










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActualidades Biológicas
Print version ISSN 0304-3584
Actu Biol vol.30 no.88 Medellín Jan./June 2008
ARTÍCULOS DE INVESTIGACIÓN
VARIACIÓN ESPACIO–TEMPORAL DE LA ASOCIACIÓN FITOPLANCTÓNICA EN DIFERENTES MOMENTOS DEL PULSO HIDROLÓGICO EN LA CIÉNAGA DE AYAPEL (CÓRDOBA), COLOMBIA
SPATIAL AND TEMPORAL VARIATION OF THE PHYTOPLANKTON ASSEMBLAGE IN DIFFERENT MOMENTS OF HYDROLOGICAL FLOOD IN THE AYAPEL SWAMP (CÓRDOBA), COLOMBIA
Esnedy Hernández–Atilano1; Néstor J. Aguirre2; Jaime A. Palacio3; John J. Ramírez–Restrepo4
1 Grupo de Investigación en Gestión y Modelación Ambiental (GAIA). Sede de Investigaciones Universitarias (SIU), Universidad de Antioquia. A. A. 1226. Medellín (Antioquia), Colombia. esheat@jaibana.udea.edu.co
2 Grupo de Investigación en Gestión y Modelación Ambiental (GAIA). Sede de Investigaciones Universitarias (SIU), Universidad de Antioquia. A. A. 1226. Medellín (Antioquia), Colombia.naguirre@jaibana.udea.edu.co
3 Grupo de Investigación en Gestión y Modelación Ambiental (GAIA). Sede de Investigaciones Universitarias (SIU), Universidad de Antioquia. A. A. 1226. Medellín (Antioquia), Colombia.japalaci@jaibana.udea.edu.co
4 Grupo de Limnología Básica y Experimental (LimnoBasE). Instituto de Biología, Universidad de Antioquia. A. A. 1226. Medellín (Antioquia), Colombia. johnra77@yahoo.com.
Resumen
Esta investigación analizó la asociación fitoplanctónica en el principal cuerpo de agua del sistema cenagoso de Ayapel (Córdoba), Colombia en diferentes períodos del pulso hidrológico. Los resultados obtenidos en seis jornadas de campo entre septiembre de 2004 y junio de 2005 en cinco estaciones de muestreo, mostraron que la estructura de la asociación del fitoplancton y la dinámica física y química varía en relación al pulso hidrológico. En términos generales fueron registrados 37 taxones, de los cuales Cylindrospermopsis raciborskii y Lyngbya cf. limnetica representantes de la división Cyanoprocariota, mostraron las abundancias más importantes con respecto a otros taxones. Las variaciones en la estructura fueron representadas principalmente por la distribución de abundancias y el biovolumen de la asociación, los cuales presentaron mayores valores en las épocas seca y de transición a lluvias.
Palabras clave: fitoplancton, pulso hidrológico, ciénega Ayapel, Colombia
Abstract
This investigation analyzed the variation of the phytoplanktonic assemblage in the main water body of the swamp system of Ayapel (Córdoba), Colombia in different periods from the hydrological pulse. The results in six field periods between September 2004 and June 2005 and in five sampling stations showed that the structure and dynamics of the assemblage and physical and chemical variables vary in relation to the hydrological pulse. In general terms 37 taxa were registered, being Cylindrospermopsis raciborskii and Lyngbya cf. limnetica the most important taxa. The variations in the structure of the assemblage, presented greater in the dry periods and in the transition to rain period.
Key words: phytoplankton, hydrological pulse, s Ayapel (Córdoba), Colombia swamp Ayapel, Colombia
INTRODUCCIÓN
Las ciénagas y las zonas inundables actúan como reguladores de los regímenes hidrológicos de las áreas tributarias, son amortiguadores climáticos, fuente de aprovisionamiento de agua y tienen un alto valor biogénico (Neiff, 1999). En estos ambientes limnéticos existen numerosas formas de vida y entre ellas las algas planctónicas son una de las principales bases de incorporación de carbono en la red ecológica del ambiente limnético (Margalef, 1983).
La llanura aluvial del río San Jorge presenta algunos sistemas lagunares asociados, entre los cuales se destaca la ciénaga de Ayapel. Sin embargo, el conocimiento ecológico de este sistema es incipiente y se basa principalmente en el estudio del manejo integral de la Ciénaga (Ecoestudios y CVS, 1989) y el de impacto ambiental por minería realizado por la Universidad de Antioquia para la Corporación Autónoma de los Valles del Sinú y San Jorge en 1990. En la actualidad se desconoce la dinámica morfológica e hidrológica y los procesos biogénicos asociados al sistema cenagoso de Ayapel. A pesar de que se ha detectado una tendencia progresiva hacia la reducción del contacto hidráulico entre el complejo lagunar y el río San Jorge, no es posible realizar aún predicciones confiables de este proceso (Aguirre et al., 2005). Igualmente se desconoce cómo la productividad biológica del sistema cenagoso representado en parte por la cadena trófica establecida en el sistema sostiene el recurso pesquero.
Este estudio investigó la asociación fitoplanctónica de la ciénaga de Ayapel y sus afluentes en diferentes períodos hidrológicos con el fin de establecer cómo la variación del pulso hidrológico y en consecuencia la dinámica física y química del agua influencia cambios en la estructura y la oferta trófica del fitoplancton en el área de estudio. Para cumplir con este objetivo, se efectuaron seis muestreos en diferentes períodos hidrológicos en cinco estaciones y planteándose como pregunta de investigación central el cómo varía la estructura de la asociación en respuesta a diferentes momentos del puso hidrológico y se hipotetizó que durante las épocas de lluvias la asociación del fitoplancton es homogénea espacialmente, así como también el aumento de nivel reduce las densidades de las comunidades, lo anterior debido a la mayor conectividad entre el sistema cenagoso.
MATERIALES Y MÉTODOS
Descripción del área de estudio. El municipio de Ayapel se encuentra localizado en el extremo oriental del departamento de Córdoba, en límites con los departamentos de Sucre, Bolívar y Antioquia, Colombia, sobre la vertiente oriental del río San Jorge, el sistema hídrico más importante y alrededor del cual gira la actividad económica regional (figura 1).
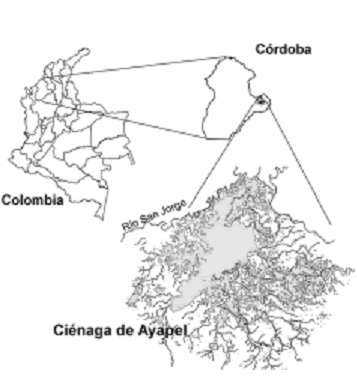
Figura 1. Ubicación de la ciénaga de Ayapel (Córdoba), Colombia
En su llanura aluvial el río San Jorge tiene asociados varios sistemas de ciénagas de gran importancia como San Marcos, San Benito, la Hormiga y Ayapel (Aguirre et al., 2005; IGAC, 1986). La Ciénaga de Ayapel tiene una extensión aproximada de 106,4 km2 en época de aguas altas y posee un sistema hidrológico propio que recoge las precipitaciones de las quebradas de los costados oriental y sur (Quebrada Quebradona, Quebrada Escobilla, Caño Trejos, Caño Muñoz y Caño Don Matías). Su principal nexo hidrológico establece el flujo de intercambio con Río San Jorge por el caño Grande. Como
reguladora de los regímenes hidrológicos de la zona, la ciénaga se alimenta de caudales de creciente del río San Jorge y ocasionalmente del río Cauca. El territorio se ubica en una zona de bosque húmedo tropical, en donde se presenta una época de lluvias de abril a noviembre y una época seca de diciembre a marzo. La precipitación anual fluctúa entre 2.000 y 2.500 mm, la temperatura ambiente alcanza valores mayores a 25 ºC (IGAC, 1986; Aguirre et al., 2005). En el complejo cenagoso de Ayapel fueron establecidas cinco estaciones en el espejo principal del agua (figura 1).
La estación uno (E1), se encuentra ubicada al frente del sitio El Cedro (8º 117' 42,5' N y 75º 08' 12,3' O), zona de influencia de la quebrada Piedras y hace parte de la zona más profunda de la ciénaga. La estación dos (E2), está localizada frente al casco urbano de Ayapel (8o 18' 57,3' N y 75o 07' 34,5' O) en la zona más profunda de la ciénaga. La estación (E3), se encuentra ubicada en la zona cercana a Caño Pinto (8º 19' 21,9' N y 75º 06' 57,4' O), zona de alto depósito de materiales, altamente influenciada por los cambios de nivel debido a la influencia de los caños Barro y Muñoz. La estación cuatro (E4), se localizó en el sector La Miel (8o 20' 4' N y 75o 06' 38' O), presentando características e influencias similares a las de la estación E3. Por último la estación cinco (E5) (08º 23' 22,3' N y 75º 04' 56,1' O), se ubicó en la zona de la Ciénaga Los Toros, que representa un sector altamente influenciado por la entrada de los caños Grande y Barro.
Para la realización de las campañas de campo se tuvieron en consideración seis diferentes momentos del nivel hidrológico en la ciénaga: aguas altas en ascenso (15–30 de septiembre de 2004), aguas altas (12–27 de noviembre de 2004), aguas bajas en descenso (1–15 de febrero de 2005), aguas bajas (7–20 de marzo de 2005), aguas bajas en ascenso (25 de abril–6 de mayo de 2005) y aguas en ascenso (8–18 de junio de 2005) (figura 2).
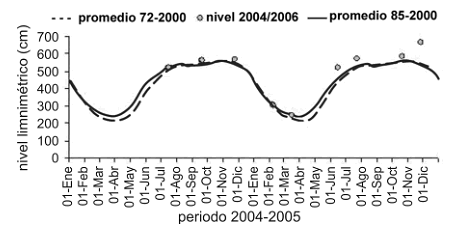
Figura 2. Variación de los niveles limnimétricos mensuales durante el año 2004/2005; series representativas de los promedios desde el año 1972–2000, 1985–2000 de la ciénaga de Ayapel (Córdoba), Colombia (fuente: Aguirre et al., 2005)
Muestreo de variables físicas y químicas. Por medio de una botella tipo Kemerer de volumen de 1,5 l se obtuvieron muestras integradas de la columna de agua, a las cuales les fueron medidas in situ la concentración de oxígeno disuelto por medio de un oxímetro modelo WTW, 300/Set; el pH y potencial redox mediante un peachímetro (medidor de pH) modelo WTW, 330/Set–1; la conductividad eléctrica y los sólidos disueltos con un conductímetro WTW, 300/set; y la transparencia se obtuvo utilizando un disco de Secchi. Las concentraciones de nitratos (NO–3), nitritos (NO–2), amonio (NH+4) y ortofosfatos (PO–34) fueron medidas en el laboratorio con un espectrofotómetro Spectroquant Nova 60 modelo KGaA 62293. La dureza total fue cuantificada por un kit Merck y los sólidos totales por medio del método gravimétrico.
Muestreo de la asociación fitoplanctónica. Las muestras de agua obtenidas de cinco secciones de la columna de agua (subsuperficie, 50% de la zona fótica, límite de la zona fótica, mitad de la profundidad de la columna de agua en la zona afótica y fondo (antes de tocar el sedimento), fueron mezcladas en un recipiente plástico del cual se retiró un volumen de un litro, el cual fue fijado con 5 ml de lugol al 10%. Para obtener las distancias de atenuación de la luz en las cuales se colectaron tres de las cinco submuestras en la zona fótica, se utilizó el coeficiente de atenuación de la luz descrito en los procedimientos de Ros (1979).
Análisis de laboratorio. La observación de las muestras se llevó a cabo con un microscopio invertido Leica DMIN, provisto de una reglilla ocular. La muestra de agua previamente agitada fue concentrada sedimentándose en una cámara de 50 ml. A continuación se retiró un mililitro del precipitado y se dispuso en una cámara de conteo de Sedgwick–Rafter en la cual se contaron 30 campos en un aumento de 400 X. Para la cuantificación de la densidad por mililitro se aplicó la expresión de Ros (1979).
La definición taxonómica de los taxones fitoplanctónicos se realizó bajo los criterios de Bourrelly (1966, 1968, 1985), Desikachary (1959), Huber–Pestalozzi (1938, 1976, 1955, 1961, 1983), Prescott et al. (1982), Ramírez (2000) y Strebel y Krauter (1988). Determinación del biovolumen o volumen celular. Para estimar el biovolumen se empleó el método del volumen celular medio a partir de las dimensiones de por lo menos 20 células seleccionadas aleatoriamente en el microscopio y la correspondencia de la forma celular a un sólido geométrico (Hillebrand et al., 1999). La densidad absoluta (en org/ml) de un taxón, así como el promedio del número de células de algas que forman colonias fue multiplicado por su volumen celular medio para obtener un estimativo de la biomasa de cada taxón en µm3/l. Para los filamentos verdaderos y falsos se consideró el volumen total de los mismos, para colonias y cenobios se determinó el número medio de células por individuo y con base en él se estimó el posterior volumen del organismo. Procesamiento y análisis de la información. Se presenta una tabla con los registros medios y de desviación estándar de cada una de las variables analizadas (tabla 1). Para cada estación y muestreo son presentados tablas y perfiles de abundancia con base en el biovolumen de los taxones, con el cual también fueron calculados los valores de los índices de diversidad (Shannon y Weaver, 1949), equidad (Pielou, 1969) y dominancia (Simpson, 1949), adicionalmente también se estimó la riqueza de numérica de especies. Con el fin de examinar la composición general descriptiva se determinó el índice de similaridad de Bray–Curtis con el cual se ejecutaron análisis de agrupamiento cuantitativo por medio del método de agrupamiento completo utilizando las abundancias de los taxones registrados en el estudio. Para determinar si los aportes de biovolumen y los índices de diversidad presentaron diferencias estadísticamente significativas entre momentos y sitios de muestreo se aplicó se aplicó la prueba no paramétrica Kruskall yWallis.
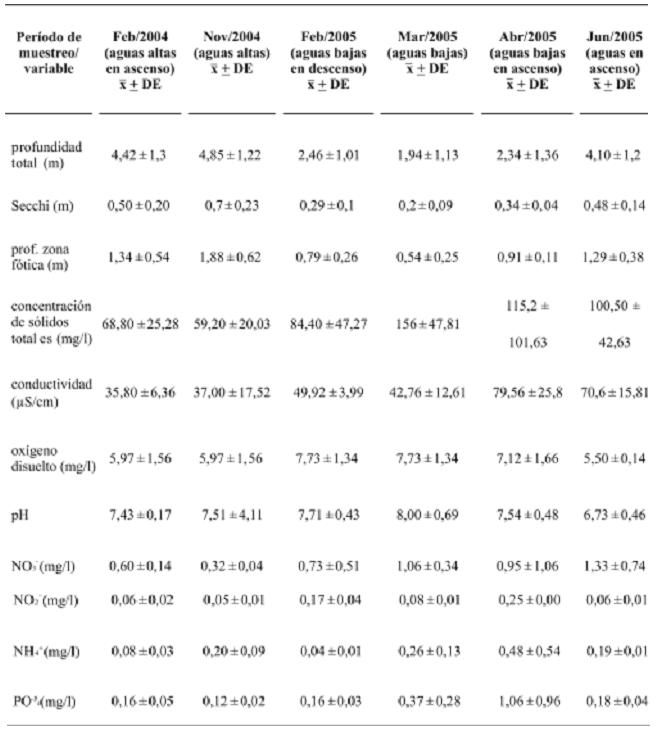
Tabla 1. Registro de los valores medios () y de desviación estándar (DE) para la ciénaga de Ayapel (Córdoba), Colombia, en las cinco estaciones durante los seis períodos de estudio [fuente de datos, matriz general del proyecto: Relación Río–Ciénaga, Ayapel (Aguirre et al., 2005)]
RESULTADOS
Variables físicas y químicas. Exceptuando la profundidad, las variables físicas y químicas presentaron un comportamiento espacial uniforme (tabla 1). Los efectos del pulso hidrológico de la ciénaga se reflejan claramente en la dinámica de la profundidad total, la transparencia Secchi y la profundidad de la zona fótica. Mientras las mayores profundidades y transparencias se presentaron en las épocas de lluvias (septiembre y noviembre), en los períodos secos (febrero y marzo) la columna de agua se redujo, especialmente en la estación cinco en la cual se registraron profundidades menores a los 0,5 m de profundidad. Los valores más altos de la concentración de sólidos totales fueron 168 mg/l en febrero y 220 mg/l en marzo, ambos muestreos correspondientes a época de estiaje. En los periodos de aguas bajas las transparencias registraron valores menores a 0,5 m de profundidad, especialmente en el mes de marzo.
En la ciénaga de Ayapel los mayores valores de conductividad tuvieron lugar en los muestreos realizados en abril y junio con valores entre 55,95 y 119,20 ìS/cm, mientras en los demás períodos de muestreo los valores fueron menores. El oxígeno disuelto presentó mayores concentraciones en los muestreos ejecutados en aguas bajas y en períodos de transición, la mayoría de estos valores fueron superiores a 6,0 mg/l. El incremento de esta variable fue paralelo a un aumento en el pH indicio de una mayor actividad fotosintética. Esta variable presentó un comportamiento homogéneo en el espacio, pero en el tiempo su variación está representada por el registro de valores más altos en los muestreos realizados en marzo y abril, en los cuales la mayoría de los valores son superiores a 7,3.
Las concentraciones de nitratos (NO3–) en el sistema cenagoso fluctuaron dentro de intervalos de sistemas libres de altas cargas de nutrientes (Bartram y Ballance, 1996; Chapman, 1996), ya que la mayoría de las concentraciones no superan 1 mg/l. Las formas de nitrito (NO2–) mostraron concentracio nes bajas, menores a 0,26 mg/l. El nitrógeno amoniacal (NH4+), no registró variaciones importantes y las concentraciones se mantuvieron en niveles menores a 0,48 mg/l. Sólo durante el mes de abril en la estación cinco fue registrado un valor alto (1,43 mg/l), este resultado podría estar reflejando la incidencia de Caño Grande en esta zona ya que durante esta época de muestreo las aguas comienzan su ascenso desde el río hacia la ciénaga. Con respecto a los ortofosfatos (PO–34) durante los periodos de lluvias (septiembre y noviembre 2004 y junio 2005) se presentaron las concentraciones mas bajas, mientras que en época seca y de transición (febrero, marzo y abril) se registraron las mayores concentraciones. La tendencia de los ortofosfatos fue similar a la de las diferentes formas de nitrógeno, valores relativamente mayores en los muestreos realizados en el mes de marzo y especialmente en el mes de abril. Las concentraciones registradas se pueden considerar en su mayoría bajas y algunas intermedias, en consecuencia no representan una limitación por fósforo para los productores primarios en la ciénaga de Ayapel.
Estructura de la asociación fitoplanctónica. La asociación fitoplanctónica en el sistema de Ayapel estuvo dominada por la división Chlorophyta en términos del número o riqueza de taxones, pero en términos del biovolumen el grupo más importante fueron las algas verde–azules. En total se registraron 37 taxones fitoplanctónicos en las cinco estaciones y los seis muestreos. Entre todos estos taxones, doce pertenecen a la división Chlorophyta, seis a la división Cyanoprocariota, cinco a Euglenophyta, once a Bacillariophyta, uno a la división Xantophyta, al igual que uno para la división Dynophyta. A pesar de que las algas verdes y las diatomeas agrupan el mayor número de taxones, las cianoprocariotas Cylindrospermopsis raciborskii y Lyngbya cf. limnetica mostraron los registros de biovolumen más importantes durante el período de estudio (figura 3), por lo cual las cianoprocariotas en general registran un porcentaje importante del biovolumen.
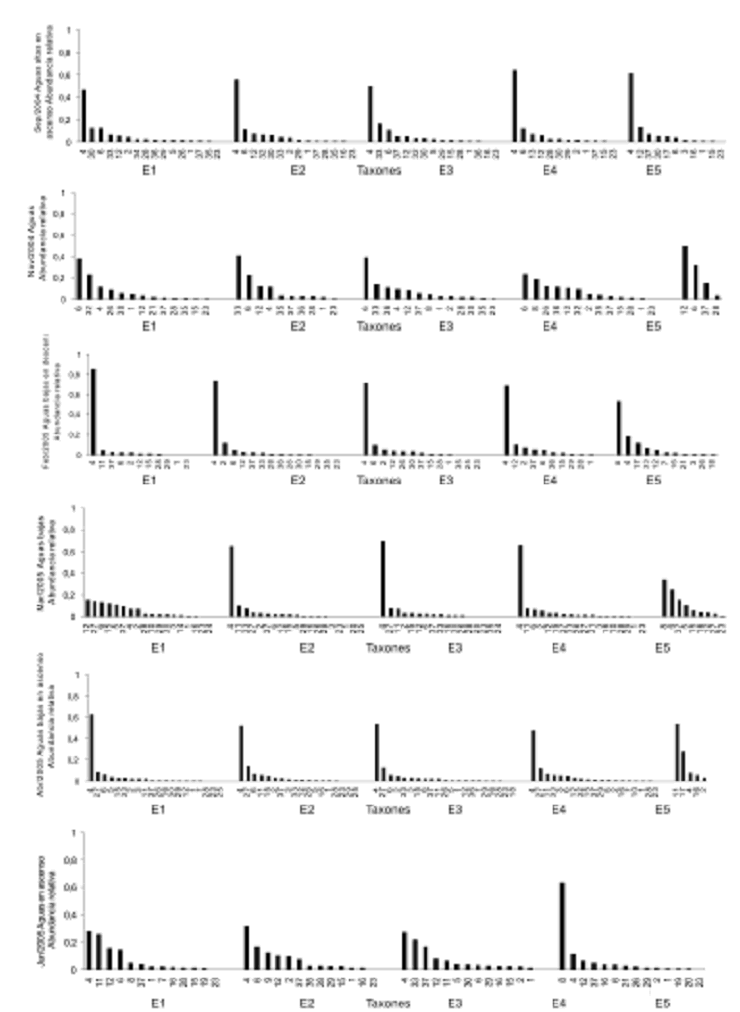
Figura 3. Variación espacio–temporal del biovolumen relativo de la asociación fitoplanctónica en la ciénaga de ciénaga de Ayapel (Córdoba), Colombia, en el período de estudio (E = estación). La numeración del eje x corresponde a los taxones fitoplanctónicos 1. Merismopedia minina, 2. Coelosphaerium naegelianium, 3. Anabaena cf. fertilíssima, 4. Cylindrospermopsis raciborskii, 5. Oscillatoria sp., 6. Lyngbya cf. limnetica, 7. Euglena oxyuris, 8. Phacus cf. longicauda, 9. Phacus pleuronectes, 10. Strombomonas cf. gibberosa, 11. Trachelomonas cf. ecrea, 12. Aulacoseira granulata, 13. Fragilaria sp. 2, 14. Fragilaria sp. 1, 15. Synedra cf. ulna, 16. Navicula sp., 17. Pinnularia cf. episcopalis, 18. Cymbella sp., 19. Gomphonema sp., 20. Eunotia sp., 21. Nitzschia cf. acicularis, 22. Cyclotella sp., 23. Scenedesmus brasiliensis, 24. Coelastrum cf. microporum, 25. Crucigenia sp., 26. Ankistrodesmus cf. bernardii, 27. Monoraphidium contortum, 28. Hyaloraphidium contortum, 29. Cosmarium sp., 30. Closterium setaceum, 31. Closterium sp., 32. Tetraedrom gracile, 33. Staurastrum leptocladum, 34. Xanthidium sp., 35. Centritractus belanophorus, 36. Peridinium cf. bipes, 37. Cryptomonas sp.
Los estimadores de riqueza Chao y Jack–knife, sugirieron que el número de especies esperado para la asociación es de 38 y 39, respectivamente, cifra cercana a la riqueza observada, la cual fue de 37 taxones.
Distribución espacio–temporal. En la figura 3 se observan las curvas descendentes de las abundancias relativas de los gremios fitoplanctónicos hallados en cada estación durante cada muestreo. Se puede observar en general que siempre se presentó un taxón con biovolumen importante acompañado en promedio por once taxones de menor importancia. En la mayoría de los casos la abundancia relativa del taxón predominante fue dos veces la abundancia de los demás taxones presentes. Se obtuvo además que los taxones con mayor abundancia se hallaron entre en las épocas secas y de transición a lluvias (muestreos realizados en febrero, marzo y abril).
Estas especies como se mencionó anteriormente fueron C. raciborskii y L. cf. limnetica, siendo C. raciborskii el más importante;sin embargo, otros taxones de menor abundancia fueron altamente frecuentes. Entre éstos se destacaron Aulacoseira granulata, Hyaloraphidium contortum y Cryptomonas sp.
Tendencias de distribución similar en la composición de taxones fueron también reflejadas por el análisis de agrupamiento (figura 4). En este gráfico, las estaciones uno a cuatro tienden a presentar un mayor porcentaje de similaridad con respecto a la estación cinco, las cual se subagrupa en el extremo superior del esquema. La composición espacio–temporal general de los taxones durante el período de estudio presentó un alto porcentaje de similaridad (44,5%), lo cual sugiere que existe un importante número de taxones de presencia común entre estaciones y muestreos (figura 4).
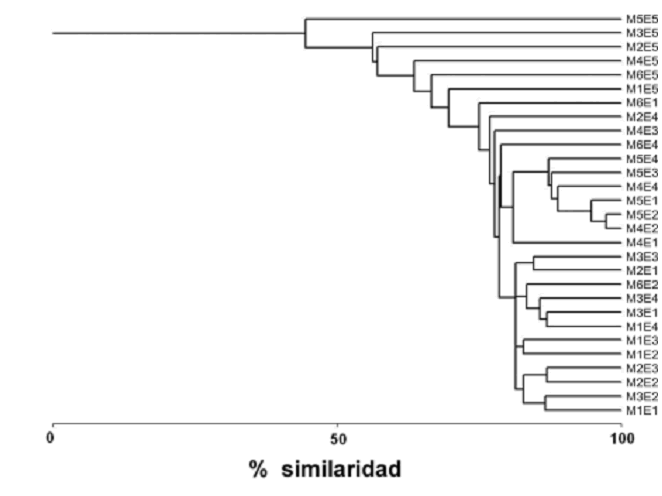
Figura 4. Análisis de agrupamiento entre las especies registradas durante el estudio empleando el índice de similaridad de Bray–Curtis y ligamiento completo con base en la presencia–ausencia de los taxones fitoplanctónicos en la ciénaga de Ayapel (Córdoba), Colombia (M = muestreo; E = estación)
Biovolumen. En la figura 5 se ilustra el perfil del biovolumen celular total para cada estación por momento de muestreo. Para los muestreos de las épocas secas y de transición a lluvias (febrero, marzo y abril, respectivamente) se presentó un incremento considerable del biovolumen, en especial en las estaciones uno, dos, tres y cuatro, las cuales se encuentran ubicadas en el eje central del espejo principal de la ciénaga; las diferencias entre muestreos fueron estadísticamente significativas (p 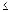 0,05). Con respecto a los demás muestreos el bajo biovolumen presentó poca variación entre estaciones.
0,05). Con respecto a los demás muestreos el bajo biovolumen presentó poca variación entre estaciones.
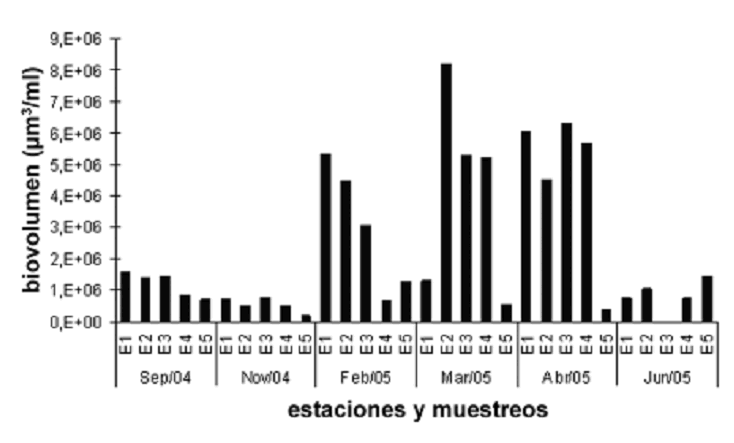
Figura 5. Variación espacio–temporal del biovolumen (µm3/ml) durante el período de estudio en la ciénaga de Ayapel (Córdoba), Colombia. Valores derivados de la prueba Kruskal–Wallis: (estaciones: 7,45, p = 0,11; muestreos: 10,83, p = 0,05)
Índices de diversidad. Los índices de diversidad y equidad no variaron significativamente y sus coeficientes de variación en el período de estudio no presentan un valor superior al 24%. Por su parte, el índice de dominancia registró una variación alta (42%), ya que durante los muestreos realizados en época seca y de transición a lluvias las abundancias estuvieron concentradas en C. raciborskii. La tendencia temporal muestra que mientras en febrero el índice de dominancia es mayor, la distribución en noviembre fue más uniforme, es así como los perfiles de distribución expresados por medio de los índices reflejaron contrastes entre los dos períodos hidrológicos (figura 6). En cuanto a los valores medios registrados, Shannon podría ser considerado intermedio (1,59 nats/ind), la equidad de Pielou tiende a ser alta (0,63) y la dominancia de Simpson es baja (0,34).
Figura 6. Perfil espacio–temporal de los índices de diversidad (H`), equidad (J) y dominancia (D). Prueba Kruskal–Wallis Diversidad Shannon y Weaver (estaciones: 3,67, p = 0,45; muestreos: 10,21, p = 0,07); equidad de Pielou (estaciones: 1,88, p = 0,76; muestreos: 13,32, p = 0,02); dominancia de Simpson (estaciones: 1,21, p = 0,88; muestreos: 12,31, p = 0,03), cálculos basados en el biovolumen de la ciénaga de Ayapel (Córdoba), Colombia
DISCUSIÓN
Las planicies de inundación son reconocibles como mosaicos de ecosistemas altamente dinámicos, donde la estabilidad y la diversidad están condicionadas primariamente por la hidrología y los flujos de materiales (Neiff, 1999). Varios autores coinciden en la identidad propia de estos sistemas (Gopal, 1994; Mitsch y Gosselink, 1993; Neiff, 1999) y particularmente en Colombia, Ramírez y Viña (1998) han mencionado que las condiciones bióticas y abióticas de los sistemas cenagosos están estrechamente ligadas a los períodos climáticos.
En términos generales y sólo con algunas excepciones, tanto las características físicas, químicas y bióticas del sistema cenagoso de Ayapel mostraron una variación temporal importante asociada al pulso hidrológico. A pesar de que la dinámica fisicoquímica del sistema cenagoso de Ayapel es relativamente homogénea en el espacio, la asociación fitoplanctónica respondió con un incremento en el biovolumen registrado en la época seca y de transición a lluvia en contraste con la época de aguas altas en el cual fue menor.
Durante el período de estudio la ciénaga de Ayapel podría ser caracterizada como un sistema de pH moderado, altamente turbio, sobretodo durante las épocas secas, con reducidas transparencias en sitios cuya profundidad es menor a 0,50 m en febrero y marzo respectivamente, épocas en las cuales se registró una mayor concentración de sólidos totales y disueltos; estas respuestas manifiestan los efectos del pulso en el sistema, dilución en época lluviosa (aumento de nivel) y concentración de partículas en la época seca (bajo nivel). La dinámica de los nutrientes también reflejó el contraste del pulso hidrológico entre los diferentes períodos de muestreo, evidenciando tendencias a una concentración mayor durante el estiaje con respecto a la temporada de lluvias altas. De esta manera, las formas de nitrógeno (amonio y nitrato), así como las concentraciones relativamente intermedias de ortofosfatos revelaron el mismo patrón temporal: mayores concentraciones durante la época seca y de transición a lluvia.
La variación de las agremiaciones fitoplanctónicas en climas tropicales es controlada por la disponibilidad de nutrientes y la radiación subacuática (Esteves, 1998). En el sistema de Ayapel se podría decir que la disponibilidad de luz podría ser un factor de peso en la baja densidad o biovolumen del fitoplancton ya que las transparencias son reducidas y son aún menores cuando el nivel de la ciénaga desciende en respuesta a la reducción en la columna de agua y la resuspensión de partículas aumentada por la acción del viento en una columna poco profunda.
Como se señaló para las variables ambientales, la asociación fitoplanctónica también varió en respuesta al pulso hidrológico; así lo reflejaron los principales descriptores estructurales de la asociación como la distribución de abundancias, el biovolumen y los índices de diversidad. Sin embargo, algunos atributos generales como el número y la composición de especies no mostraron variaciones importantes.
La composición de la agremiación fitoplanctónica presentó una importante similaridad espacio–temporal, lo que demuestra la adaptabilidad de los taxones más importantes ante la variación del pulso.
El fitoplancton estuvo dominado por taxones que presentan adaptaciones a bajas concentraciones de nutrientes en forma inorgánica. Así, Cylindrospermopsis y Lyngbya poseen una alta afinidad por el fósforo; además, Cylindrospermopsis posee la capacidad de fijar nitrógeno atmosférico, gracias a lo cual puede hacerse dominante en condiciones de bajas concentraciones de nutrientes.
Adicionalmente las especies de estos dos géneros poseen una alta capacidad de suspensión, gracias a lo cual, las pérdidas por sedimentación son bajas. Todo lo anterior hace que las poblaciones de Lyngbya y Cilindrospermopsis sean muy estables una vez que éstas se establecen (Mur et al., 1999). Ambas especies son pequeñas formas filamentosas pertenecientes a la división Cyanoprocariota las cuales responden rápidamente a los cambios ambientales, estas algas pueden multiplicarse especialmente en situaciones de estrés ambiental que en el caso del complejo de Ayapel son promovidas por las perturbaciones naturales de estiaje y llenado del sistema y esta facultad o estrategia ecológica les confiere cierta ventaja en un sistema de pulso de nivel como el de Ayapel.
Otros taxones de baja abundancia pero muy frecuentes en el período de estudio fueron Cryptomonas y Aulacoseira. Las especies del género Cryptomonas son consideradas como algas oportunistas de rápido crecimiento, gracias a su capacidad heterotrófica facultativa; se ven particularmente favorecidas en ambientes con bajas concentraciones de nutrientes mineralizados, en donde al tiempo que se dan altas turbiedades y concentraciones moderadas de nutrientes en forma orgánica, (Ortega y Rojo, 2000; Tell et al., 2005). Por su parte las especies del género Aulacoseira presentan un alto grado de variabilidad morfológica intraespecífica que está asociado con su capacidad de adaptación a muy diversas condiciones hidráulicas, gracias a lo cual pueden encontrarse tanto en ambientes lóticos como lénticos, pero preferiblemente someros. Pueden ser abundantes en lagunas de inundación (ciénagas) particularmente durante la época de llenado, cuando se resuspenden desde el sedimento (Farrell et al., 2001).
En términos generales las concentraciones de nitratos y ortofosfatos registradas independientemente de la variación espacio–temporal no representan una limitación por fósforo para los productores primarios en la ciénaga de Ayapel.
Factores físicos y climatológicos son responsables de los determinantes ecológicos de la ocurrencia y dominancia de C. raciborskii (Bouvy et al., 2000; Nascimento et al., 2000) y con respecto a esto en la ciénaga de Ayapel, un área expuesta a cambios abióticos fuertes influenciados por el pulso hidrológico, los filamentos de este organismo seleccionan como estrategia ecológica producir un número importante de descendientes de pequeño tamaño, característica de los estrategas C de selección r. Según Margalef (1983), los organismos fitoplanctónicos en los sistemas tropicales son más estrategas r que k; lo cual corresponde a una estrategia de adaptación a la vida planctónica estrechamente relacionada con la turbiedad y la concentración de nutrientes, como en el caso de Ayapel.
La especie C. raciborskii, puede a menudo dominar el fitoplancton de algunas asociaciones (Padisák, 1997). Dentro de los grupos funcionales de fitoplancton propuestos por Reynolds (1998), Cylindrospermopsis es un típico representante de aguas cálidas de capas mezcladas y supremamente tolerante a deficiencias de nitrógeno y luz (Reynolds et al., 2002), como en el caso de Ayapel sobretodo en la época seca, cuando la zona fótica es mayormente reducida. Para C. raciborskii, se reporta baja palatabilidad (Bouvy et al., 2001) lo que sugiere que además de su adaptabilidad ecológica su población se favorece por la baja herviboría. En estos términos y en el marco de bajas densidades fitoplanctónicas en el sistema, la mayor disponibilidad de C. raciborskii no constituye una importante oferta ambiental trófica en el sistema.
Cylindrospermopsis raciborskii es potencialmente tóxica por la producción de hepatotoxinas (Ohtani et al., 1992) y neurotoxinas (Lagos et al., 1999), por lo cual su monitoreo es importante para la salud humana y los aspectos ambientales de un sistema acuático, adicionalmente, posee la capacidad de formar florecimientos y de disminuir la biodiversidad por medio de la eliminación de otras especies fitoplanctónicas (Bouvy et al., 2000).
Adicionalmente, se debe tener en cuenta la incidencia de los vientos en conjugación con la profundidad de la columna muestreada, la columna de agua de la estación cinco fue menos profunda con respecto a las demás estaciones, hecho que se acentuaba durante las épocas secas, por lo cual la influencia del viento en una somera columna de agua provocó mayor resuspensión de sedimentos y en consecuencia, una importante limitación de luz en la zona fótica afectando el desarrollo fitoplanctónico, evidencia de ello fue que en esta estación no se registró un incremento del biovolumen en época seca y de transición a lluvias como sucedió en las demás estaciones.
Ramírez y Viña (1998), basándose en el estudio de diferentes ciénagas en el país encontraron que la turbidez con respecto a la disponibilidad de nutrientes juega un papel mucho más importante en el desarrollo de altas densidades fitoplanctónicas, ya que la reducida transparencia limita la disponibilidad de luz, caso del sistema cenagoso de Ayapel. En contraste con otros sistemas cenagosos, Ayapel experimenta una importante exportación de materiales y un bajo tiempo de retención hidráulica (Aguirre et al., 2005). Zapata (2005) encontró que el tiempo medio de residencia anual en la ciénaga es de 37 días, se podría considerar que este tiempo de residencia es bajo. Históricamente en Ayapel los mayores tiempos de retención hidráulica se alcanzan en los meses de mayo (52,5 días) y septiembre (41,1 días) y a medida que el volumen disminuye se alcanzan los menores valores en el mes de marzo (22.1 días) (Aguirre et al., 2005). Estos hallazgos conjuntamente con los reducidos valores de la transparencia permiten suponer que los gradientes temporales son los que gobiernan las variaciones ambientales y ecológicas del fitoplancton en la ciénaga. En varias ciénagas del país se ha encontrado que especies pertenecientes al grupo de las cianoprocariotas tienen una gran importancia en la dinámica del sistema y se ha mencionado que sus abundancias no pueden ser atribuidas a bajas relaciones nitrógeno–fósforo, sino a una condición oportunista ante fluctuaciones ambientales.
CONCLUSIONES
La estructura de la asociación fitoplanctónica en la ciénaga de Ayapel registró cambios en el biovolumen, el perfil de abundancias y los índices de diversidad en los diferentes momentos del pulso hidrológico; así mismo, las variables ambientales influenciadas por el pulso, principalmente la transparencia, la concentración de nutrientes y los sólidos totales registraron variaciones en los valores según el momento de muestreo. Con respecto a la composición de la asociación fitoplanctónica fue similar espacio–temporalmente mientras que el biovolumen aumentó durante el estiaje cuando el nivel del agua disminuyó y la concentración de nutrientes es levemente mayor. El reducido número de taxones encontrados podría estar relacionado con el corto tiempo de residencia de la ciénaga y la baja transparencia del agua, ya que implican menor permanencia de los hábitats para la suspensión del fitoplancton en una zona fótica reducida. Considerando que el pulso hidrológico es el factor determinante de la dinámica de ecosistemas de planicie de inundación (Huszar, 2003; Huszar et al., 1999; Junk et al., 1989; Neiff, 1999; Sioli, 1984) se espera que las agremiaciones fitoplanctónicas sean profundamente influenciadas por los pulsos hidrológicos del macrosistema, éste fue el caso de la ciénaga de Ayapel, ya que atributos estructurales de la asociación fitoplanctónica cambiaron en función del pulso hidrológico muestreado.
AGRADECIMIENTOS
Los autores manifiestan sus más sinceros agradecimientos a COLCIENCIAS, al IDEAM por la información sobre los registros históricos de los niveles limnimétricos mensuales en la ciénaga, a CORPOAYAPEL y a los estudiantes y profesores de las universidades de Antioquia y Nacional sede Medellín que participaron en el proyecto.
REFERENCIAS
1. Aguirre NJ, Palacio JA, Flórez MT, Wills A, Caicedo O, Jiménez LF, Villegas NE, Palacio CA, Toro M, Vélez JI. 2005. Informe final del proyecto COLCIENCIAS: Análisis de la relación río–ciénaga y su efecto sobre la producción pesquera en el sistema cenagoso de Ayapel, Colombia. Grupo de Investigación en Gestión y Modelación Ambiental GAIA, Facultad de Ingeniería, Universidad de Antioquia. Medellín, Colombia. [ Links ]
2. Bartram J, Ballance R. 1996. Water quality monitoring: a practical guide to the design and implementation of freshwater quality studies and monitoring programmes. UNEP, WHO. Londres, Inglaterra. [ Links ]
3. Bourrelly P. 1966. Les algues d'eau douce. Initiation à la systématique. Tome I: Les Algues Vertes. N. Boubée y Cie. París, Francia. [ Links ]
4. Bourrelly P. 1968. Les algues d'eau douce. Initiation à la systématique. Tome II: Les Algues jaunes et brunes. Chrysophycées, Phéophycées, Xanthophycées et Diatomées. N. Boubée y Cie. París, Francia. [ Links ]
5. Bourrelly P. 1985. Les algues d'eau douce. Initiation à la systématique. Tome III: Les Algues bleues et rouges. Les Euglénoides, Peridinies et Cryptomonadines. N. Boubée y Cie. París, Francia. [ Links ]
6. Bouvy M, Pagano M, Troussellier M. 2001. Effects of a Cyanobacterial bloom (Cylindrospermopsis raciborskii) on bacteria and zooplankton communities in Ingazeira reservoir (northeast Brazil). Aquatic Microbial Ecology, 25:215–227. [ Links ]
7. Bouvy M, Falcão D, Marinho M, Pagano M, Moura A. 2000. Occurrence of Cylindrospermopsis (Cyanobacteria) in 39 Brazilian tropical reservoirs during the 1998 drought. Aquatic Microbial Ecology, 23:13–27. [ Links ] [ Links ]
9. Desikachary TV. 1959. Cyanophyta. Indian Council of Agricultural Research New Delhi, Department of Botany, University of Madras. India. [ Links ]
10. Ecoestudios y CVS. 1989. Estudio del manejo integral de la Ciénaga Ayapel. Montería (Córdoba), Colombia. [ Links ]
11. Esteves F. 1998. Fundamentos de limnología. Segunda edición. Interciencia. Río de Janeiro, Brasil. [ Links ]
12. O'Farrell I, Tell G, Podlejski A. 2001. Morphological variability of Aulacoseira granulata (Ehr.) Simonsen (Bacillariophyceae) in the Lower Paraná River (Argentina). Limnology, 2(2):65–71. [ Links ]
13. Gopal B. 1994. The role of ecotones (transiton zones) in the conservation and management of tropical inland waters. Mitteilungen Internationale Vereinigung für Limnologie, 24:17–25. [ Links ]
14. Hillebrand H, Dürselen CD, Kirschtel D, Tamar UP. 1999. Biovolume calculation for pelagic and benthic microalgae. Journal of Phycology, 35:403–424. [ Links ]
15. Huber–Pestalozzi G. 1938. Das Phytoplankton des Süßwassers. Sistematik und Biologie. Allgemeiner Teil: Blaualgen, Bakterien, Pilze. Band XVI, Teil 1, Hälfte 1. En: Thienemann A, Elster HJ, Ohle W (eds.). Die Binnengewässer. Schweizerbart. Stuttgart, Alemania. [ Links ] [ Links ]
17. Huber–Pestalozzi G. 1961. Das Phytoplankton des Süßwassers. Sistematik und Biologie. Chlorophyceae (Grünalgen). Ordnung: Volvocales. Band XVI, Teil 5. En: Thienemann A, Elster HJ, Ohle W (eds.). Die Binnengewässer. Schweizerbart. Stuttgart, Alemania. [ Links ]
18. Huber–Pestalozzi G. 1976. Das Phytoplankton des Süßwassers. Sistematik und Biologie. Chrysophyceen. Farblose Flagellaten. Heterokonten. Band XVI, Teil 2, Hälfte 1. En: Thienemann A, Elster HJ, Ohle W (eds.). Die Binnengewässer. Schweizerbart. Stuttgart, Alemania. [ Links ]
19. Huber–Pestalozzi G. 1983. Das Phytoplankton des Süßwassers. Sistematik und Biologie. Chlorophyceae (Grünalgen) Ordnung: Chlorococcales. Band XVI, Teil 7, Hälfte 1. En: Thienemann A, Elster HJ, Ohle W (eds.). Die Binnengewässer. Schweizerbart. Stuttgart, Alemania. [ Links ]
20. Huszar VLM, Da Silva LH. 1999. A estructura da comunidade fitoplanctônica no Brasil: cinco décadas de estudos. En: Sociedade Brasileira de Limnologia (ed.). LIMNOtemas. Río de Janeiro, Brasil. [ Links ]
21. Huszar VLM. 2003. Memorias del curso de ecología del fitoplancton continental. Universidad Nacional de Colombia. Bogotá, Colombia. [ Links ]
22. IGAC. 1986. Estudio general de suelos de los municipios de Ayapel, Buenavista, Planeta Rica y Pueblo Nuevo (Departamento de Córdoba). Instituto Geográfico Agustín Codazzi, Subdirección Agrológica. Bogotá, Colombia. [ Links ]
23. Junk WJ, Bayley PB, Sparks RE. 1989. The 'flood pulse' concept in river floodplain systems. Canadian Special Publication of Fisheries and Aquatic Sciences, 106:110–127. [ Links ]
24. Lagos N, Onodera H, Zagatto PA, Andrinolo D, Azevedo SM, Oshima Y. 1999. The first evidence of paralytic shellfish toxins in the freshwater cyanobacterium Cylindrospermopsis raciborskii, isolated from Brazil. Toxicon, 37:1359–1373. [ Links ]
25. Margalef, R. 1983. Limnología. Omega S. A. Barcelona, España. [ Links ]
26. Mitsch W, Gosselink JG. 1993. Wetlands. Van Nostrand Reinhold. Nueva York, Estados Unidos. [ Links ]
27. Mur RL, Skulberg OM, Utkilen H. 1999. Cyanobacteria in the Environment. En: Chorus I and Bartram J (eds.). Toxic Cyanobacteria in water: A guide to their public health consequences. St. Edmundsbury Press, Bury St Edmunds, Suffolk. Londres, Inglaterra. [ Links ] [ Links ]
29. Neiff JJ. 1999. El régimen de pulsos en ríos y grandes humedales de Sudamérica. Pág. 99 En: Málvarez AI (ed.). Tópicos sobre humedales subtropicales y templados de Sudamérica. Impreso por la Oficina Regional de Ciencia y Tecnología de la UNESCO para América Latina y el Caribe _ORCYT. Montevideo, Uruguay. [ Links ]
30. Ohtani I, Moore R, Runnegar MTC. 1992. Cylindrospermopsin: a potent hepatotoxin from the blue–green alga Cylindrospermopsis raciborskii. Journal of the American Chemical Society, 114:7941–7942. [ Links ]
31. Ortega E, Rojo C. 2000. Fitoplancton del Parque Nacional Las Tablas de Daimiel. II. Diatomeas y clorofitas. Anales del Jardín Botánico de Madrid, 58(1):17–37. [ Links ]
32. Padisák J. 1997. Cylindrospermopsis raciborskii (Woloszynska) Seenayya et Subba Raju, an expanding, hightly adaptive cyanobacterium: worldwide distribution and review of its ecology. Archiv für Hydrobiologie, Supplement, 4:563–593. [ Links ]
33. Pielou EC. 1969. Shannons formula as a measure of specific diversity–its use and disuse. American Naturalist, 100:463–465. [ Links ]
34. Prescott GW, Bicudo CE, Vinyard WC. 1982. Desmidiaceae: Placodermae. Part II. Section 4. En: Prescott GW, Bicudo CE, Vinyard WC. A Synopsis of North American Desmids. University of Nebraska Press, Lincoln and London. [ Links ]
35. Ramírez A, Viña G. 1998. Limnología colombiana: aportes a su conocimiento y estadísticas de análisis. Primera edición. Editorial Panamericana. Bogotá, Colombia. [ Links ]
36. Ramírez JJ. 2000. Fitoplancton de agua dulce: aspectos ecológicos, taxonómicos y sanitarios. Editorial Universidad de Antioquia. Medellín, Colombia. [ Links ]
37. Reynolds CS. 1998. What factors influence the species composition of phytoplankton in lakes of different trophic status? Hydrobiologia, 369:11–26. [ Links ]
38. Reynolds CS, Huszar V, Kruk C, Flores LN, Melo S. 2002. Towards a functional classification of the freshwater phytoplankton. Journal of Plankton Research, 24(5):417–428. [ Links ]
39. Ros J. 1979. Prácticas de ecología. Ediciones Omega S. A. Barcelona, España. [ Links ]
40. Shannon CE, Weaver W. 1949.The Mathematical Theory of Communication. University of Illinois Press. Urbana, Estados Unidos. [ Links ]
41. Simpson EH. 1949. Measurement of diversity. Nature, 163:688. [ Links ] [ Links ]
43. Strebel H, Krauter D. 1988. Das Leben im Wassertropfen. Mikroflora und Mikrofauna des Süßwassers. Franckh–Kosmos Naturführer. Stuttgart, Alemania [ Links ]
44. Tell G, O'Farrell I , Lombardo G. 2005. Euglenoid morphospecies replacement along a hydraulic gradient of the Lower Paraná Basin (Argentina). Freshwater Biology, 50:616. [ Links ]
45. Zapata JF. 2005. Dinámica hidrológica en la ciénaga de Ayapel, modelación del balance hídrico y simulación hidrodinámica. Tesis de maestría. Maestría en Recursos Hidráulicos. Universidad Nacional de Colombia, Medellín, Colombia. [ Links ]

