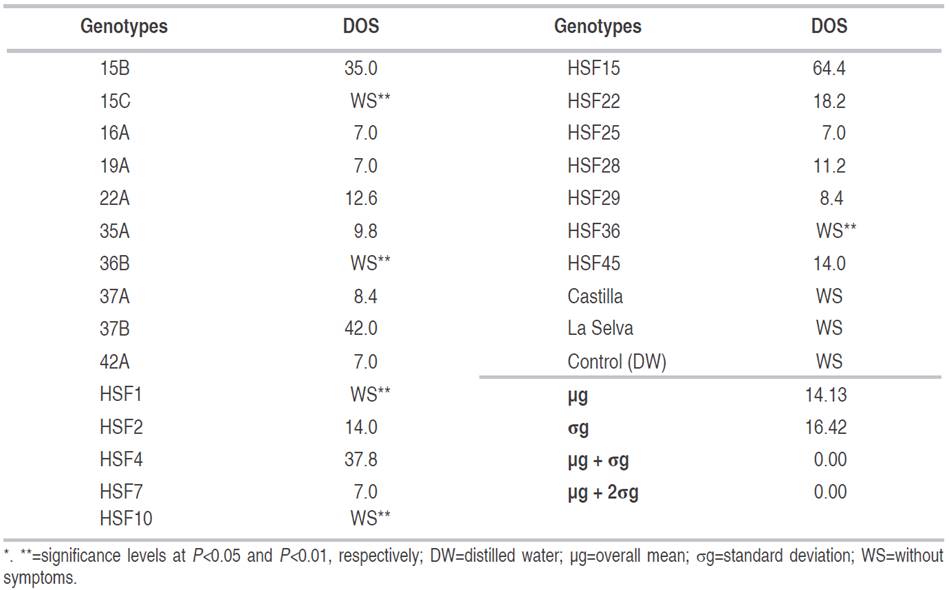
Genotypes 16A, 19A, 42A, HSF7 and HSF25 showed the lowest averages, with the onset of symptoms at seven days post-inoculation (DPI), as demonstrated by chlorotic leaves and loss of turgor. The highest average was obtained by HSF15 with 64.4 DPI, followed by 37B with 42 DPI, and HSF4 with 37.8 DPI (Table 6).
It is worth mentioning that Ortiz (2011) reported incubation periods of 8 to 10 days for plants inoculated with F. oxysporum and subjected to excess water stress in another Solanaceae (Physalis peruviana) and 18 to 24 days for plants also inoculated under normal humidity conditions. These data added to those reported by Clavijo (2014), who found the appearance of symptoms such as yellowing, necrosis and defoliation in about six days after inoculation with the pathogen, suggest variability in the first place, with respect to the environmental influence on the expression of the pathogen and secondly, to the virulence of the inoculated strain. This aspect confirms the importance of the response of the genotypes that were re-inoculated, since their initial behavior did not vary.
Incidences (I) of vascular wilt were between 0 and 100%. Specifically, 77.3% of the genotypes (17 genotypes) showed I of 100% and 22.7% showed 0% incidences (5 genotypes). Genotypes 15C, 36B, HSF1, HSF10 and HSF36 displayed 0% I and S; thus, demonstrating resistance to isolate Fo15 and to the mix of isolates Fo1, Fo2, Fo16 and Fo19. These genotypes could be used in genetic improvement programs, evaluated as parents in different environments to determine their genetic potential and adaptation to the producing areas of the department of Nariño, based on the fact that the progress of these programs is more efficient when there is an adequate characterization of the attributes of interest in a population of a given species (Morillo et al., 2019).
For the control of this disease, numerous practices and activities have been reported that are generally based on the use of agrochemicals with their economic and environmental implications. That is the reason why the genetic improvement of resistance as a form of effective control for this disease has allowed to develop increasingly efficient strategies (García-Velasco et al., 2020). In this regard, it is worth considering that the Fusarium genus, due to its great variability, is one of the most difficult to manage of all fungal groups (Dean et al., 2012). Thus, the use of resistant varieties could reduce the incidence of the disease and for this purpose it is necessary to have genotypes within the species of interest that are favorable in this aspect (Horinouchi et al., 2011). This situation confirms the importance of identifying the genotypes presented here as candidates for genetic improvement programs.
In contrast, genotypes 42A, 35A, 16A, HSF2, HSF7, HSF25 and HSF45 showed I of 100% and S between 72% and 88% (Figures 1 and 2), being highly susceptible to pathogen, with symptoms of generalized chlorosis, necrosis of new leaves, necrosis in main leaves, and loss of turgor. These results are consistent with those obtained by Narváez and Zambrano (2006) and Gallardo (2005), who evaluated different genotypes of S. quitoense and between 9 and 11 week of evaluation reached the highest level of severity in 90% of their seedlings and the remaining 10% presented low levels as chlorosis and flaccidity in the lower leaves. On the other hand, it is also worth mentioning the work of Arizala et al. (2011), who evaluated the response to F. oxysporum of wild species of lulo, in which they highlighted low incidence for Solanum hirtum and Solanum marginatum, the former with physical compatibility in the use of grafts with S. quitoense, which is the reason why, in addition to those mentioned, it is possible to think about interspecific improvement processes for the study area.
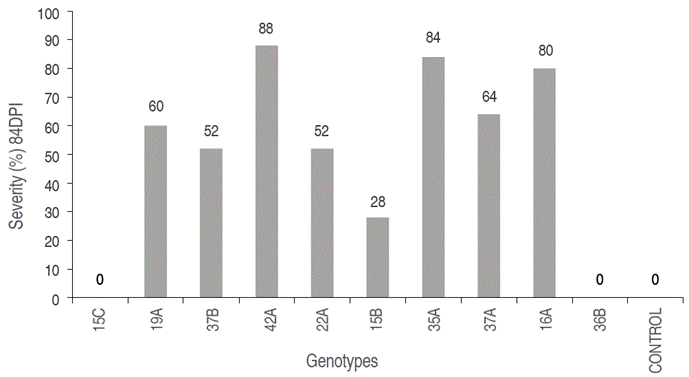
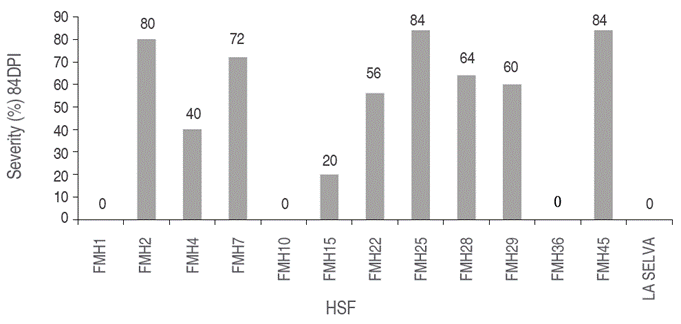
Figure 2 Average severity (%) at 84 DPI in 12 half sibling families (HSF) of lulo propagated by stakes.
Genotypes HSF15 and 15B showed S of 20% and 28%, respectively, with slight chlorosis in new leaves and a slight delay in growth. Also, these genotypes were re-inoculated with the mix of Fo1, Fo2, Fo16, and Fo19 obtaining S of 30% and 33%, finding that the two genotypes survived during the trial period, suggesting tolerance or a resistance mechanism different of the other genotypes. According to Forero et al. (2015), this expression of tolerance can be due to physiological factors, such as an increased concentration of chlorophyll in leaves or an increased number of new leaves or sprouts, which promote the emission and opening of floral buds or increase nutrient uptake. Also, these two genotypes, must be considered for evaluation in different environments to determinate his agronomic potential.
CONCLUSIONS
Regarding the growth variables evaluated, genotypes 22A, 15B and 15C stood out for showing the greatest increase in plant height over the susceptible Castilla control, which also, with respect to the increase in stem diameter, showed a lower value than the other genotypes, thus indicating the favorable behavior of the populations evaluated with respect to the presence of the disease.
Based on their differential response and with respect to the other genotypes in terms of periods of observation of symptoms, incidence and severity of the disease, genotypes 15C, 36B, HSF1, HSF10 and HSF36 did not show symptoms related to the inoculation of isolates Fo15 of F. oxysporum and the mixture of four other isolates, even after re-inoculation. This aspect is relevant for the genetic improvement of the species, since these could form the basis of a program based on resistance to Fusarium oxysporum for the department of Nariño, an evaluation that logically should be developed in an integral framework with other variables of productivity and agronomic behavior under cultivation conditions.

