










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkActualidades Biológicas
versión impresa ISSN 0304-3584
Actu Biol v.29 n.87 Medellín jul./dic. 2007
ARTÍCULOS DE INVESTIGACIÓN
ESTRUCTURA Y COMPOSICIÓN DEL ENSAMBLAJE DE MURCIÉLAGOS (CHIROPTERA) ASOCIADO A UN CAFETAL CON SOMBRÍO EN LA MESA DE LOS SANTOS (SANTANDER), COLOMBIA
STRUCTURE AND COMPOSITION OF THE BAT ASSEMBLAGE ASSOCIATED TO A SHADE COFFEE PLANTATION IN MESA DE LOS SANTOS (SANTANDER), COLOMBIA
David A. Ortegón–Martínez1; Jairo Pérez–Torres2
1Pontificia Universidad Javeriana. Carrera 65, # 94A–64. Bogotá, Colombia. Direción electrónica: <david.ortegon@javeriana.edu.co>
2Laboratorio de Ecología Funcional, Unidad de Ecología y Sistemática (UNESIS). Pontificia Universidad Javeriana. Carrera. 7, # 43–82. Bogotá, Colombia. Dirección electrónica: <jaiperez@javeriana.edu.co>
Resumen
Se describe la estructura y composición del ensamblaje de murciélagos presente en el cafetal con sombrío de la hacienda 'El Roble' (Santander), Colombia, a 1.650 m de altitud, entre septiembre y noviembre de 2003. Debido a la heterogeneidad espacial de la zona de estudio se escogió un muestreo por conglomerados. Se muestrearon 5 zonas correspondientes a 3 tipos de hábitats utilizando redes de niebla colocadas a dos diferentes alturas. Se capturó un total de 147 individuos pertenecientes a 11 especies de las familias Phyllostomidae (10 especies) y Vespertilionidae (1 especie). Las especies más abundantes fueron Sturnira lilium, Artibeus lituratus y Carollia brevicauda. Se utilizaron valores tróficos y valores de importancia para describir la estructura trófica del ensamblaje, que mostró un predominio de los frugívoros sedentarios con respecto a los frugívoros nómadas, insectívoros de follaje e insectívoros aéreos. Al comparar las proporciones de los gremios tróficos en los diferentes hábitats mediante los radios de probabilidad (Odds ratio), cada gremio presentó una mayor probabilidad de ser encontrado en un tipo de hábitat diferente al ocupado por los otros gremios tróficos. En el caso de los frugívoros sedentarios y nómadas esto se explicó en términos de sus estrategias de alimentación. La estructura del ensamblaje encontrada, junto con la presencia de Phyllostomus discolor (Phyllostominae) nos permiten sugerir que el cafetal con sombrío de la hacienda 'El Roble' constituye una importante fuente de alimento y refugio para las especies asociadas a este monocultivo.
Palabras clave: Cafetales con sombrío, composición, Chiroptera, Colombia, estructura
Abstract
The structure and composition of the bat assemblage on the shade coffee plantation at 'El Roble' farm (Santander), Colombia, located at 1.650 m of altitude was described between September and November 2003. Due to the spatial heterogeneity of the area a cluster sampling design was chosen. Five sampling zones or clusters corresponding to 3 habitat types were sampled using mist nets at 2 different heights. All captured bats were measured, aged and its reproductive condition was determined. Fecal samples were collected. A total of 147 individuals from 11 species belonging to Phyllostomidae (10 species) and Vespertilionidae (1 species) families were captured. The most abundant species were Sturnira lilium, Artibeus lituratus, and Carollia brevicauda, this dominance pattern along with the presence of Phyllostomus discolor (Phyllostominae) suggested, that the shade coffee plantation at 'El Roble' farm, is in similar conservation conditions to primary and secondary forests with a low intervention degree. Trophic values and importance values were used to describe the trophic structure of the assemblage wich was dominated by sedentary frugivores, followed by foliage gleaner insectivores, nomadic frugivores and aerial insectivores, respectively. When proportions of the trophic groups among habitats were compared by using the Odds ratios technique, each trophic group presented a higher probability to be captured in a different habitat type than the other trophic groups. In the particular case of the sedentary and nomadic frugivores this was explained in terms of its feeding strategies.
Key words: Chiroptera, Colombia, composition, shade coffee plantations, structure
INTRODUCCIÓN
En Latinoamérica, el café es el agro–ecosistema más representativo en elevaciones medias y cubre un área total de 5,5 millones de hectáreas cultivadas (FAO, 2006). En Colombia, aunque en los últimos 10 años el área cultivada de café ha disminuido a una tasa del 5,5% anual, aún constituye aproximadamente el 36% (560.000 ha) de los cultivos permanentes (FAO, 2006). Adicionalmente el café se cultiva en elevaciones medias (500–2.000 m) de relieves montañosos y laderas volcánicas de las cordilleras oriental y central que se caracterizan por presentar altos niveles de biodiversidad y endemismo (Perfecto y Armbretch, 2003). En estos lugares, las prácticas de cultivo tecnificado de café implican la deforestación y es únicamente en las áreas de cultivo de café tradicional donde se ha mantenido la cobertura arbórea original (Perfecto y Armbretch, 2003). El café es producido mediante una amplia variedad de técnicas, que se clasifican de acuerdo con el grado de conservación del estrato arbóreo de la siguiente manera: a) rusticano, b) policultivo tradicional, c) policultivo comercial, d) monocultivo con sombrío y e) monocultivo sin sombrío (Fuentes–Flores, 1979). Las cuatro primeras mantienen la heterogeneidad espacial al conservar diferentes estratos de vegetación en el cultivo. En el rusticano las plantas arbustivas originales son substituidas por plantas de café. En el policultivo tradicional, además de sustituir el estrato arbustivo del bosque por plantas de café, se siembran otro tipo de árboles de explotación económica (Greenberg y Rice, 2000). En el policultivo comercial, los árboles que formaban la cobertura original son reemplazados por leguminosas y cítricos. En el monocultivo bajo sombra la cobertura arbórea es reemplazada por una o dos especies de árboles de sombrío (Greenberg y Rice, 2000). En el monocultivo sin sombrío, la cobertura arbórea es eliminada y solamente se mantienen las plantas de café tolerantes al sol; transformándose de un sistema agro–forestal a un sistema totalmente agrícola (Greenberg y Rice, 2000). Para el año 2000, Colombia contaba con 260.000 ha de café cultivado bajo sombrío (López, 2002). Varios autores han propuesto que el café con sombrío además de ser viable económicamente, causa efectos de menor impacto sobre la biodiversidad que los cultivos más tecnificados (Fuentes–Flores, 1979; Numa et al., 2005; Perfecto et al., 1996; Perfecto y Armbretch, 2003; Pineda et al., 2005) en la medida que mantiene una alta complejidad estructural en su interior (Perfecto et al., 1996). En muchas regiones del país, los bosques de montaña han sufrido un fuerte proceso de deterioro y pérdida. En algunos casos la cobertura arbórea de la cual dependen muchas especies animales es mantenida principalmente por los cafetales con sombrío, e incluso pueden constituir la única cobertura disponible para especies sensibles a la pérdida de hábitat. Sin embargo, se desconoce la diversidad de fauna y flora asociada a estos sistemas, por lo que es importante describir los componentes faunísticos que persisten en ellos.
Este estudio se centra particularmente en los murciélagos, que con 178 especies reportadas, son el orden de mamíferos con mayor número de especies en Colombia (Alberico et al., 2000), y cumplen diferentes papeles funcionales desde consumidores primarios hasta depredadores de aves, murciélagos, roedores y otros vertebrados pequeños (Fenton et al., 1992, Medellín 2003). Además algunas especies de quirópteros son sensibles a la degradación del hábitat por lo que pueden servir como indicadores biológicos del grado de disturbio de los ecosistemas (Altringham, 1996; Fenton et al., 1992; Medellín et al., 2000; Pineda et al., 2005). Se describe la riqueza, diversidad y estructura trófica del ensamblaje de murciélagos presente en un cafetal con sombrío en la zona de la Mesa de Los Santos (Santander), Colombia. En esta zona prácticamente la única cobertura arbórea presente es la de este sistema de producción, y este trabajo contribuye a conocer qué especies de murciélagos persisten en este monocultivo.
MATERIALES Y MÉTODOS
Área de estudio. Este trabajo se llevó a cabo en la hacienda 'El Roble' ubicada en la vereda Los Cacaos, municipio de Piedecuesta y vereda La Granja, municipio de Los Santos, departamento de Santander, Colombia, a 1.650 m de altitud, en una zona de vida definida como Bosque Húmedo Pre–montano (CORPES, 1991) (figura 1). La hacienda tiene un área total de 334 ha de las cuáles 220 ha son cultivos de café con sombrío. Se encuentra rodeada por un paisaje altamente intervenido y degradado debido a la constante extracción de madera y al creciente número de fincas de recreación y clubes.

Figura 1. Localización geográfica del área de estudio en la hacienda 'El Roble' en la Mesa de los Santos (Santander, Colombia) [tomado de Peraza (2003)]
En la hacienda se encuentran tres diferentes tipos de cultivo: rusticano, policultivo tradicional y policultivo comercial, en diferentes estados de desarrollo formando cinco hábitats estructuralmente distintos, distribuidos de forma irregular y conformando una matriz heterogénea (figura 2).
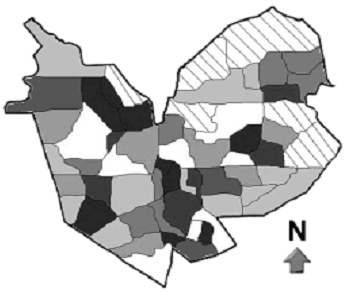
Figura 2. Distribución de los hábitats estructurales en la hacienda 'El Roble' en la Mesa de los Santos (Santander), Colombia, cada tono de color representa un tipo de hábitat, el área rayada corresponde a lotes donde no hay cultivos de café
En la región se presentan dos estaciones lluviosas al año (abril–mayo y septiembre–octubre). El muestreo se realizó entre septiembre y noviembre del 2003 cubriendo así la temporada de lluvias y la transición a la temporada seca del segundo semestre del año.
Debido a la heterogeneidad en cuanto a formas, tamaños y tipos de cultivo de los lotes de la hacienda (figura 2) se realizó un muestreo por conglomerados (Martínez, 2002). Se dividió la hacienda en cinco zonas de muestreo (conglomerados) de 64 ha, cada uno de las cuáles incluía diferentes tipos de hábitats en su interior.
En cada conglomerado se ubicó un transecto lineal de 240 m de longitud, cuyo centro coincidía con el del conglomerado y su dirección fue determinada al azar. Se establecieron cinco puntos de muestreo por transecto con una separación aproximada de 60 m. En cada uno de ellos se colocaron dos redes de niebla (2,6 x 6 m con ojo de 2 cm2) a dos diferentes alturas (0–4 m y > 4 m), con el fin de aumentar el éxito de captura de los murciélagos (Kunz y Kurta, 1988).
Se hizo un muestreo de manera consecutiva en los cinco conglomerados alternándolos cada cuatro noches. El orden de muestreo se estableció aleatoriamente al inicio del mes de septiembre y se mantuvo igual durante los siguientes meses de muestreo. Las redes permanecieron abiertas toda la noche (18:00–06:00) y se revisaron cada media hora. Teniendo en cuenta que la probabilidad de captura se puede ver afectada por la intensidad lumínica, ya que podría incrementar la probabilidad de detectar las redes (Hecker y Brigham, 1999; Kunz y Kurta, 1988; Negraeff y Brigham, 1995), los muestreos se programaron para ser realizados evitando la fase de luna llena.
Los individuos capturados fueron identificados hasta nivel de especie, para lo cual se elaboró una clave de campo a partir de las claves de Emmons y Feer (1996), Fernández et al. (1988), Medellín et al. (1997), Muñoz (2001) y, Timm y LaVal (1998). Se colectó un macho y una hembra de cada especie para hacer una colección de referencia la cual fue depositada en la colección de mamíferos del Museo Javeriano de Historia Natural (PUJ–MUJ). La corroboración de la identidad taxonómica de los ejemplares colectados y de los criterios de identificación en campo se hizo con la ayuda del Dr. Michael Alberico (q.e.p.d.). Los individuos capturados se colocaron en bolsas de tela por aproximadamente una hora para obtener muestras de materia fecal; las cuáles fueron preservadas en alcohol al 70% para su posterior análisis. Los individuos que no eran colectados fueron marcados con un anillo metálico numerado en el antebrazo y posteriormente liberados.
Se realizó una caracterización estructural de la vegetación en cada uno de los cinco tipos de hábitats de la hacienda, mediante un levantamiento de 0,1 ha (50 x 20 m) siguiendo la metodología de muestreo de plantas leñosas propuesta por Gentry (1982). Se registró la especie, altura total (AA), diámetro mayor de copa (DM), diámetro menor de copa (Dm), DAP y cobertura del estrato arbóreo (C). Para el estrato arbustivo se registró sólo la especie y la altura de cada individuo (Aa). Para obtener un estimativo relativo del tamaño de los árboles se utilizó la ecuación:
TA = [(DM x Dm)/DAP] x AA
Posteriormente, se calculó un índice de complejidad estructural de la vegetación para cada hábitat mediante la fórmula:
CE = (TA x C x Aa)/100
Entre más alto es el valor del índice mayor es la complejidad bien sea por el incremento del tamaño de los árboles, la cobertura de la vegetación o la altura del estrato arbustivo.
Análisis de información. Como indicador de la abundancia relativa de las especies se cálculo del éxito de captura (Pérez–Torres 2004) en cada conglomerado usando la fórmula:
E = (N/m x h) x 100.
Donde: N = número de individuos capturados; m = metros cuadrados (m2) de red; h = número de horas de muestreo.
La composición fue cuantificada mediante el índice de riqueza específica alfa (á) de Fisher (Moreno, 2000). Se generaron funciones de acumulación de especies considerando las semanas como unidades de respuesta. Para eliminar la influencia del orden en el que se adicionaron las semanas, la muestra fue aleatorizada 1.000 veces utilizando el programa EstimateS Software (Colwell, 2000). Con el objeto de calcular el número esperado de especies, estas curvas se ajustaron a los modelos de dependencia lineal y ecuación de Clench a través del procedimiento de regresión no lineal del programa Statistica (Statsoft, 2000) utilizando las ecuaciones:
E(s) = (a/b) • (1 – e –bx) Modelo de Dependencia Lineal (Dep. lineal)
E(s) = (ax)/(1+bx) Ecuación de Clench
Donde: a = constante que representa la tasa de incremento de la lista al inicio de la colección; b = constante que representa la pendiente de la curva; x = número acumulado de muestras; E(s) = número de especies esperadas en x muestras.
Ambos modelos predicen la riqueza específica para una muestra de x individuos como el valor a través del cual una curva de acumulación de especies alcanza la asíntota, la cual se calcula como la relación entre las constantes a/b (Soberón y Llorente, 1993).
Se describió la estructura del ensamblaje considerando la dominancia y la equidad, para lo cual se utilizó el índice de Simpson y el índice de Shannon respectivamente (Magurran, 2003). Con el propósito de conocer si los murciélagos capturados en toda el área de muestreo pertenecían al mismo ensamblaje, se comparó el éxito de captura de los cinco conglomerados mediante un análisis de varianza para medidas repetidas (Roldán, 1995). Se comprobó la normalidad de los datos aplicando la prueba de Shapiro–Wilks y la homogeneidad de varianzas mediante la prueba de Hartley (Martínez y Martínez, 1997). Para los análisis estadísticos se utilizó el programa Statistix versión 8.0 (Analytical Software, 2003). Se calculó la riqueza, la dominancia y la equidad para cada uno de los conglomerados y se compararon estableciendo los intervalos de confianza del 95% utilizando el programa PAST versión 1.21 (Hammer et al., 2004).
La estructura trófica se describió a partir del análisis de las muestras fecales siguiendo a Rivas–Pava et al. (1996). Se registró la presencia de semillas, insectos, material vegetal y material indeterminado en cada muestra. Se calculó la frecuencia relativa de cada uno de estos componentes alimenticios en la muestra total de cada especie y se determinó su contribución a cada categoría trófica calculando los valores tróficos (VT) con la fórmula (Soriano 1983, Willig 1986, Rivas–Pava et al. 1996):
VT = Xi/N.
Donde: Xi = número total de muestras fecales de la especie i con determinado componente alimenticio; N = número total de muestras fecales analizadas de la especie i.
Para describir la estructura trófica del ensamblaje de murciélagos, se calculó el valor de importancia (VI) (Smith y Genoways, 1974):
VI = [(Σ VTi )/S] x 100.
Donde: VTi = valor trófico de la especie i en cada categoría; S = número de especies en la muestra.
Se estableció si la probabilidad de encontrar cada una de las categorías tróficas variaba según el tipo de hábitat, mediante los Odds ratios o radios de probabilidad (Montgomery y Runger, 1996) calculados de la siguiente forma:
Odd = p/1–p.
Donde p es la probabilidad de ocurrencia de una categoría trófica en un hábitat determinado. Si al comparar los Odds ratios de una categoría trófica en dos hábitats el valor obtenido es igual a uno, significa que no existen diferencias en la probabilidad de encontrar esa categoría trófica en los dos hábitats. Por el contrario, cualquier valor obtenido diferente de uno, establece una diferencia en la probabilidad de encontrar determinada categoría trófica en los dos hábitats comparados. Como estimador estadístico se calcularon los intervalos de confianza del 95% de los Odds ratios mediante la siguiente fórmula:

Donde Za/2 es el punto crítico superior que corresponde al porcentaje a/2 de la distribución normal estándar (Montgomery y Runger, 1996)
Dada la ubicación aleatoria de los transectos, en los hábitats 1 y 4 no quedaron instaladas redes de niebla. Por esta razón las comparaciones solo incluyen los hábitats 2, 3 y 5.
RESULTADOS
Vegetación. El hábitat 1 presentó la menor complejidad estructural. Carecía de estrato arbóreo y el estrato arbustivo estaba exclusivamente representado por platas de café con un promedio de altura de 2,38 m.
El hábitat 2 presentó un estrato arbóreo cuya altura promedio fue de 27 m. Presentaban copas amplias superpuestas proporcionando una cobertura entre 81–86% al estrato arbustivo. El estrato arbustivo estaba conformado por plantas de café de más de 12 años de edad, con un promedio de altura de 2 m. Las ramas de las plantas de café se encontraban superpuestas dificultando el paso entre los surcos.
El hábitat 3 contaba con un estrato arbóreo de 35,2 m de altura en promedio con copas amplias y superpuestas, proporcionando una cobertura entre el 91–98%. El estrato arbustivo estaba conformado por plantas de café con altura promedio de 2,72 m. En este estrato se encontraba también plátano con una altura promedio de 6,5 m y sembrados cada 10 m.
El hábitat 4 presentaba un estrato arbóreo cuya altura promedio era de 8,69 m y una sobreposición de las copas baja, proporcionando una cobertura al estrato arbustivo entre el 48–59%. Las plantas de café del estrato arbustivo tenían en promedio 1,4 m de altura, intercalada con plátano con altura promedio de 3,45 m y sembradas cada 6 m.
El hábitat 5 presentaba árboles que tenían en promedio 13,6 m de altura y proporcionando una cobertura entre el 58–70%. El estrato arbustivo presentaba plantas de café que tenían 1,65 m de altura en promedio, sembradas cada 1,5 m y plátano con altura promedio de 4,1 m sembrados cada 6 m.
El hábitat 3 presentó la mayor complejidad estructural (CE), seguido de los hábitats 2, 5 y 4; respectivamente. En la tabla 1 se presenta la información de la caracterización de la vegetación en cada uno de los cinco tipos de hábitats encontrados en la hacienda 'El Roble'.

Tabla 1. Promedios de las variables estructurales de los hábitats de la hacienda 'El Roble' en la Mesa de los Santos (Santander), Colombia (entre paréntesis aparecen los valores los máximos y mínimos; NA = no aplica, AA = altura del árbol; DM = diámetro mayor de la copa; Dm = diámetro menor de la copa; DAP = diámetro a la altura del pecho; C = cobertura; Aa = altura del arbusto; CE = complejidad estructural)
Ensamblaje de murciélagos. Dado que al comparar el éxito de captura, índice de riqueza específica alfa de Fisher, índice de Simpson e índice de Shannon entre los cinco conglomerados no se encontraron diferencias significativas; los individuos capturados fueron considerados como elementos del mismo ensamblaje de murciélagos en el área de estudio. Por esta razón los análisis posteriores se hicieron considerando la hacienda 'El Roble' como una única unidad de muestreo.
El éxito de captura de la hacienda 'El Roble' presentó valores muy variables (figura 3). En el mes de septiembre correspondiente a la temporada de lluvias alcanzó su máximo valor. En octubre, mes en el que finalizó la temporada de lluvias, descendió hasta su valor más bajo y en noviembre (correspondiente a la transición hacia la temporada seca) presentó un valor intermedio entre los dos anteriores. Este patrón es similar al encontrado en algunos ecosistemas neotropicales y se debe a que en la temporada de lluvias hay una mayor disponibilidad de frutos y algunos grupos de insectos que en la temporada seca. Esto se refleja en una reducción de la abundancia y la diversidad de murciélagos al final de la temporada de lluvias y en menor medida durante la transición a la temporada seca (Bonaccorso, 1979; Heithaus et al., 1975).
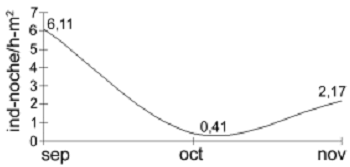
Figura 3. Éxito de captura de murciélagos en la hacienda 'El Roble' en la Mesa de los Santos (Santander), Colombia en el periodo de septiembre a noviembre de 2003 (ind = individuos; h = hora)
Se capturaron 147 murciélagos pertenecientes a once especies, diez de ellas de la familia Phyllostomidae y una de la familia Vespertilionidae (tabla 2) después de 501 horas de muestreo y un éxito total de captura de 1,81 individuos–noche/hora–m2. Las capturas, horas, noches de muestreo, esfuerzo de muestreo y éxito de captura por conglomerado se muestran en la tabla 3.

Tabla 2. Número de individuos capturados por especie en la hacienda 'El Roble' en la Mesa de los Santos (Santander), Colombia

Tabla 3. Valores totales de noches, horas, esfuerzo y éxito de captura por conglomerado en la hacienda 'El Roble' en la Mesa de los Santos (Santander), Colombia
La especie dominante fue Sturnira lilium, seguida por Carollia brevicauda y Artibeus lituratus (tabla 2). Las especies menos abundantes (menos del 5% de la muestra) fueron Glossophaga soricina, Sturnira ludovici, Carollia perspicillata y Glossophaga longirostris y aquellas con abundancias especialmente bajas o inferiores al 1% fueron Anoura geoffroyi y Eptesicus fuscus.
La curva de acumulación de especies (figura 4) se ajustó muy bien al modelo de dependencia lineal (R2 = 0,98) y al modelo de la ecuación de Clench (R2 = 0,98). Según las predicciones del modelo de dependencia lineal, en el muestreo se logró la captura del 100% de las especies esperadas (11 spp.) y según las predicciones de la ecuación de Clench, se logró la captura del 85% de las especies esperadas (14 spp.).

Figura 4. Ajuste de la curva de acumulación de especies (especies observadas) de murciélagos a los modelos de dependencia lineal (Dep. lineal) y ecuación de Clench (especies esperadas), las barras verticales representan intervalos de confianza del 95% de los datos observados en la hacienda 'El Roble' en la Mesa de los Santos (Santander), Colombia
La distribución de abundancias se ajustó al modelo Log–normal (P > 0,05). El valor de riqueza fue a = 2,115 < 2,752 < 2,753, la dominancia fue D = 0,20 < 0,24 < 0,30 y la equidad fue H' = 1,57 < 1,78 < 1,89.
Debido a la baja abundancia dentro de la muestra, los nectarívoros/polinívoros no fueron considerados. La estructura trófica se caracterizó por la dominancia de los frugívoros sedentarios (42%) seguidos por los insectívoros de follaje (27%), los frugívoros nómadas (20%) y los insectívoros aéreos (11%), respectivamente.
Al comparar las proporciones de los gremios tróficos entre los hábitats utilizando los Odds ratios (figura 5), se halló que los frugívoros sedentarios presentaban una mayor probabilidad de ser encontrados en el hábitat 3, que era el que presentaba la mayor complejidad estructural (CE = 286,28). Los insectívoros y los frugívoros nómadas presentaron una mayor probabilidad de ser encontrados en los hábitat 2 (CE = 147,34) y 5 (CE = 75,46), respectivamente.

Figura 5. Odds ratio por hábitat: A. Hábitat 2; B. Hábitat 3; C. Hábitat 5 [FS = frugívoro sedentario; FN = frugívoro nómada; IN = insectívoro; las líneas verticales representan intervalos de confianza del 95%; el eje Y representa el valor de probabilidad de ocurrencia (por debajo del punto de corte en 1) o de no ocurrencia (por encima del punto de corte en 1) de una categoría trófica en un hábitat determinado, si el valor es igual a uno, significa que la probabilidad de encontrar o no encontrarlo es la misma; la altura de cada barra se debe interpretar como el número de veces que es probable o no encontrar un individuo en un hábitat (donde una unidad equivale al 100%)]
DISCUSIÓN
El ajuste de la curva de acumulación de especies a los modelos de dependencia lineal y ecuación de Clench (R2 = 0,98), permite considerar las muestras obtenidas como una representación plausible del ensamblaje de murciélagos presente en el cafetal con sombrío de la hacienda 'El Roble' bajo las condiciones temporales y espaciales en las que se desarrolló este trabajo.
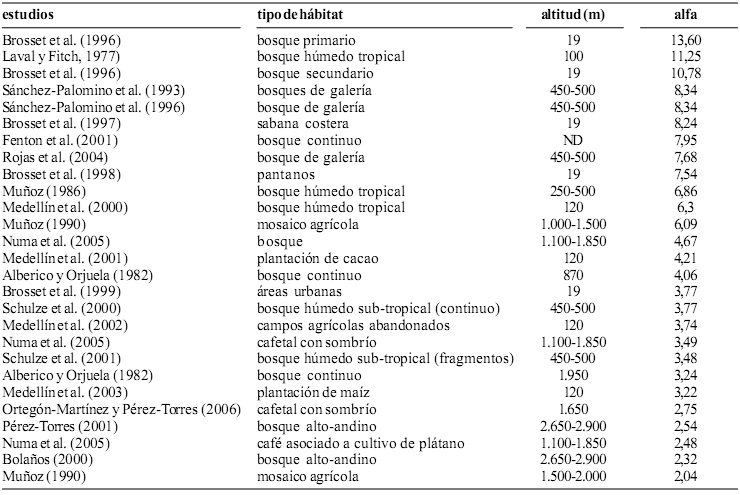
La riqueza específica medida a través del índice alfa de Fisher (a = 2,75) fue menor a la encontrada en la mayoría de los estudios realizados anteriormente en diferentes ecosistemas colombianos de menor altitud (anexo 1). Resultado consistente con la existencia de un patrón de reducción de la riqueza de especies de murciélagos conforme se asciende sobre el nivel del mar planteado por Tamsitt y Valdivieso (1963) y Muñoz (1990), acompañado con el hecho que la mayoría de estos estudios (Medellín et al., 2000; Muñoz, 1986, 1990; Rojas et al., 2004; Sánchez–Palomino et al., 1993) se realizaron incluyendo las dos temporadas climáticas del año y en algunos casos como en el estudio de Fenton et al. (2001) se utilizaron técnicas de captura y registro de la presencia de especies de murciélagos diferentes a las redes de niebla.
La equidad del ensamblaje de murciélagos de la hacienda 'El Roble' (H' = 1,78) fue mayor a la encontrada en ecosistemas agrícolas tecnificados y áreas abiertas, siendo a su vez, un valor similar al encontrado por varios autores en ecosistemas naturales o modificados que a pesar de presentar diferentes grados de intervención se caracterizaron por conservar alta complejidad estructural en su interior (tabla 4).

Tabla 4. Valores del índice de Shannon en estudios realizados por otros autores [entre paréntesis se muestran los intervalos de confianza del 95% del presente estudio en la hacienda 'El Roble' en la Mesa de los Santos (Santander), Colombia]
La distribución log–normal y el valor encontrado para el índice de Simpson (D = 0.241) son el reflejo de un ensamblaje con una dominancia no muy marcada. Hay una especie dominante (9%), varias abundantes (36%) y una gran proporción de especies poco abundantes (54%), patrón similar al observado por otros autores en ensamblajes de murciélagos estudiados con métodos similares (Estrada y Coates–Estrada, 2002; Fleming et al., 1972; LaVal y Fitch, 1977; Pérez–Torres, 2001; Soriano, 1983).
Las especies dominantes en la muestra: Sturnira lilium, Carollia brevicauda y Artibeus lituratus coinciden con las encontradas en otros estudios realizados en ecosistemas de bosque primario, secundario o cafetales con sombrío con bajo grado de intervención antrópica y altamente conservados (Brosset et al., 1996; Medellín et al., 2000; Numa et al., 2005). Esto, acompañado por la presencia de Phyllostomus discolor considerada una especie indicadora del buen estado de conservación de los ecosistemas (Fenton et al., 1992; Galindo–González, 2004; Medellín et al., 2000) permite pensar en el cafetal con sombrío de la hacienda 'El Roble' como un ecosistema que se encuentra en condiciones de conservación comparables con las de ecosistemas de bosque natural.
La alta proporción de especies con abundancias bajas dentro del muestreo: Glossophaga soricina (4,1%), Sturnira ludovici (3,4%), Carollia perspicillata (2%) y Glossophaga longirostris (2%), Anoura geoffroyi (0,7%)y Eptesicus fuscus (0,7%) puede deberse a una combinación entre su baja abundancia en la zona de estudio, falta de esfuerzo de muestreo y a la utilización de redes de niebla como única técnica de captura ya que muchas de ellas han sido señaladas como especies comunes y abundantes en otras localidades cercanas. En el caso de Carollia perspicillata, la hacienda el Roble se encuentra cercana al límite superior de su distribución altitudinal, lo que puede explicar su baja ocurrencia.
La dominancia de especies frugívoras sobre insectívoras en el muestreo, se encuentra acorde con los patrones observados en diferentes localidades neotropicales (Bolaños, 2000; Rivas–Pava et al., 1996; Schultze et al., 2000; Soriano, 1983, 2000). Esto se explica por la constante oferta alimenticia proporcionada por las plantas de los ecosistemas tropicales (Rivas–Pava et al., 1996; Soriano, 1983), en este caso particular, hay una alta variedad de árboles de sombrío que ofrecen frutos, néctar y polen a los murciélagos frugívoros promoviendo su dominancia en este ecosistema. Es posible que la baja proporción de especies insectívoras dentro de la muestra se deba a la técnica de muestreo utilizada. Las redes de niebla permiten capturar especies que vuelan por debajo del dosel de los árboles y en el caso de los murciélagos insectívoros, la mayoría de las especies vuelan sobre el estrato arbóreo o a alturas superiores a las que se colocaron las redes de niebla en este trabajo (Bernard, 2001; Fleming et al., 1972; Kalko et al., 1996; Sánchez–Palomino et al., 1993).
Dentro del grupo de los frugívoros, la dominancia de los sedentarios frente a los nómadas, puede deberse a que los frugívoros sedentarios se alimentan de árboles con frutos pequeños y de producción continua durante todo el año, siendo característicos de los bosques secundarios o ecosistemas con algún grado de intervención (Galindo–González, 1998; Schultze et al., 2000). En el caso de la hacienda 'El Roble' estos árboles son muy abundantes ya que constituyen el sombrío plantado en los lotes de policultivo comercial y policultivo tradicional. Las especies Piper bogotense (Piperaceae), Erytrina eludis (Fabaceae) y Cecropia sp. (Cecropiaceae), son las principales especies cultivadas en estos lotes. Por otro lado, los frugívoros nómadas se alimentan de frutos de producción masiva y corta disponibilidad temporal, producidos por árboles de mayor tamaño y característicos de bosques primarios (Rivas–Pava et al., 1996; Schultze et al., 2000; Soriano, 2000) los cuáles se encuentran en menor cantidad dentro del área de estudio y están distribuidos principalmente en los lotes de cultivo rusticano, siendo Psidium guajava (Myrtaceae), Cordia alliadora (Boraginaceae) y Solanum sp. (Solanaceae) las especies más comunes dentro de estos lotes.
Los frugívoros sedentarios presentaron mayor probabilidad de ser encontrados en un hábitat con alta complejidad estructural. Lo cual obedece a su tipo de alimentación, caracterizado por presentar itinerarios más o menos fijos de búsqueda
de frutos (Rivas–Pava et al., 1996), junto con la conservación de sitios de percha permanentes y cercanos a la fuente de alimento que les puede evitar la necesidad de desplazamiento por áreas de menor complejidad estructural y mayor riesgo de depredación.
Los frugívoros nómadas por su parte, presentaron mayor probabilidad de ser encontrados en el hábitat con menor complejidad estructural, lo cual es reflejo de sus necesidades de alimentación. Debido a que tanto los árboles con los frutos de los cuales se alimentan como los refugios nocturnos donde consumen los frutos se encuentran más distanciados entre sí en comparación con los de los frugívoros sedentarios (Rivas–Pava et al.,1996), deben cubrir distancias que oscilan entre los 2–10 km cada noche para encontrar su alimento (Galindo–González, 1998; Heithaus, 1982). Esto hace necesario su desplazamiento a lo largo de diferentes tipos de hábitats, incluso en aquellos donde el estrato arbóreo se encuentra poco desarrollado y hay una alta exposición a depredadores pero a través del cual pueden llegar a las zonas donde se alimentan.
En el caso de los insectívoros, por tratarse en su mayoría de especies generalistas que consumen además de insectos, frutos, néctar y material vegetal con excepción de Eptesicus fuscus que es considerada una especie enteramente insectívora (Kurta y Baker, 1990; Muñoz, 2001) no fue posible plantear una explicación con respecto a su distribución diferencial en los hábitats 2, 3 y 5 de la hacienda 'El Roble'. En este caso es necesario hacer un análisis detallado de su dieta para encontrar posibles relaciones con la oferta alimenticia en cada hábitat.
Al analizar el número de individuos totales capturados por hora (figura 6) se observó la existencia de 3 picos de captura bien definidos a lo largo de la noche, el mayor de ellos entre las 22:00–02:00 horas con un total de 70 capturas, seguido por otros dos picos mucho menores, el primero entre las 18:30–20:30 horas con un total de 33 capturas y el segundo entre las 04:00–05:30 horas con 29 capturas en total. Este patrón de captura sugiere que los murciélagos presentes en el cafetal con sombrío de la hacienda 'El Roble' forrajean en su interior y por eso presentan la mayor tasa de captura entre las 22:00–02:00 horas, consideradas por otros autores como de alta actividad de forrajeo en especial para especies frugívoras (Charles–Dominique, 1991; Galindo–González, 1998; Nowak, 1994).
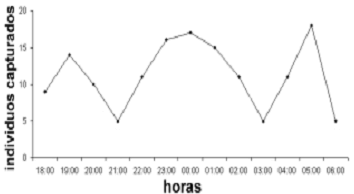
Figura 6. Total de individuos capturados por hora en el cafetal con sombrío de la hacienda 'El Roble' en la Mesa de los Santos (Santander), Colombia entre septiembre y noviembre del 2003.
En conclusión el patrón de dominancia de especies frugívoras encontrado en este trabajo coincide con el de otros estudios realizados en ecosistemas de bosque primario y secundario con bajo grado de intervención antrópica y altamente conservados.
La diversidad del ensamblaje de murciélagos presente en el cafetal con sombrío de la hacienda 'El Roble', descrita en términos de dominancia y equidad, adquirió valores similares a los encontrados por otros autores en hábitats con complejidad estructural alta y bajos niveles de intervención antrópica. Lo anterior resalta la contribución potencial que los cafetales con sombrío ofrecen para mantener la diversidad a nivel regional cuando los hábitats naturales han sido alterados o han desaparecido.
El patrón de captura durante la noche sugiere que el cafetal con sombrío de la hacienda 'El Roble' es un agro–ecosistema de alta importancia para las especies de murciélagos que allí encuentran refugio y alimento. Adquiere mayor importancia si se tiene en cuenta que esta rodeado por un paisaje altamente intervenido y degradado donde la cobertura arbórea ha sido prácticamente eliminada constituyendo uno de los pocos lugares donde especies sensibles a la degradación de su hábitat pueden sobrevivir.
Es importante dedicar especial atención a sistemas productivos como el de la hacienda 'El Roble', ya que pueden convertirse en un modelo que integre la agricultura sostenible, la conservación biológica y la estabilidad económica y social.
AGRADECIMIENTOS
La empresa Café Mesa de Los Santos apoyó económica y logísticamente la fase de campo de este proyecto. El Laboratorio de Ecología Funcional de la Pontificia Universidad Javeriana facilitó los equipos y materiales. Germán Jiménez colaboró en la revisión de los documentos del proyecto. Marco Vinicio Martínez colaboró en la fase de campo. Camilo Peraza fue quién hizo los contactos para acceder a la hacienda. Este trabajo hizo parte del proyecto 'Diversidad de aves y murciélagos en cafetales con sombrío en la hacienda el Roble (Mesa de los Santos, Santander), (ID:000573)' financiado por la Pontificia Universidad Javeriana.
REFERENCIAS
1. Alberico M, Cadena A, Hernández–Camacho J, Muñoz–Saba Y. 2000. Mamíferos (Synapsida: Theria) de Colombia. Biota Colombiana, 1(1):43–75. [ Links ]
2. Altringham JD. 1996. Bats, biology and behavior. Oxford University Press. New York, U. S. A. [ Links ]
3. Analytical Software. 2003. Statistix 8.0 for Windows. WEB: <http://www.analyticalsoftware.com>. [ Links ]
4. Bernard E. 2001. Vertical stratification of bat communities in primary forests of Central Amazon, Brazil. Journal of Tropical Ecology, 17:115–126. [ Links ]
5. Bolaños T. 2000. Dieta y estructura trófica de la comunidad de Chiroptera en cuatro áreas de bosque andino en el sector sur–occidental de la sabana de Bogotá, Colombia. Trabajo de grado. Departamento de Biología. Facultad de Ciencias Pontificia Universidad Javeriana. Bogotá, Colombia. [ Links ]
6. Bonaccorso FJ. 1979. Foraging and reproductive ecology in a Panamanian bat community. Bulletin of the Florida State Museum, Biological Series, 24:359–408. [ Links ]
7. Brosset A, Charles–Dominique P, Cockle A, Cosson J, Masson D. 1996. Bat communities and deforestation in French Guiana. Canadian Journal of Zoology, 74(11):1974–1982. [ Links ]
8. Charles–Dominique P. 1991. Feeding strategy and activity budget of the frugivorous bat Carollia perspicillata (Chiroptera: Phyllostomidae) in French Guiana. Journal of Tropical Ecology, 7:243–256. [ Links ]
9. Colwell RK. 2000. EstimateS 7.0 Statistical estimation of species richness and shared species for samples. Department of Ecology and Evolutionary Biology. University of Connecticut. U. S. A. WEB: <http://viceroy.eeb.uconn.edu/EstimateS> [ Links ]
10. CORPES. 1991. Atlas ambiental del Departamento de Santander. Primera edición. Ingeniería Gráfica. Cali, Colombia. [ Links ]
11. Emmons LH, Feer E. 1996. Neotropical rainforest mammals. A field guide. Second edition. The University of Chicago Press. Chicago, U. S. A. [ Links ]
12. Estrada A, Coates–Estrada R. 2002. Bats in continuous forest, forest fragments and in an agricultural mosaic habitat–island at Los Tuxtlas, México. Biological Conservation, 103:237–245. [ Links ]
13. FAO. 2006. The Agricultural Bulletin Board on Data Collection, Dissemination and Quality of Statistics project. FAO Homepage <http://faostat.fao.org/faostat/collectionsagriculture>. Fecha de consulta: agosto 2 de 2006. [ Links ]
14. Fenton MB, Acharya L, Audet D, Hickey MB, Merriman MK, Obrist D, Byrne M. 1992. Phyllostomid bats (Chiroptera: Phyllostomidae) as indicators of habitat disruption in the neotropics. Biotrópica, 24(3):440–446. [ Links ]
15. Fenton MB, Bernard E, Bouchard S, Hollis L, Jhonston DS, Lausen CL, Ratcliffe JM, Ruskin DK, Taylor JR, Zigouris J. 2001. The bat fauna of Lamanai, Belize: roosts and trophic roles. Journal of Tropical Ecology, 17:511–524. [ Links ]
16. Fernández A, Guerrero R, Lord R, Ochoa J, Ulloa G. 1988. Mamíferos de Venezuela: lista y claves para su identificación. Primera edición. Asociación Venezolana para el Estudio de los Mamíferos. Caracas, Venezuela. [ Links ]
17. Fleming TH, Hooper ET, Wilson DE. 1972. Three Central American bat communities: Structure, reproductive cycles and movement's patterns. Ecology, 53:556–569. [ Links ]
18. Fuentes–Flores R. 1979. Coffee production systems in Mexico. Pp. 60–71. En: de las Salas F (ed.). Workshop on agroforestry systems in Latin America. The Tropical Agronomic Center for Research and Teaching. Turrialba, Costa Rica. [ Links ]
19. Galindo–González J. 1998. Dispersión de semillas por murciélagos: su importancia en la conservación y regeneración del bosque tropical. Acta Zoológica Mexicana, 73:57–74. [ Links ]
20. Galindo–González J. 2004. Clasificación de los murciélagos de la región de los Tuxtlas, Veracruz, respecto a su respuesta a la fragmentación del hábitat. Acta Zoológica Mexicana, 20:239–243. [ Links ]
21. Gentry AH. 1982. Patterns of Neotropical plant diversity. Evolutionary Biology, 15:1– 84. [ Links ]
22. Greenberg R, Rice RA. 2000. Manual de café bajo sombra y biodiversidad en el Perú. Primera edición. Migratory Bird Center Smithsonian Institution. Washington, D. C., U. S. A. [ Links ]
23. Hammer O, Harper DAT, Ryan PD. 2004. PAST: Paleontological statistics version 1.21. University of Oslo. Norway. WEB: <http://www.folk.uio.no/ohammer/past/>. [ Links ]
24. Hecker KR, Brigham RM. 1999. Does moonlight change vertical stratification of activity by forest–dwelling insectivorous bats? Journal of Mammalogy, 80:1196–1201. [ Links ]
25. Heithaus ER, Fleming T, Opler PA. 1975. Foraging patterns and resource utilization in seven species of bats in a seasonal tropical forest. Ecology, 56:841–854. [ Links ]
26. Heithaus ER. 1982. Coevolution between plants and bats. Pp. 327–367. En: Kunz TH (ed.). Ecology of bats. Plenum Press. New York, U. S. A. [ Links ]
27. Kalko EK, Handley Jr CO, Handley D. 1996. Organization, diversity and long–term dynamics of a Neotropical bat community. Pp. 504–555. En: Cody ML, Smallwood JA (eds.). Long–term studies of vertebrate communities. Academic Press. San Diego, U. S. A. [ Links ]
28. Kunz TH, Kurta A. 1988. Capture methods and holding devices. Pp. 1–29. En: Kunz TH (ed.). Ecological and behavioral methods for the study of bats. Smithsonian Institution Press. Washington, D. C., U. S. A. [ Links ]
29. Kurta A, Baker RH. 1990. Eptesicus fuscus. Mammalian Species, 356:1–10. [ Links ]
30. LaVal RK, Fitch HS. 1977. Structure, movements and reproduction in three Costa Rican bat communities. Occasional Papers Museum of Natural History of the University of Kansas, 69:1–28. [ Links ]
31. López W. 2002. Café: Técnica y tradición. Primera edición. Universidad Santiago de Cali. Cali, Colombia. [ Links ]
32. Magurran AE. 2003. Ecological diversity and its measurement. Second edition. Princeton University Press. New York, U. S. A. [ Links ]
33. Martínez C. 2002. Muestreo, algunos métodos y sus aplicaciones prácticas. Décimo primera edición. Interamericana Editores. Bogotá, Colombia. [ Links ]
34. Martínez R, Martínez N. 1997. Diseño de experimentos: Análisis de datos estándar y no estándar. Primera edición. Editora Guadalupe Ltda. Bogotá, Colombia. [ Links ]
35. Medellín RA, Arita HT, Sánchez O. 1997. Identificación de los murciélagos de México, clave de campo. Primera edición. Asociación Mexicana de Mastozoología. México D. F., México. [ Links ]
36. Medellín RA, Equihua M, Amin MA. 2000. Bat diversity and abundance as indicators of disturbance in neotropical rainforests. Conservation Biology, 14(6):1666–1675. [ Links ]
37. Medellín RA. 2003. Diversity and conservation of bats in México: Research, priorities, strategies and actions. Wildlife Society Bulletin, 31(1):87–97. [ Links ]
38. Montgomery DC, Runger GC. 1996. Probabilidad y estadística aplicadas a la ingeniería. Quinta edición. McGraw–Hill–Interamericana Editores. México D. F., México. [ Links ]
39. Moreno CE. 2000. Manual de métodos para medir la biodiversidad. Primera edición. Estado de Hidalgo, México. [ Links ]
40. Muñoz J. 1986. Murciélagos del Parque Natural 'El Refugio' (Antioquia, Colombia). Actualidades Biológicas, 12:66–76. [ Links ]
41. Muñoz J. 1990. Diversidad y hábitos alimenticios de murciélagos en transectos altitudinales a través de la cordillera Central de los Andes en Colombia. Neotropical Fauna and Environment, 1:1–17. [ Links ]
42. Muñoz J. 2001. Los murciélagos de Colombia: Sistemática, distribución, descripción, historia natural y ecología. Primera edición. Editorial Universidad de Antioquia. Medellín, Colombia. [ Links ]
43. Negraeff E, Brigham RM. 1995. The influence of moonlight on the activity of little brown bats (Myotis lucifugus). International Journal of Mammalian Biology, 60:330–336. [ Links ]
44. Nowak RM. 1994. Walker´s bats of the World. Fifth edition. The Johns Hopkins University Press. London, UK. [ Links ]
45. Numa C, Verdú JR, Sánchez–Palomino P. 2005. Phyllostomid bat diversity in a variegated coffee landscape. Biological Conservation, 122:151–158. [ Links ]
46. Pérez–Torres J. 2001. Efecto de la fragmentación sobre la estructura y composición de la comunidad de murciélagos en bosques alto–andinos. Tesis de maestría. Departamento de postgrado. Facultad de Ciencias. Pontificia Universidad Javeriana. Bogotá, Colombia. [ Links ]
47. Pérez–Torres J. 2004. Dinámica del ensamblaje de murciélagos en respuesta a la fragmentación en bosques nublados, un modelo de ecuaciones estructurales. Tesis de Doctorado en Ciencias Biológicas. Departamento de Postgrado. Facultad de Ciencias. Pontificia Universidad Javeriana. Bogotá, Colombia. [ Links ]
48. Perfecto I, Rice RA, Greenberg R, van der Voort ME. 1996. Shade coffee: A disappearing refuge for biodiversity. BioScience, 46(8):598–608. [ Links ]
49. Perfecto I, Armbretch I. 2003. The coffee agroecosystem in the neotropics: Combining ecological and economical goals. Pp. 160–281. En: Vandermeer JH (ed.). Tropical Agroecosystems. CRC Press. Washington, D. C., U. S. A. [ Links ]
50. Pineda E, Moreno C, Escobar F, Halffter G. 2005. Frog, bat and dung beetle diversity in the cloud forest and coffee agroecosystems of Veracruz. Conservation Biology, 19(2):400–410. [ Links ]
51. Rivas–Pava P, Sánchez–Palomino P, Cadena A. 1996. Estructura trófica de la comunidad de quirópteros en bosques de galería de la Serranía de la Macarena (Meta–Colombia). Contributions in Mammalogy: A memorial volume Honoring Dr. J. Knox Jones, Jr. Museum of Texas University. Texas, U. S. A. [ Links ]
52. Rojas AM, Cadena A, Stevenson PR. 2004. Preliminary study of the community at the CIEM, Tinigua National Park, Colombia. Field studies: Fauna and Flora of La Macarena, Colombia, 14:45–56. [ Links ]
53. Roldán JL. 1995. Como elaborar un proyecto de investigación. Primera edición. Editorial Campbell. Murcia, España. [ Links ]
54. Sánchez–Palomino P, Rivas–Pava P, Cadena A. 1993. Composición, abundancia y riqueza de especies de la comunidad de murciélagos en bosques de galería de la Serranía de la Macarena (Meta–Colombia). Caldasia, 17(2):301–312. [ Links ]
55. Schultze MD, Seavy NE, Whitacre DF. 2000. A comparison of the phyllostomid bat assemblages in undisturbed neotropical forest and in forest fragments of a slash and burn farming mosaic in Petén, Guatemala. Biotropica, 32(1):174–184. [ Links ]
56. Smith JD, Genoways HH. 1974. Bats of Margarita Island, Venezuela with zoogeographic comments. Bulletin of Southern California Academy of Science, 73: 64–79. [ Links ]
57. Soberón J, Llorente J. 1993. The use of species accumulation functions for the predictions of species richness. Conservation Biology, 7:480–488. [ Links ]
58. Soriano PJ. 1983. La comunidad de Quirópteros de las selvas nubladas de los andes de Mérida. Patrón reproductivo de los murciélagos frugívoros y las estrategias fenológicas de las plantas. Tesis de Maestría. Universidad de los Andes. Mérida, Venezuela. [ Links ]
59. Soriano PJ. 2000. Functional structure of bat communities in tropical rainforests and andean cloud forests. Ecotropicos, 13(1):1–20. [ Links ]
60. StatSoft. 2000. Statistica for Windows. StatSoft, Inc. WEB: <http://www.statsoft.com>. [ Links ]
61. Tamsitt JR, Valdivieso D. 1963. Records and observations on colombian bats. Journal of Mammalogy, 44:168–180. [ Links ]
62. Timm RM, LaVal R. 1998. A field key to the bats of Costa Rica. Occasional Publication Series of the University of Kansas. Center of Latin American Studies. University of Kansas. Kansas, U. S. A. [ Links ]
63. Willig MR. 1986. Bat community structure in South America: A tenacious chimera. Revista Chilena de Historia Natural, 59:151–168. [ Links ]
Anexo 1. Riqueza especifica de murciélagos encontrada en otros estudios (ND = información no disponible en la publicación)
Recibido: febrero 2007
Aceptado: noviembre 2007

