











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
Permalink
The species hemp or cannabis (Cannabis sativa L.) belongs to the Cannabaceae family and its origin and uses date back to ancient times in Asia and the Middle East (Molina 2008; Clarke and Merlin 2013). Although it is considered a monotypic species, there are three subspecies (sativa, indica, and ruderalis) with highly polymorphic characteristics (Spitzer-Rimon et al. 2019). The cultivation of cannabis for medicinal or psychoactive purposes is based on the use of globular trichomes present in floral structures, mainly in female flowers, which is why the use of feminized genotypes is essential. Moreover, hermaphrodite plants produce fibers and seeds with an adequate ratio of male and female flowers (Razumova et al. 2016; Spitzer-Rimon et al. 2019).
This species is classified as a dioecious plant, which only occurs in 6% of all plant species. Both male and female inflorescences are found in different individuals. Their expression is linked to both hormonal homeostasis of the genes localized on the XX chromosomes in the case of female plants, and the genes on XY chromosomes in male plants (Negrutiu et al. 2001; Hobza et al. 2018). However, cases of hermaphroditism can occur, where both female and male flowers differ in the same individual (Salentijn et al. 2019). Both inflorescences develop orderly at the internodes in the axillary buds on the plants' main axis and lateral branches. Each phytomer (internode and axillary bud) comprises a large, webbed leaf in charge of photosynthetic processes, and a pair of bracts that support two single flowers originating from an axillary bud (Spitzer-Rimon et al. 2019). The female flowers have a simple structure comprising a perigonal bract covering the ovary. At maturity, the flower is covered with globular trichomes where Phyto cannabinoids are synthesized. It also has a pair of stigmas that emerge unevenly from the inside. Male flowers appear in hanging panicles with five sepals and five stamens; their anthers disperse pollen when they reach maturity -a process favored by the wind (Moliterni et al. 2004; Cervantes 2007).
Given the species evolution, several environmental and genetic factors are responsible for the physiological process of flowering, e.g., temperature and photoperiod, which directly influence the morphology of the genesis flower (Amaducci et al. 2008). Cannabis is considered a photoperiod plant and therefore responds to day length; when days are shortened (fewer light hours), the plant induces the flowering process and reaches the reproductive stage. Plants from seed reach maturity at the fourth phytomer, when the differentiation of the apical bud, located on the main stem, generally appears (Moliterni et al. 2004). Here, single flowers are observed on the bracts, some reach their anthesis; however, it can continue from axillary buds until the plants are subjected to short days (suspension of additional light). Some changes in the architecture and morphology of the plants are observed (Amaducci et al. 2008; Spitzer-Rimon et al. 2019).
Under appropriate age, temperature, and luminosity, the genetic expression that regulates sexual differentiation in plants -closely related to environmental interactions and hormonal synthesis- is triggered (Sarath and Mohan Ram 1979; Blazquez and Weigel 2000; Moliterni et al. 2004). Several authors found that the production of female flowers in cannabis plants is favored by increasing the concentrations of auxins, cytokinins, and ethylene. In contrast, the masculine ones were favored by increasing gibberellins (Sarath and Mohan Ram 1979; Mohan Ram and Sett 1982b; Lubell and Brand 2018).
Cannabis growers use sexual reversion methods to produce pollen with male gametes, which leads to sexual seed production when crossing them with female plants. The XX genes dominate the female's originating plant (Lubell and Brand 2018). Silver thiosulfate (AgNO3) is the most used compound by seed producers to achieve sexual reversion in female plants; however, it is highly toxic for handling and to soil and water sources. Other alternatives are substances such as aminoethoxy-vinyl-glycine (AVG) and 1-Methylcyclopropene (1-MCP), whose molecules are less toxic for humans, animals, and the environment (Blankenship and Dole 2003). Studies by Mohan Ram and Sett (1982b) and Lubell and Brand (2018) demonstrated the efficacy of STS and AVG to produce male flowers with viable pollen from female cannabis plants. Therefore, the present work aims to evaluate different methods of sexual reversion in female cannabis plants as a preliminary part of the species breeding program in the search for genotypes, anticipating the industrial growth that the cultivation of cannabis could have in the country.
MATERIALS AND METHODS
Location
The trials were developed in the municipality of Bello, Antioquia, at the facilities of Breeder's of Colombia SAS. This company is registered before the Colombian Agricultural Institute (ICA by its Spanish acronym) as a cannabis breeding research unit for psychoactive cannabis, according to resolution No. 00030034 of August 14, 2018.
Experimental design
A completely randomized design with eight treatments and four repetitions was used. The treatments consisted of protocols for the sexual reversion of female plants through apex drip and foliar spraying of Silver Thiosulfate (STS), Aminoethoxy-vinyl-glycine (AVG), 1-Methylcyclopropene (1-MCP), and Gibberellic acid (AG3), plus a control treatment without application. Each compound was prepared separately and diluted in demineralized water plus Tween-80 (0.01%) as a surfactant (Table 1).
Table 1 Dosage and application of the compounds used for the sexual reversion of female cannabis plants.
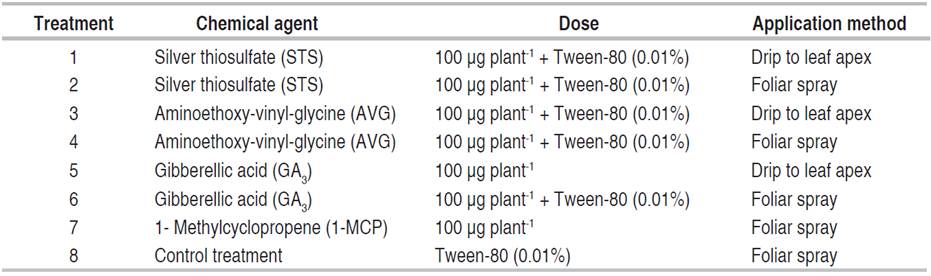
Cuttings obtained from female cannabis plants were sown in 10 L plastic pots using the substrate of coconut fiber, rice, mushroom, and sawdust in equal proportions; then, they were placed under a plastic cover and subjected to a photoperiod of 18/6 h (Day/Night). Out of the 18 h a day, the first 12 h correspond to sunlight and the remaining 6 h were provided by 30 W and 5000 K LED bulbs. The pots were distributed in three rows with 12 plants each at 0.5 m between plants and 1m between rows.
Each treatment was applied 20 µg plant-1 for 5 consecutive days and 100 µg of the compound per plant. The application was made 60 days after pot transplantation, when the additional light was suspended (6 h), thus modifying the relationship to 12/12 h (Day/Night).
Each compound (treatment) was applied through foliar spray (s) using a 20 µg solution in 10 mL of distilled water +0.01% Tween-80. The drip to the foliar apex (g) consisted of applying a 10 µL drop of a solution containing 20 µg of each compound with a micropipette. In the 1-MCP treatment, the application consisted only of foliar spraying. In contrast, the control treatment plants only received the Tween-80 solution (0.01%) in 20 mL of water applied in foliar spraying.
Number of male flowers and pollen viability
The male flowers in anthesis (open petals and mature anthers) were collected weekly and were counted by plant. The pollen of each male flower was extracted by vibration and deposited on a slide fixed with a drop of tetrazolium salt. After 30 seconds, the pollen grains were observed in a microscope under a 10X objective, and 200 grains were counted. Subsequently, their viability status was determined; those that presented a reddish coloration were considered viable, and those without coloration, non-viable.
Data analysis
An analysis of variance and Shapiro-Wilk test were performed for normality and the results were subjected to the Tukey Multiple comparison test (P<0.05). In cases where the normality assumption was not fulfilled, the non-parametric Kruskal-Wallis' test was used. The differences between treatments were analyzed by the Wilcoxon signed-rank test using the Agricolae package of the R statistical software Core Team 2023.
RESULTS AND DISCUSSION
Number of male flowers
Since data did not comply with the normality assumption (Shapiro-Wilk, W=0.46779, and P<2.2x10-16), they were analyzed under the non-parametric Kruskal-Wallis’ test (chi-square of 94.345, df=7 and P<2.2x10-16). The comparisons were made by the Wilcoxon test adjusted to the Holm method (Table 2).
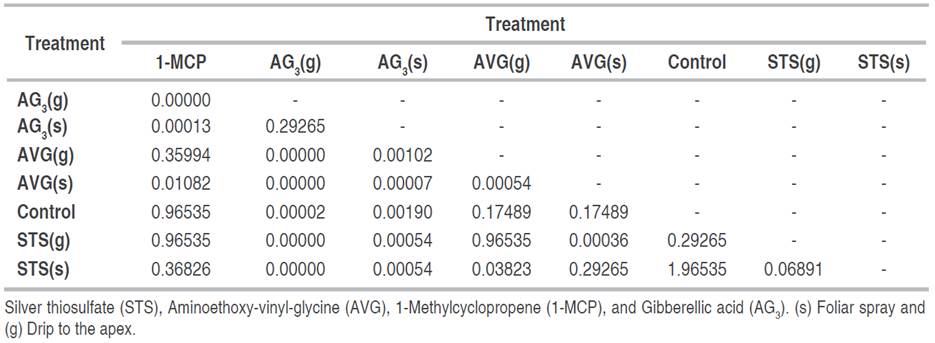
Table 2 P values comparing the median of the sexual reversion treatments of female Cannabis plants by Wilcoxon test.
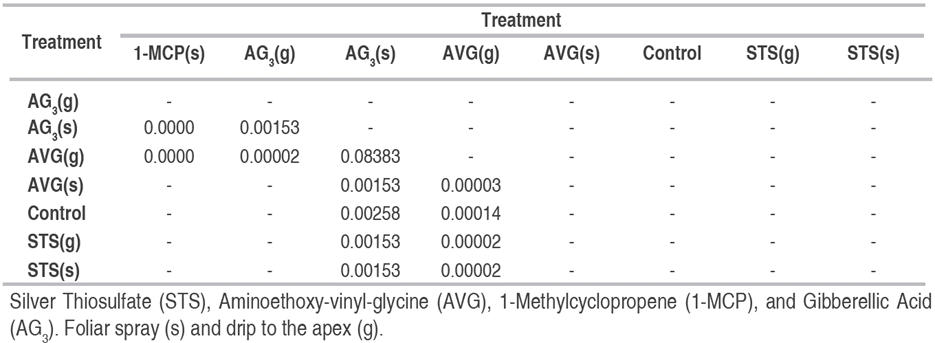
Figure 1 shows the number of male flowers produced by implementing sexual reversion methods compared to the control treatment without reversion. In this sense, the cannabis plants in the control treatment presented female flowers in their entirety, which indicates that the female character was preserved when cuttings taken from female mother plants were used. The plants treated with reversion agents, AG3 and AVG treatments, produced male flowers due to their action on the reproductive physiology of the species.
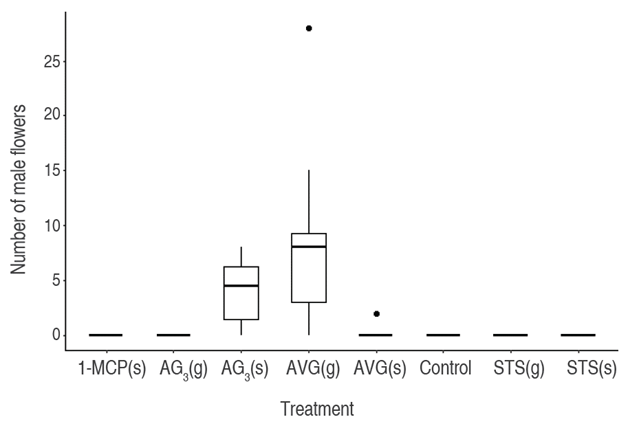
Figure 1 Boxplot of the number of male flowers in response to the treatments. Silver thiosulfate (STS), Aminoethoxy-vinyl-glycine (AVG), 1-Methylcyclopropene (1-MCP), Gibberellic acid (AG3). Foliar spray (s) and drip to the apex (g).
The difference between successful sexual reversal treatments and those that did not differentiate between male and female flowers is clear. The treatments with AVG(g) and AG3(s) showed positive results in the number of male flowers, with a total of 132 and 32 male flowers per treatment, respectively. Regarding plants treated with AVG(s), only one differed by two male flowers; however, no statistically significant difference was observed between AVG(g) and AG3(s) treatments (P=0.08383).
The first symptom observed in plants treated with AVG(g) was the yellowing and discoloration of the green pigments in the leaf apex where the compound was applied, without evidence of compromising the meristem (Figure 2A), unlike the treated plants with AVG(s), which did not present this symptom.

Figure 2 A) Visible symptoms of AVG(g); B) Production of male flowers at 100 µg of AVG; C) Male and female flower buds in plants treated with AG3(s); D) Morphological differences associated with the application of AG3; E) Viable pollen obtained from reverted plants; and F) Male flowers (reduced males).
Between 25 and 30 days after applying the treatments, the formation of the first male flower nodes was observed. With 100 µg of AVG(g), the plants presented only male flowers on the apex where the application was made directly; however, some female flowers produced from the plant apex were observed at the base of the inflorescence. These observations agree with those reported by Mohan Ram and Sett (1982b), who noticed that 75 µg of AVG was sufficient to reverse female-to-male flowering in 100% of the treated plants. Unlike the results obtained in this work, those authors indicate that concentrations of 100 µg of AVG produced abscission of the leaves and inhibition of new nodes. In this study, it was not observed at that concentration of AVG(g), since the plants showed satisfactory results in sexual reversion (Figure 2B).
In plants treated with gibberellins, between the first and second week after GA3 application, a reduction of internodes length, width, and number of leaflets, and curling of the tips were observed besides the apex damage in the plants (Figures 2C, D). However, it should be noted that despite the affectation of the foliar apex, two plants treated with AG3(s) produced flowers at the end of the experiment, initially male flowers, followed by the differentiation of female flowers in the apical shoots (Figure 2C). Male flowers in the plants treated with AG3(s) appeared late, between one and two weeks after the formation of flowers in the other treatments. These results are similar to those obtained by Sarath and Mohan Ram (1979). They determined that 50 µg of AG3 was sufficient to induce sexual reversion in female cannabis plants and that 100 µg of AG3 only induced flowering in 50% of the treated plants, with viable pollen grains within their anthers. Likewise, Mohan Ram and Sett (1982b) reported that 75 µg of AVG applied to female Cannabis plants induces reduced males with 69% pollen viability, which could germinate between 30 and 35 min after being submerged in a sucrose solution medium.
Regarding 1-MCP -although Lubell and Brand (2018) propose evaluating this substance as a possible agent for sexual reversal in female cannabis plants- its ability to occupy ethylene receptors in cannabis plants is justified. The results in this study, this treatment under the application method did not induce the production of male flowers, nor were atypical symptoms observed in the plants concerning the control treatment.
Furthermore, none of the plants treated with the STS solution under the methods (spray or micro drip) at the evaluated concentrations expressed male flowers in female cannabis plants effectively. Contrary to that reported by Mohan Ram and Sett (1982a) and Sarath and Mohan Ram (1979), who indicate that the use of 100 µg of the silver ion (AgNO3) and 50 µg of the STS compound (Ag(S2O3)) was sufficient to reverse more than 80% of the flowers in 100% of treated female Cannabis plants. In addition, Lubell and Brand (2018) state that 3 mM of STS induces between 75 and 100% of the production of male flowers in female cannabis plants. Despite the non-inductive effect of STS for sexual reversion observed in this work, it should be noted that STS is the most used method by cannabis growers to induce the production of male flowers in feminized plants.
Pollen viability
Since only three treatments (AVG(g), AVG(s), and AG3(s)) led to the production of male flowers, the analysis for the pollen viability variable was done by comparing the percentage of viable pollen among the three treatments: those that presented a reddish coloration were considered viable (Figure 2E), and those without coloration, non-viable. The non-parametric Kruskal-Wallis’ test (P=0.0003611) shows a difference between treatments. The Wilcoxon signed-rank test did not show significant differences between the AVG(g) and AG3(s) treatments (P=0.59657), but there was a difference between these two treatments with the AVG(s).
The maximum percentage of pollen viability (96.11%) was observed in reverted male flowers produced by applying AG3(s), followed by 51.05% viability in pollen obtained from reverted flowers with AVG(g), and 6.25% viability using AVG(s) (Figure 3).
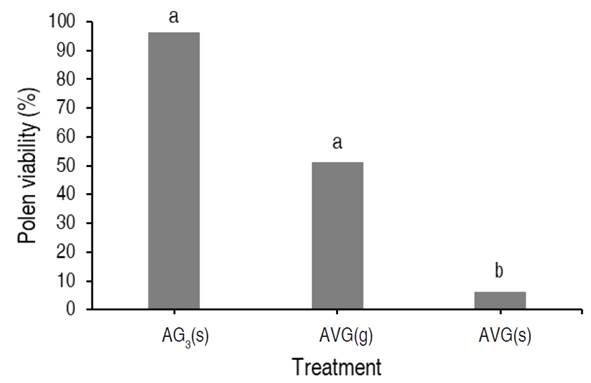
Figure 3 Percentage of pollen viability in male flower from successful reversion treatments. Aminoethoxy-vinyl-glycine (AVG) and Gibberellic acid (AG3). Foliar spray (s) and drip to the apex (g). Different letters between treatments indicate significant differences according to the non-parametric Wilcoxon signed-rank test (P<0.05).
Plant height
After applying the sexual reversal treatments, a variation in plant height was observed between the moment of flower induction and the end of the experiment. The height data analyzed under the non-parametric Kruskal-Wallis' test showed differences between treatments (chi-squared=80.288, df=7, P-value=1.203x10-14). The Wilcoxon signed-rank test shows a difference between all the treatments and the gibberellic acid treatment for this variable -AG3(g) and AG3(s), and even between these two treatments (Table 3).
Table 3 Wilcoxon signed-rank test for the plant height variable in response to sex reversal treatments.
The plants treated with AG3(s) elongated their internodes in the main and secondary branches, unlike the treatment with AG3(g), where this symptom was more marked in the central ápex (Figure 4). In both treatments, morphological changes were previously described. The plants treated with AG3(g) presented chlorosis followed by necrosis of the central apex and growth detection, unlike the treatment with AG3(s), which produced flowers despite reactive apical damage. Similar observations are reported by Sarath and Mohan Ram (1979), where plants treated with AG3 showed elongation of the stems.
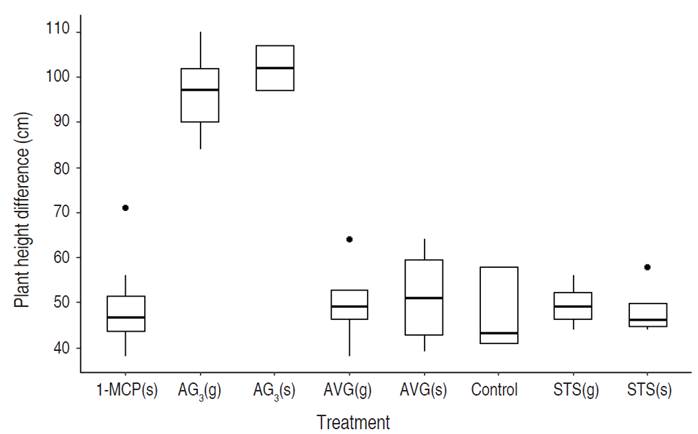
Figure 4 Boxplot of the plant's height in response to the treatments. Silver thiosulfate (STS), Aminoethoxy-vinyl-glycine (AVG), 1-Methylcyclopropene (1-MCP), and Gibberellic acid (AG3). (s) Foliar spray and (g) Drip to the apex.
Several factors influence the floral production of plants, especially in angiosperms, e.g., environmental variations, plant age, and metabolic and epigenetic pathways; they play an essential role during meristem differentiation (Feng et al. 2020). The first stimulus for sexual differentiation in cannabis plants occurs with age. In this sense, Moliterni et al. (2004) found that in plants from sexual seed, the differentiation of vegetative to reproductive meristems can be observed from the fourth internode on, when plants produce single flowers between internodes. Since the cuttings were taken from adult plants, the results obtained in this work showed that the plants formed single flowers between the internodes. Flowering presented after two weeks after changing the photoperiod to a 12/12 ratio presented morphological changes in the architecture of the branches, as reported by Spitzer-Rimon et al. (2019). They indicated that from the beginning of the short days, a severe shortening is observed among the phytomers and there were changes in the size and number of leaflets.
Gerashchenkov and Rozhnova (2013) propose that floral morphogenesis is determined by factors that hierarchically trigger internal plant signals. Results in the genetic coding of protein synthesis and some phytohormones found in homeostasis are strongly influenced by environmental factors. The Cucurbitaceae family showed that unisexual flowers that develop individually in each internode keep ethylene reduced in the meristematic apices, thus favoring the production of male flowers. Likewise, the floral meristems produce deficient levels of ethylene, thus maintaining the androgynous condition. In contrast, the short days, low temperatures, and high levels of ethylene in the meristematic and floral apices induce the production of female flowers, considering that ethylene is a critical hormone for sexual differentiation in this species (Feng et al. 2020; Martínez and Jamilena 2021).
In the case of cannabis, a similar mechanism can occur in the synthesis and signaling of ethylene since applying agents that inhibit the biosynthesis of this hormone, such as AVG or STS and AgNO3, induces the production of male flowers in this species. Different studies reported that applying Silver Thiosulfate (STS) in concentrations of 3 mM favored the production of male cannabis flowers from female plants under short-day conditions (Lubell and Brand 2018). Moreover, Mohan Ram and Sett (1982a) found that 75 µg of AgNO3 was necessary to inhibit the action of ethylene in the production of only male flowers. Mohan Ram and Sett (1982b) found that the application in the apex of 75 µg of AVG induces male flowers in this species. Likewise, applying ethylene-releasing agents such as Ethephon in concentrations of 1,920 mg L-1 to cannabis plants subjected to sexual reversal with AgNO3 or CoCl2 could reverse the masculinizing effect of these compounds, thus emitting female flowers again (Mohan Ram and Sett 1982c).
Mohan Ram and Sett (1982a) evaluated the silver ion AgNO3 and the STS complex to form male flowers in female cannabis plants. The authors propose a classification of the types of flowers found in the branches of treated plants. The reduced male's category coincides with the results found in this study, since the male flowers mainly presented from three to four stamens (Figure 2F); likewise, the pollen in these flowers was not easily shed by the anthers or dispersed by the wind, as occurs naturally in flowers from male plants. Similar observations are reported by Lubell and Brand (2018), who report that pollen from female cannabis plants subjected to sexual reversal treatments with STS applications is not as abundant and its natural release is not as effective. Regarding the germination percentages observed in this work, these results agree with Sarath and Mohan Ram (1979), who experimented with female cannabis plants treated with 100 µg of AG3 and male flowers with viable pollen. Likewise, Mohan Ram and Sett (1982b) reported that 75 µg of AVG applied to female Cannabis plants reduced the production of male flowers with viable pollen, with germination of 69% between 30 and 35 min after being immersed in a sucrose solution medium.
CONCLUSIONS
The AVG and AG3 compounds are effective in sexual reversal and enable the production of male flowers from female cannabis plants. They can reverse the feminizing effects in plants and induce male flowers with viable pollen, thus becoming an alternative that can be included in genetic breeding programs for this species.

