










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActualidades Biológicas
Print version ISSN 0304-3584
Actu Biol vol.29 no.86 Medellín Jan./June 2007
ARTÍCULOS IV CONGRESO COLOMBIANO DE BOTÁNICA
ANATOMÍA DE HOJA Y TALLO DE UN MORFOTIPO DE IRESINE DIFFUSA H. Y B. EX WILLD. (AMARANTHACEAE) DEL VOLCÁN GALERAS (NARIÑO), COLOMBIA
LEAF AND STEM'S ANATOMY OF AN IRESINE DIFFUSA H. Y B. EX WILLD. (AMARANTHACEAE) MORPHOTYPE FROM THE GALERA'S VOLCANO (NARIÑO), COLOMBIA
Carlos A. Agudelo1; Wilton Valencia2; Gerardo A. Torres3; Lyda P. Mosquera4; Manuel Castrillón5
1 Programa de Biología, Centro de Investigación en Biodiversidad y Biotecnología (CIBUQ), Universidad del Quindío. Armenia (Quindío), Colombia y Universidad del Valle. Cali (Valle del Cauca), Colombia. agudelohenao@yahoo.com.
2 Universidad del Valle. Cali (Valle del Cauca), Colombia. wilvalen@gmail.com.
3 Unidad de Microscopía Electrónica, Museo de Historia Natural, Universidad del Cauca. Popayán (Cauca), Colombia. gator@unicauca.edu.co.
4 Unidad de Microscopía Electrónica, Museo de Historia Natural, Universidad del Cauca. Popayán (Cauca), Colombia. lmosquera@unicauca.edu.co.
5 Programa de Ingeniería de Sistemas, Universidad del Cauca. Popayán (Cauca), Colombia. castrillon@unicauca.edu.co.
Resumen
Se presenta por primera vez una caracterización anatómica de hojas y tallos de un morfotipo de Iresine diffusa H. y B. ex Willd. (Amaranthaceae) que crece en las pendientes del volcán Galeras, municipio de Pasto, Departamento de Nariño, Colombia. Las muestras fueron fijadas, incluidas y cortadas y las placas medidas en microscopio óptico. La descripción anatómica procede de una muestra de cinco individuos, de los cuales se hicieron cortes de la hoja en la margen y en la vena principal y del tallo transversal y longitudinalmente. El corte de la hoja permitió la identificación de la cutícula, la epidermis superior uniestratificada y tricomas simples, cortos, no ramificados y uniseriados. El parénquima en empalizada es uniestratificado, con células cilíndricas, estrechas o amplias. El parénquima lagunar posee varias capas de células, espacios intercelulares, células con drusas y haces tipo Kranz. En la vena principal el colénquima es de tipo angular, posee 3–6 haces vasculares. En el tallo el tejido parenquimático forma un anillo, tiene células con drusas, haces vasculares de tipo colateral abierto, esclerénquima en casquetes de fibras extraxilares de tipo perivascular y xilema pluriestratificado, con tráqueas: anuladas, helicadas y punteadas con punteaduras alternas, también se observaron traqueidas.
Palabras clave: anatomía, Amaranthaceae, Iresine diffusa, hoja, tallo
Abstract
A leaf and stem's anatomic characterization of an Iresine diffusa H. & B. ex Wild. (Amaranthaceae) morphotype is presented for the first time. It grows up on the Galeras Volcano slopes, Pasto's town, Nariño's deparment, Colombia. The samples were fixed, included and cut and the laminas were measured in optical microscope. The anatomical description originated in a five individuals' sample, in which sections were made in leaf's margin and main vein and from the transverse and longitudinal stem. The leave's section allowed the identification of cuticle, uniestratificate adaxial epidermis and simple, short, unbranched and uniseriate trichomes. The palisade parenchyma is uniestratificate, with cylinder narrow or wide cells. The lagunar parenchyma has several layers of cell, intercellular spaces, cells with druses and bundle Kranz's type. In the main vein the collenchyma is of angular type, it has 3–6 vascular bundle. In the stem, the parenchyma tissue takes form of a ring, it has cells with druses, vascular bundle from collateral opened, sclerenchyma in islands of extraxylary fibers of perivascular type and pluriestratificate xilem, with traqueas: annuled, helicated and pointed with alternate pits. Tracheids were also observed.
Key words: anatomy, Amaranthaceae, Iresine diffusa, leaf, stem
INTRODUCCIÓN
Para la familia Amaranthaceae la clasificación más aceptada actualmente de acuerdo con Müller y Borsch (2005) es la propuesta por Townsend (1993), la cual está basada en Schinz (1934). Este último, derivó su concepto de Bentham y Hooker (1880). Según Townsend (1993) Amaranthaceae es una familia diversa en las regiones neotropicales, tropicales y en el sureste de Asia y Australia. El centro de diversidad de la subfamilia Gomphrenoideae, en la cual se circunscribe Iresine diffusa H. y B. ex Willd., está en Centro América y México, en varios hábitats y altitudes de los Andes, también como en Venezuela y Brasil.
La taxonomía de la especie objeto de este trabajo, Iresine diffusa H. y B. ex Willd., no es clara, como tampoco lo es su nomenclatura, por lo cual fue revisada por Shinners (1962); adicionalmente, Eliasson (1987) en su tratamiento para la Flora de Ecuador la define como 'una especie muy variable y de una sinonimia extensa', además describe dos variedades de ella, a saber: I. diffusa var. diffusa e I. diffusa var. spiculigera. Igualmente, plantea que la variación es considerable en el tamaño de los órganos florales, como también en la presencia y tipo de pubescencia sobre el ráquis y las hojas, además anota que cuando las formas extremas son comparadas, ellas sugieren que son especies diferentes.
Por lo anterior, el presente estudio tiene como objetivo iniciar la caracterización de la anatomía de hojas y tallos de los morfotipos que I. diffusa tiene en Colombia, con el fin de contribuir a la búsqueda de nuevos caracteres que faciliten su taxonomía.
MATERIALES Y MÉTODOS
Se tomaron muestras vivas de uno de los morfotipos que exhibe la especie Iresine diffusa en Colombia, en este caso una hierba postrada tipo bejuco, de 1–2 m de altura, que crece en las faldas del volcán Galeras en un rango altitudinal que oscila entre 2.700 y 2.900 m. Se colectaron muestras de hojas y tallos de cinco individuos tomados aleatoriamente, de las hojas se extrajeron tres muestras a nivel de la mar gen y tres de la vena principal, así como tres muestras del tallo para cortes transversal y longitudinal respectivamente.
Las muestras fueron llevadas al laboratorio de microscopía electrónica de la Universidad del Cauca en donde fueron sometidas al protocolo de Gonzáles–Santander (1968) para la preparación de muestras para Microscopia Óptica de Alta Resolución (M. O. A. R.), de manera como que se narra a continuación.
Fijación con glutaraldehído al 0,2 M en BP a pH 7,4 por 48 h, lavado del tejido en buffer (dos veces) por 15 min, posfijación en OsO4 por 2 h, lavado con buffer (dos veces) durante 15 min. Luego la deshidratación en etanol al 25% (dos cambios) por 10 min, al 35% (dos cambios) por 10 min, al 50% (dos cambios) por 10 min, al 75% (dos cambios) durante 10 min, al 95% (tres cambios) por 30 min, finalmente en etanol puro (tres cambios) por 30 min. Seguidamente el aclaramiento en etanol:acetona (3:1) por 10 min, luego en proporción 1:1 por 10 min, luego 1:3 por 10 min y finalmente la inmersión en acetona pura por 20 min. A continuación la preimbibición en resina Spurr:acetona (1:3) por 1 h., luego 1:1 por 1 h, pasa a 3:1 durante 1 h., al final en resina Spurr por 18 h. Las muestras se ubicaron en moldes y se orientaron de acuerdo al tipo de corte que se les practicaría y se incluyeron definitivamente en resina pura, la cual se polimeriza a 65 ºC durante 72 h.
Obtenidos los bloques se cortaron en un ultramicrótomo Ultracut Leica con cuchilla de vidrio, se realizaron cortes de 2 m (micras), los cuales se tiñeron con azul de toluidina, se fijaron permanentemente en porta y cubreobjetos mediante resina, se observaron en un microscopio óptico de alta resolución Nikon Microphot, se capturaron las imágenes en cámara digital y se realizó la caracterización morfométrica, tanto cualitativa como cuantitativa, de los tejidos y células mediante el Sistema de Análisis de Imágenes Leica QWIN. Los datos fueron procesados en Excell (versión 2003) y los resultados de las mediciones que se incluyen en la descripción corresponde a rangos y promedios de las células más grandes y más pequeñas localizadas en cada tejido de cada placa, asimismo, los datos de los tejidos son promedios de mediciones realizadas en las zonas más amplias y más estrechas.
Ejemplares de referencia: El estudio histológico se basó en colecciones de C. A Agudelo depositadas en el Herbario de la Universidad del Valle (CUVC) y de la Universidad del Quindío (HUQ) e identificados con los números de colección 5060, 5061, 5062, 5063 y 5064. Igualmente, de los ejemplares estudiados han quedado dos colecciones, una de más de 150 micropreparados permanentes y otra de más de 200 imágenes digitales.
RESULTADOS
Anatomía lámina foliar. El corte transversal de la hoja a nivel de la margen (figura 1) permitió la identificación de la cutícula, la cual es gruesa y se halla sobre la epidermis superior, también sobre ésta se observaron tricomas simples, cortos, no ramificados y uniseriados (figura 2). La epidermis próxima a la haz está conformada por una sola capa de células, las cuales son de forma variable, ya que generalmente son rectangulares a redondeadas pero también ovaladas a redondeadas, el tamaño de las células varía de 82,36–11,17 m de longitud y de 28,3–9,3 m de ancho (figura 1).
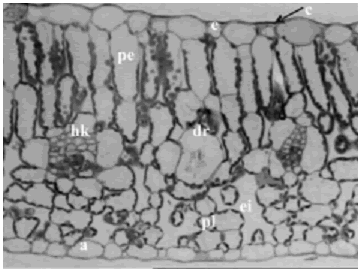
Figura 1. Iresine diffusa. Corte transversal de hoja a nivel de la margen a 16X, se observa la cutícula (c), la epidermis adaxial (e), parénquima en empalizada (pe) con cloroplastos, parénquima lagunar (pl), haz vascular tipo kranz (hk), espacio intercelular (ei), drusa (dr) y la epidermis abaxial (a). Ejemplar de referencia C. A. Agudelo 5061 (CUVC)
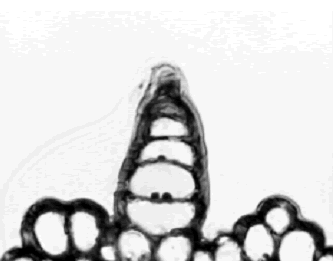
Figura 2. Iresine difusa. Tricoma epidérmico simple, uniseriado, no ramificado y corto en corte transversal de hoja a 40X. Ejemplar de referencia C. A. Agudelo 5074 (CUVC)
El parénquima en empalizada o clorofílico consta generalmente de una capa de células, pero también pueden ser dos, dichas células miden 106–26,02 m de longitud por 27,04–12,87 m de ancho, su forma es cilíndrica, pueden ser estrechas o amplias o en algunos casos son bastoniformes, sin espacios intercelulares, poseen cloroplastos abundantes y cerca de las paredes celulares, estas últimas son delgadas (figura 1).
El parénquima lagunar tiene un ancho total que puede oscilar entre 180,13–118,74 m, en dicho espacio se encuentran de 3 a 7 capas de células sin una organización específica, las células miden de 59,19–8,57 m de longitud por 23,54–8,57 m de ancho, son de forma variable, ya que en general son amorfas, pero también hay cuadradas, rectangulares o redondeadas, tienen paredes no engrosadas y con algunos nutrientes de reserva, el tejido presenta abundantes y grandes espacios intercelulares, además puede poseer células de 70,49–52,53 m de longitud por 56,82–39,54 m de ancho con cristales tipo drusas que alcanzan un tamaño entre 56,24–28,03 m de longitud por 48,27–1411 m de ancho. En este tejido se visualizan grandes cámaras estomáticas (figura 3), amorfas y de tamaño muy variable, ya que pueden ocupar hasta 4 capas de células del tejido lagunar o incluso llegar hasta la región abaxial de las células clorofílicas y miden entre 78,28–50,51 m de longitud.
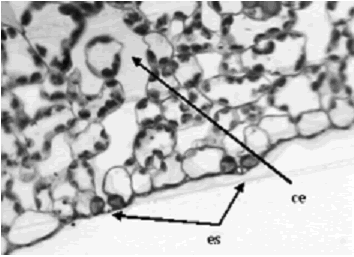
Figura 3. Iresine diffusa. Estoma (es) y su cámara estomática (ce) en corte transversal de hoja a 32X. Ejemplar de referencia C. A. Agudelo 5074 (CUVC)
Inmersos en el tejido lagunar se pueden encontrar haces vasculares con típica anatomía Kranz y haces vasculares correspondientes a venas pequeñas. El haz tipo Kranz tiene un longitud total de 108,8–93,48 m y 76,21–88,83 m de ancho y está conformado por 6 a 11 células grandes que originan la vaina, dichas células son parenquimáticas, de forma variable, ya que van desde redondas a cuadradas, rectangulares o amorfas, además se nota una tendencia a que las células laterales sean más grandes y largas que las de los polos, que son redondeadas y más pequeñas. En el interior de la vaina se observó dos grupos de células, las del xilema en posición adaxial y las de floema abaxialmente (figura 1). Finalmente, se observó la epidermis abaxial conformada por una sola capa de células, cada una de las cuales mide de 42,1–9,57 m de longitud por 18,18–6,77 m de ancho, son de forma rectangular a redondeada, sobre todo las más pequeñas tienden a ser esféricas, también presentan estomas (figura 3) de 35,33–20,3 m de longitud aproximadamente y tricomas similares a los ya descritos.
La vena principal. En el corte transversal se identificaron en su orden la cutícula, la epidermis, tanto adaxial como abaxial, constituida por una sola capa de células, de tamaño entre 26,21–5,02 m de longitud y de 16,9–7,1 m de ancho, de forma redondeada, paredes delgadas y tricomas similares a los mencionados.
A continuación se observó el colénquima de tipo angular (figura 4), el cual existe en dos zonas, una alargada junto a la epidermis inferior (región abaxial) conformada generalmente por dos capas de células y otra corta y engrosada contigua a la epidermis superior en el abultamiento que origina el nervio medio (región adaxial), en este último sitio puede poseer de 2 a 4 capas de células, el ancho total de este tejido junto a la epidermis inferior, zona en la cual es más constante su grosor, es de 74,09–28,09 m, las células del colénquima miden de 36,51–10,6 m de longitud por 32,82–9,73 m de ancho, poseen paredes engrosadas, son de forma redondeada, sin espacios intercelulares visibles y no hay presencia de cristales.
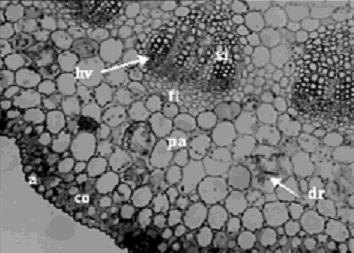
Figura 4. Iresine difusa. Corte transversal de hoja a nivel de la vena principal a 10X, se observa la epidermis abaxial (a), colénquima (co), parénquima (pa), drusa (dr), haz vascular (hv), floema (fl) y xilema (xi). Ejemplar de referencia C. A. Agudelo 5063 (CUVC)
En la región adaxial del nervio se halla una capa de células que corresponde al tejido de empalizada similar al descrito en el corte transversal de la lámina, pero cuando el área que ocupa el nervio forma una protuberancia cuadrangular sobresaliente, las células de tal tejido se observan menos organizadas, se disponen oblicuamente y su forma puede pasar de cilíndricas a ensanchadas centralmente. En algunos casos, dependiendo del desarrollo de los tejidos y de los haces vasculares, se puede observar que el tejido lagunar continúa y está presente en la zona media del nervio principal con una o dos capas de células, o a veces ninguna.
Luego, se encuentra el parénquima lagunar, este tejido posee varias capa de células, el ancho total del tejido es de 250,7–164,83 m, sus células miden de 106,11–19,5 m de longitud por 73,48–16,5 m de ancho, son generalmente redondeadas, pero también hay cuadrangulares, amorfas y pentagonales, posee espacios intercelulares pequeños y presencia de células redondeadas portadoras de drusas. La célula que contiene una drusa puede medir de 91,29–66,7 m de longitud por 73,17–57,63 m de ancho y la drusa, que es de oxalato de calcio (Lozada, s. f.), alcanza 74,79–59,85 m de longitud por 64,84–40,94 m de ancho. El parénquima se localiza tanto en posición adaxial como abaxial con respecto a los haces vasculares, incluso puede rodearlos completamente, o rodear cada haz de forma individual o rodear grupos de haces.
Posteriormente se observó la presencia de 3–6 haces vasculares formando un arco cóncavo–convexo, abierto, o en ocasiones se distribuyen formando una figura elipsoide o incluso se observó un solo haz. Cada haz tiene un tamaño variable, la longitud total oscila de 266,63–213,22 m y el ancho total de 252,71–175,18 m, los haces pueden verse individualmente o de forma grupal, en esta última disposición se forma una banda continua. La constitución del haz incluye varias bandas, la primera de parénquima floemático que mide 63,82–36,54 m de ancho, la segunda de floema que mide 60,24–32,33 m de ancho y posee células generalmente redondeadas, pero también hay cuadrangulares a rectangulares y la tercera de xilema que mide 146,96–91,71 m de ancho con células redondeadas y en algunos casos rectangulares (figura 4).
Anatomía caulinar, sección transversal. La epidermis tiene una sola capa de células, cada una de 29,94–9,15 m de longitud y 90,28–7,42 m de ancho y de forma redondeada, sobre ella se diferencia una gruesa cutícula. Dado que el tallo tiende a ser rectangular el colénquima, que es tipo angular, se distribuye de forma grupal, especialmente en las zonas abultadas angulares, es un tejido de 2 a 6 capas de células y un grosor total de 141,61–14,31 m, cada célula mide de 35,98–14,12 m de longitud y 27,79–8,74 m de ancho, posee paredes engrosadas, su forma es generalmente redondeada, aunque también puede ser ovaladas y no se diferencian espacios intercelulares (figura 5).
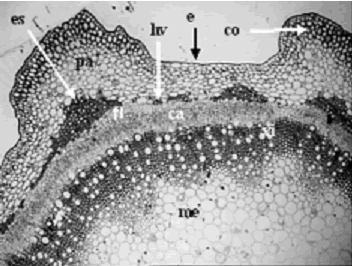
Figura 5. Iresine difusa. Corte transversal de tallo a 4X, se observa la epidermis adaxial (e), colénquima (co), parénquima (pa), anillo de haces vasculares (hv), esclerénquima (es), floema (fl), cambium (ca), xilema (xi) y médula (me). Ejemplar de referencia C. A. Agudelo 5063 (CUVC)
A continuación se observó la presencia del tejido parenquimático, el cual forma un anillo alrededor del tallo, tiene de 5 a 10 capas de células, con un grosor total de 276,1–32,33 m, cada célula mide de 86,33–15,86 m de longitud y 52,1–9,14 m de ancho, las células pueden ser redondeadas, ovaladas o en ocasiones rectangulares. También se encuentran células conteniendo drusas, así como abundantes espacios intercelulares pequeños y de forma triangular, tomando el aspecto de un tejido de tipo lagunar.
Seguidamente se encuentra un anillo de haces vasculares de tipo colateral abierto, los cuales están conformados por grupos celulares de esclerénquima, bandas de floema y de xilema. Un haz vascular completo tiene forma ovalada, una longitud de 810,334–307,16 m y un ancho de 508,65–204,53 m. Las células de esclerénquima se agrupan en casquetes, son fibras extraxilares de tipo perivascular de acuerdo con Paniagua et al. (1997) y sus grupos miden de 474,28–69,04 m de longitud y 128,65–19,62 m de ancho, reúnen pocas células (2) a muchas, dichas células son cuadrangulares a hexagonales o redondeadas a ovaladas, poseen paredes más engrosadas que las del colénquima y están separadas por células parenquimáticas, además poseen pequeños espacios intercelulares. La banda de floema o de tejido criboso tiene 416,01–64,2 m de ancho, en la cual se puede separar el protofloema (zona adaxial) del metafloema (zona abaxial), las células son cuadrangulares, rectangulares, redondeadas u ovadas y la banda de xilema alcanza de 414,21–161,65 m, posee células claramente diferencias de protoxilema (zona abaxial) y metaxilema (zona adaxial), las células son cuadrangulares a hexagonales y redondeadas a ovadas, además poseen en algunos casos 1–4 tílides. El liber está separado del xilema por 2–3 capas de células rectangulares que conforman el cambium vascular (figura 5).
Finalmente, en la parte central se localiza la médula (figura 6), la cual mide en total 1622,06–484,84 m de diámetro, posee células redondas a ovaladas de 148,7–22,33 m de longitud y 130,11–19,4 m de ancho, presenta en ocasiones gránulos nutritivos en su interior y pequeños espacios intercelulares de forma triangular.
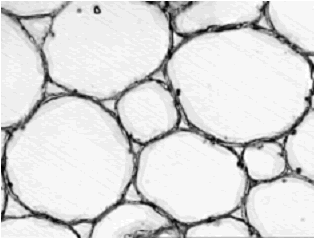
Figura 6. Iresine difusa. Células medulares en corte transversal de tallo a 32X. Ejemplar de referencia C. A. Agudelo 5070 (CUVC)
Sección longitudinal. En este tipo de cortes el tallo presenta la cutícula y la epidermis uniestratificada, cuyas células miden 84,61–19,06 m de longitud y 21,34–12,61 m de ancho y son de forma rectangular. El colénquima, es un tejido angular que se encuentra debajo de la epidermis, constan de 2 a 5 capas de células dispuestas horizontalmente, tiene un ancho total de 114,46–41,71 m, se caracteriza por sus células engrosadas que miden 195,76–46,72 m de longitud por 30,07–20,37 m de ancho y su forma varía de rectangular a elíptica, contienen en su interior gránulos nutritivos, no se observan cristales, ni espacios intercelulares (figura 7).
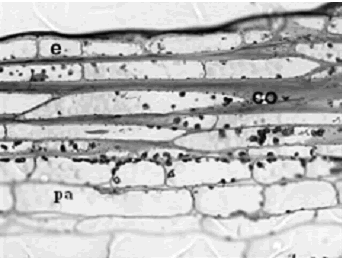
Figura 7. Iresine diffusa. Corte longitudinal de tallo a 20X, se observa la epidermis adaxial (e), colénquima (co), y el parénquima (pa). Ejemplar de referencia C. A. Agudelo 5064 (CUVC)
El tejido parenquimático (figura 7), es pluriestratificado, con hasta 10 capas de células organizadas verticalmente que cubren un espacio de 323,01–137,74 m, sus células miden 164,9–22,11 m de longitud y 41,99–11,17 m de ancho y son de forma rectangular a cuadradas o en ocasiones cónicas, presenta células parenquimáticas conteniendo drusas y también poseen gránulos en su interior.
A continuación se diferencian los tejidos que conforman los haces vasculares, comenzando por el esclerénquima en la región adaxial, éste se caracteriza por presentar las paredes más engrosadas de todos los tejidos, ocupa un espacio que oscila entre 355,02–75,76 m de ancho y sus células en vista longitudinal se visualizan elipsoides, además contie
nen gránulos nutritivos. Luego se observa el floema, el cual tiene un ancho total que varía entre 144,53–109 m de ancho, es un tejido pluriestratificado organizado verticalmente y conformado por células rectangulares. El xilema es pluriestratificado, todo el tejido mide de 378,58–95,06 m de ancho, presenta distintos tipos de vasos o tráqueas (como los descritos por Esau, 1977), a saber: anuladas (figura 8), helicadas y punteada con punteaduras alternas. También se observaron células de forma alargada, elipsoide y de extremos cónicos y oblicuos que corresponden a traqueidas.
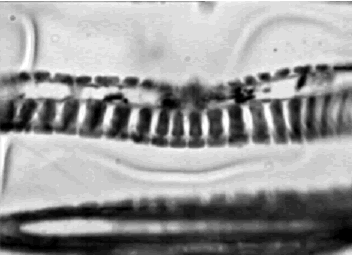
Figura 8. Iresine difusa. Tráquea anulada en corte longitudinal de tallo a 64X. Ejemplar de referencia C. A. Agudelo 5065 (CUVC)
Finalmente, está la médula, es pluriestratificada, tiene un diámetro total que oscila entre 1366,69–549,02 m, posee células de tamaño variable que miden entre 103,81–20,94 m de longitud por 69,84–15,67 m de ancho, son rectangulares a cuadradas o redondeadas.
DISCUSIÓN
Con este estudio se establece la presencia de haces vasculares tipo Kranz en el mesófilo de las hojas, especialmente en el parénquima lagunar en la especie I. diffusa. Entre las dicotiledóneas, las familias Amaranthaceae, Aizoaceae, Chenopodiaceae, Asteraceae, Euphorbiaceae, Nyctaginaceae, Portulacaceae y Zygophyllaceae tienen especies que poseen tales haces vasculares (Carolin et al., 1978), además describió el síndrome Kranz encontrado en hojas del género Gomphrena L., y asociaron su ultra–estructura con la fotosíntesis C4.
Hasta la fecha no se conocen evaluaciones de la estructura y organización de las células y tejidos en I. diffusa, por lo cual esta publicación es la primera para el género, sin embargo en la familia Amaranthaceae sí hay trabajos anatómicos como los de Lozada (s. f.) a nivel de la hoja en Gomphrena celiosioides Mart. y las investigaciones de Ferreira et al. (2003) en especies de Amaranthus L. y en Alternanthera tenella Colla, Costea y DeMason (2001), entre otros.
Lozada (s. f.) encontró en G. celosioides en el mesófilo que el tejido en empalizada está compuesto de una sola capa de células, también reportó en el corte longitudinal la presencia de tejido lacunoso con dos capas celulares de elementos de formas irregulares. En I. diffusa lo más general es que el tejido en empalizada también tenga una sola capa de células, pero pueden existir dos y en el tejido lagunar es mayor el número, ya que pueden encontrarse de 3 a 7 capas de células. En ambas especies también se determinó la presencia de células conteniendo drusas de oxalato de calcio en el mesófilo, pero en I. diffusa se halló adicionalmente los mismos cristales en el parénquima del tallo.
Fank–de–Carvalho y Graciano (2005) hallaron en G. arborescens L. f. dos tipos de tricomas, mesófilo bilateral y tejido clorenquimático dispuesto radialmente a la vaina parenquimática que envuelve los haces vasculares, mientras que aquí se encontró un solo tipo de tricomas, mesófilo igualmente bilateral y el clorénquima en ocasiones radial a la vaina, pero no siempre.
Se halló una cutícula gruesa en las hoja de I. diffusa en ambas caras, por lo cual tiene similaridades, tanto con especies de Amaranthus como de Alternanthera Forsskal., ya que Ferreira et al. (2003) reportaron lo mismo en la cara adaxial, para Amaranthus deflexus L. y A. spinosus L., mientras que en Alternanthera tenella, la cutícula es gruesa en ambas caras.
El estudio mostró uniformidad en la histología del morfotipo de I. diffusa presente en las faldas del volcán Galeras en Nariño, ya que no se observaron diferencias en los tipos de tejidos identificados de cada uno de los individuos, ni en la organización y disposición de los mismos, tampoco en las formas y tamaños de las células. El tipo y tamaño de las drusas observadas y la clase de tricomas fue igualmente similar, así como la forma y distribución del esclerénquima.
Un hecho que resultó interesante fue el hallazgo de bandas de cambium sucesivas en I. diffusa, lo cual había sido reportado para Amaranthaceae por Costea y DeMason (2001), Esau (1965), Metcalfe (1979) y Viana (1993) entre otros. Esto es lo que algunos autores han denominado 'crecimiento anómalo' o presencia de anillos sucesivos en tallos de la familia con crecimiento secundario. Rajput y Rao (1999) encontraron en Pupalia lappacea (L.) Juss., anillos sucesivos de xilema secundario y que el desarrollo de elementos que conducen nutrientes por el xilema y el floema fue restringido al sector fascicular de la banda continua que forma el cambium.
De este trabajo se concluye que caracteres anatómicos como el número de capas de células del tejido en empalizada de la lámina foliar, el grosor de la cutícula de la hoja, el número de capas de haces que se forman en el tallo y el tipo de tricomas presentes en la epidermis, entre otros podrían facilitar la diferenciación de especies de Iresine de las de otros géneros de la familia como Gomphrena y Amaranthus.
Por los anteriores argumentos, esta caracterización anatómica de uno de los morfotipos de I. diffusa se muestra como una herramienta útil para enfrentar problemas taxonómicos, abre nuevas opciones de trabajo y se espera que al caracterizar los otros morfotipos que existen en Colombia (en Cundinamarca, Antioquia, Quindío, etc.) y hacer comparaciones entre ellos surjan alternativas para el manejo a nivel intraespecífico.
AGRADECIMIENTOS
Los autores expresan su especial agradecimiento a la Universidad del Cauca, a la directora del Herbario PSO de la Universidad de Nariño, profesora Aida Elena Baca y a la bióloga Carola Lara Jiménez por el apoyo para el trabajo de campo; a la Universidad del Valle, Facultad de Ciencias Naturales y Exactas, sección Postgrados por el apoyo logístico y económico; así como también a la Universidad del Quindío, en especial al Programa de Biología y al Centro de Estudios e Investigaciones en Biodiversidad–CIBUQ.
REFERENCIAS
1. Bentham G, Hooker JD. 1880. Genera Plantarum, 3(2):20–43. [ Links ]
2. Carolin RC, Jacobs SWL, Vesk M. 1978. Kranz cells and mesophyll in the Chenopodiales. Australian Journal of Botany, 26:683–698. [ Links ]
3. Costea M, DeMason DA. 2001. Stem morphology and anatomy in Amaranthus L. (Amaranthaceae) _ Taxonomic significance. Journal of the Torrey Botanical Society, 128.3:254–281. [ Links ]
4. Eliasson UH. 1987. Amaranthaceae. Pp. 1–138. En: Harling G, Andersson L (eds.). Flora of Ecuador, 28. Göteborg, Suecia. [ Links ]
5. Esau K. 1965. Plant anatomy. 2nd ed. John Wiley and Sons. New York. [ Links ]
6. Esau K. 1977. Anatomy of seed plants. Editorial John Wiley and Sons. U. S. A. [ Links ]
7. Fank–de–Carvalho SM, Graciano–Ribeiro D. 2005. Arquitetura, anatomia e histoquímica das folhas de Gomphrena arborescens L. f. (Amaranthaceae). Acta Botanica Brasilera, 19(2):377–390. [ Links ]
8. Ferreira EA, Procópio SO, Silva EAM, Silva AA, Rufino RJN. 2003. Estudos anatômicos de folhas de espécies de plantas daninhas de grande ocorrência no Brasil. IV – Amaranthus deflexus, Amaranthus spinosus, Alternanthera tenella e Euphorbia heterophylla. Planta Daninha, 21(2):263–271. [ Links ]
9. Gonzáles SR. 1968. Técnicas de microscopía electrónica en Biología. Editorial Aguilar. Madrid. [ Links ]
10. Lozada GM. (s. f.). Estudo anatômico do eixo vegetativo de plantas daninhas que ocorrem em minas gerais. I. anatomia foliar de Gomphrena celosioides Mart. (Amaranthaceae). Universidade Federal de Lavras/(UFLA). Minas Gerais, Brasil. [ Links ]
11. Metcalfe CR, Chalk L. 1979. Anatomy of the Dicotyledons _ Systematic anatomy of the leaf and stem. V.1. 2 ed. Oxford University Press. London. [ Links ]
12. Müller K, Borsch T. 2005. Phylogenetics of Amaranthaceae based on matK/trnK sequence data _ Evidence from Parsimony, Likelihood, and Bayesian analyses. Annals Missouri Botanical Garden, 92:66–102. [ Links ]
13. Paniagua R, Nistal M, Sesma P, Álvarez M, Fraile B, Anadón R, Sáez FJ, de Miguel MP. 1997. Citología e Histología vegetal y animal. Editorial McGraw–Hill–Interamericana. España. [ Links ]
14. Rajput SS, Rao KS. 1999. Structural and developmental studies on cambial variant in Pupalia lappacea (Amaranthaceae). Annals Botani Fennici, 36:137–141. [ Links ]
15. Schinz H. 1934. Amaranthaceae. 16c:7–85. En: Engler A, Prantl K (eds.). Die Naturlishen Pflanzenfamilien. Germany. [ Links ]
16. Shinners Ll H. 1962. Illegitimacy of the names Iresine Celosia L., I. celosioides L., and I. paniculata (L.) Kuntze (Amaranthaceae). Taxon, 11(4):141–142. [ Links ]
17. Townsend CC. 1993. Amaranthaceae. Pp. 70–91. En: Kubitzki K (ed.). Families and genera of vascular plants. Springer–Verlag. Berlin. [ Links ]
18. Viana V. 1993. Contribuição ao estudo anatômico do eixo vegetativo de Amaranthus viridis L. Arquivos Jardim Botânico Rio de Janeiro, 31:15–70. [ Links ]
Recibido: agosto 2007; aceptado: noviembre 2007.

