










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActualidades Biológicas
Print version ISSN 0304-3584
Actu Biol vol.30 no.89 Medellín July/Dec. 2008
ICTIOLOGÍA
VARIACIÓN ESPACIO–TEMPORAL DE LA ASOCIACIÓN DE ESPECIES DE PECES EN LA LAGUNA DE CACHIMBERO, UN HUMEDAL EN LA CUENCA MEDIA DEL RÍO MAGDALENA, COLOMBIA
SPATIO–TEMPORAL CHANGES OF FISH ASSEMBLAGES IN LAGUNA CACHIMBERO, A WETLAND IN THE MIDDLE MAGDALENA RIVER BASIN, COLOMBIA
Alexandra Arango–Rojas1, 2; Luz F. Jiménez–Segura2, 3; Jaime A. Palacio–Baena4
1Grupo de Gestión y Modelación Ambiental. Sede de Investigaciones Universitarias (SIU), Universidad de Antioquia. A. A 1226. Medellín, Colombia. Dirección Electrónica: <aarango@uco.edu.co>
2Instituto de Biología, Bloque 7, Universidad de Antioquia, A. A. 1226. Medellín, Colombia.
3Grupo de Gestión y Modelación Ambiental. Sede de Investigaciones Universitarias (SIU), Universidad de Antioquia. A. A 1226. Medellín, Colombia. Dirección Electrónica: <ljimenez@matematicas.udea.edu.co>
4Grupo de Gestión y Modelación Ambiental. Sede de Investigaciones Universitarias (SIU), Universidad de Antioquia. A. A 1226. Medellín, Colombia. Dirección Electrónica: <japalaci@jaibana.udea.edu.co>
Resumen
La asociación de peces fue estudiada en la Laguna de Cachimbero en diferentes momentos pluviométricos. Las capturas fueron realizadas en cuatro sitios; en cada uno de ellos se muestreo las zonas litoral y pelágica. Fueron capturadas con redes de espera: 23 especies de peces, 1.372 individuos y un peso total de 96.160,28 g. Las especies Caquetaia kraussii, Cyphocharax magdalenae, Hoplias malabaricus, Pimelodus blochii, Prochilodus magdalenae y Trachelyopterus insignis fueron las más importantes en abundancia y biomasa. La mayoría de las especies fueron capturadas en zonas litorales y en sitios de muestreo más próximos a los tributarios del humedal y del canal de salida al Río Magdalena y el Caño Cachimbero. La Captura por Unidad de Esfuerzo total (cpue, g/m2) fue diferente entre las zonas litoral y pelágica y entre sitios de muestreo. Los valores de diversidad no superaron en ningún caso a 0,8 bits. La equidad alcanzó valores significativamente altos en algunos muestreos. Cyphocharax magdalenae, Pimelodus blochii y Caquetaia kraussii fueron las especies más importantes en la asociación espacio–temporal. No se encontró una relación significativa entre las características del medio y la asociación de especies.
Palabras claves: asociación de peces, humedal, Río Magdalena, variación espacial y temporal
Abstract
Fish assemblage was studied in the Cachimbero lagoon in different raining periods. Catches were made in four sampling sites; each one was sampled its litoral and pelagic zones. 23 species of fish, 1,372 specimens (96,160.28 g) were caught with the gillnets. Caquetaia kraussii, Cyphocharax magdalenae, Hoplias malabaricus, Pimelodus blochii, Prochilodus magdalenae, and Trachelyopterus insignis had the highest number of individuals and mass. Most of species were founded in litoral zones and in the sampling sites nearest to tributaries and to the outflow channel to the Magdalena River, the Cachimbero Channel. Total CPUE were different between litoral and pelagic zones and among sampling sites. Diversity values in each sampling never ever exceed 0.8 bits. Evenness was high in some climatic moments. Cyphocharax magdalenae, Pimelodus blochii, and Caquetaia kraussii were the most important fish species in the fish assemblage. We did not found any relationship between environment characteristics and the species assemblage.
Key words: fish assemblage, Magdalena River, temporal and spatial variation, wetland
INTRODUCCIÓN
Las asociaciones de especies de peces en los ríos tropicales se ven sometidas a cambios en su estructura, debido a que en dichos sistemas se presentan fuertes variaciones en el nivel del agua, asociadas con el régimen pluvial (Bayley, 1996; Junk et al., 1989; Welcomme, 1979). El incremento en el nivel de los ríos provoca su desborde hacía las áreas laterales, restableciendo la conexión entre los humedales presentes en su plano lateral y el cauce principal del río. Esta condición, que se sucede de manera periódica, activa el intercambio de organismos entre estos ambientes, potenciando cambios evidentes en la conformación de sus asociaciones de especies y en su estructura trófica (Barthem y Goulding, 1997). La reducción en la conexión entre estos sistemas es una de las causas del cambio de las comunidades de peces presentes en ellos y, en consecuencia, de su productividad (Welcomme, 1979).
La cuenca del río Magdalena es la principal vía fluvial de Colombia y tal vez la cuenca fluvial con mayor influencia humana (Galvis y Mojica, 2007). La perdidas de vegetación en sus áreas de drenaje, la construcción de presas para generación de energía eléctrica, el vertimiento de residuos sin tratamiento previo y la reducción del área de sus humedales (localmente denominadas como ''ciénagas'') debido al avance de su frontera agropecuaria son una amenaza real para la ictiofauna y la sostenibilidad del recurso pesquero (Jiménez–Segura, 2007). La construcción de diques en los canales de conexión con el canal principal del río y la posterior desecación de estos cuerpos de agua, vitales para los peces, son prácticas comunes en la cuenca media y baja de esta cuenca. A continuación se presenta una evidencia importante sobre cómo el aislamiento de estos sistemas del río principal influye de manera directa sobre la asociación de especies de peces en estos humedales del río Magdalena.
MATERIALES Y MÉTODOS
Área de estudio. La laguna de Cachimbero es una ciénaga que hace parte de un complejo cenagoso localizado en el corregimiento de Puerto Olaya, municipio de Cimitarra (Santander), y dentro de la región geográfica del Magdalena medio colombiano. Se encuentra a una altitud promedio de 200 m, es alimentada por una microcuenca con una área aproximada de 17.000 ha y sus aguas se vierten al río Magdalena por los caños Cachimbero y Negro (Restrepo y Toro, 1997). Tiene un área de espejo libre de aproximadamente 3,7 km2 y una faja perimetral de humedales con vegetación hidrófila de casi 11 km2 (figura 1). La influencia del río Magdalena en la laguna de Cachimbero es débil puesto que se sus canales de conexión han sido afectados: hay un dique en caño Negro y, la boca del caño Cachimbero fue cerrada debido al avance de la vegetación acuática.
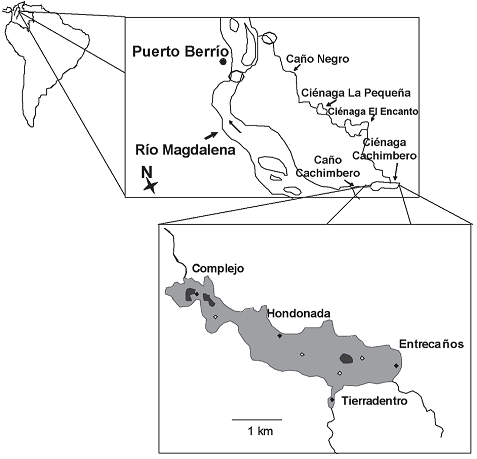
Figura 1. Ubicación de la laguna de Cachimbero, Cimitarra, (Santander) Colombia
Métodología. El trabajo de campo se realizó en cinco momentos pluviométricos contrastantes; durante el año 2003 en los meses de marzo, mayo, octubre y diciembre y en 2004 durante abril, en cada uno de ellos se muestreo durante cinco días consecutivos. La pesca se realizó en cuatro sitios: Entrecaños, Tierradentro, Hondonada y Complejo, todos ubicados dentro del espejo de agua libre y de acuerdo con la infuencia de pequeños tributarios (incluyendo caño Negro) y la boca del caño Cachimbero. En cada sitio, se ubicaron las redes en sus zonas pelágica y litoral (figura 1).
En cada momento y sitio de muestreo se registraron variables fsicoquímicas in situ: temperatura del agua, oxígeno disuelto (DO2 ), porcentaje de saturación del oxígeno disuelto, el pH, el potencial Redox, el dióxido de carbono disuelto, y conductividad eléctrica.
Para la captura de los ejemplares fueron instaladas redes estacionarias durante un período de 48 horas. Cada red presentó diferentes paños (4, 5, 6, 9, 12 y 14 cm), cada uno con 8 m2 . Para aumentar la probabilidad de captura de diferentes especies y tamaños, fueron utilizados otros aparejos de pesca (atarrayas con tamaños diferentes de malla: 1, 2 y 3 cm; líneas de anzuelo y redes de mano). Para los análisis comparativos, se utilizó solo la información proveniente de las redes estacionarias.
Cada ejemplar capturado fue pesado y medido en sus longitudes estándar y total, posteriormente fjado en solución de formaldehído 10%. Para la determinación de su especie fueron utilizadas claves taxonómicas, descripciones y las listas de Nelson (1994), Vari (1989), Dahl (1971), Eigenmann (1922, 1912) y específcas como Gery (1977) y Schultz (1944). Se siguió la clasifcación de Reis et al. (2003).
Análisis de la información. Fue defnida la estructura primaria de las asociaciones basándose en la composición, su distribución de abundancia en cada uno de los momentos de muestreo (Simpson, 1949). El número de especies esperada a partir de los datos fue estimada con base en curvas de rarefacción, y los estimadores de riqueza de Chao y Jacknnife. Además fueron calculados los descriptores emergentes de la asociación [diversidad de dominancia (Shannon y Weaver, 1948), equidad de Pielou (1969)]. Para verifcar si estos descriptores presentaron diferencias signifcativas entre momentos y sitios de muestreo, se utilizaron análisis paramétricos de varianza de una vía y para defnir si las poblaciones (abundancia y masa) se distribuían espacio–temporalmente, fue realizada una prueba de varianza de una vía para la ''Captura por Unidad de Esfuerzo'' (CPUE, g/m2 /h ó ind./m2 /h).
La infuencia de las variables ambientales sobre la estructura de las asociaciones de peces fue explorada a través de un análisis multivariado directo (redundancia canónica). Para defnir la significancia de la interacción se realizó una prueba de permutaciones (Monte–Carlo con 499 permutaciones). Todos los análisis fueron realizados sobre los datos centrados  y sometidos a una transformación de raíz cuadrada (√x + 1). Para todas las pruebas fueron utilizados los paquetes STATISTICA 6.0 y Canoco® 4.5 para ''Windows®''. Para todos los análisis se asumieron niveles de signifcancia del 95% (α < 0,05).
y sometidos a una transformación de raíz cuadrada (√x + 1). Para todas las pruebas fueron utilizados los paquetes STATISTICA 6.0 y Canoco® 4.5 para ''Windows®''. Para todos los análisis se asumieron niveles de signifcancia del 95% (α < 0,05).
RESULTADOS
Composición y abundancia de especies. En total (incluyendo las capturas con todos los aparejos de pesca) fueron capturados 2.073 individuos, pertenecientes a 23 especies (tabla 1). Las especies C. magdalenae (viejita) y P. blochii (barbudo) aportaron el 71,93% de la abundancia total, mientras que las 14 especies adicionales de las capturadas con las redes de espera representan el número y biomasa restantes.
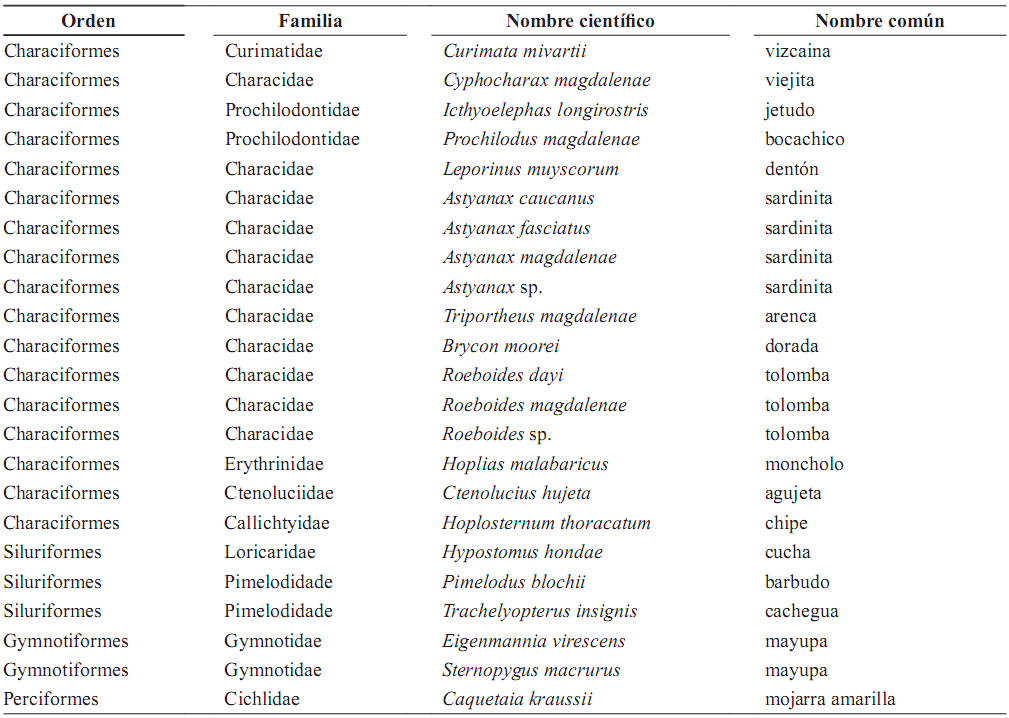
Tabla 1. Especies capturadas en la Laguna de Cachimbero, Cimitarra (Santander), Colombia durante marzo, mayo, octubre y diciembre de 2003 y abril de 2004
Con las redes de espera fueron capturadas 20 especies con 1.372 ejemplares. De estas capturas el mayor porcentaje corresponde a Characiformes 671 individuos (48,90%), Silurifomes 540 (39,35%), Perciformes 146 (11,37%) y Gymnotiformes 5 (0,36%). Las especies C. magdalenae, P. blochii, C. kraussii (mojarra), P. magdalenae (bocachico), T. insignis (cachegua) y Triportheus magdalenae (Arenca) presentaron mayores abundancias en las capturas (tabla 2). Estas seis especies agrupan el 79,39% de la biomasa y el 94,24% del número de individuos.
El mayor número de especies se capturó en la zona litoral del sitio Complejo (n = 13) y el menor en la zona pelágica del sitio Tierradentro en marzo (n = 2). A pesar de que la curva de rarefacción no alcanzó su asíntota en la mayoría de los sitios (figura 2), los estimadores de riqueza Chao y Jack–Knife sugieren que el número de especies esperado en las capturas con las redes de espera es de 20,5 y 19, respectivamente.
Diversidad, dominancia y equidad. Los valores de diversidad por muestreo no superaron los 0,8 bits. La equidad alcanzó valores altos en algunos muestreos (en marzo de 2003 en el sitio Hondonada –zona pelágica–). Los sitios Complejo y Entrecaños presentaron las mayores equidades (E > 0,70) pero los valores de diversidad no superaron los 0,59 bits.
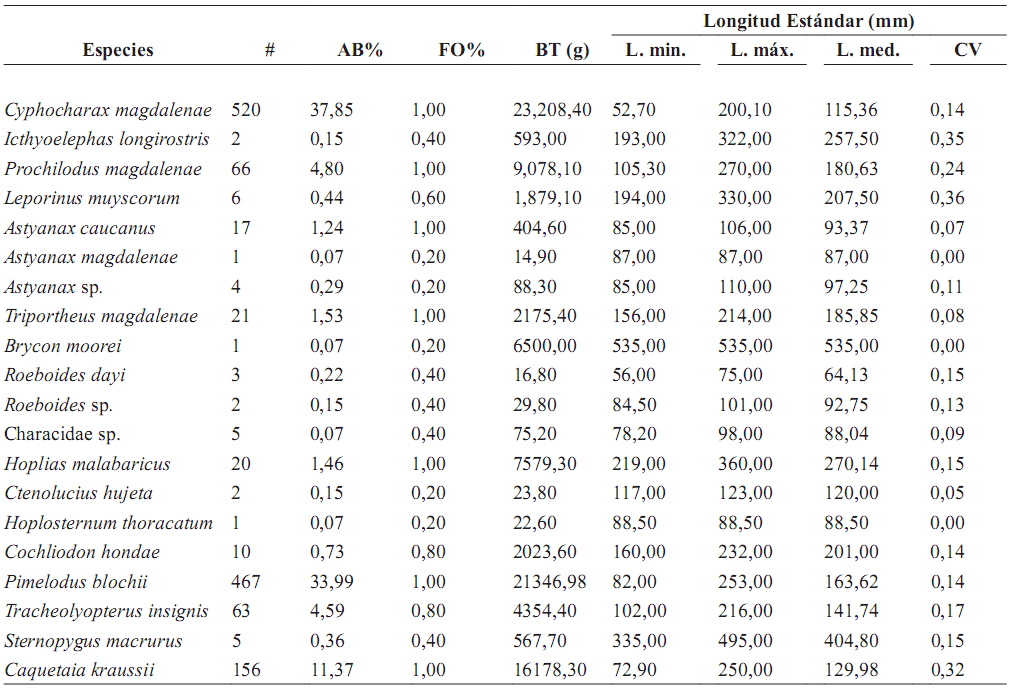
Tabla 2. Biomasa total y talla media de cada una de las especies de peces capturadas en la Laguna de Cachimbero, Cimitarra (Santander), Colombia durante marzo, mayo, octubre y diciembre de 2003 y abril de 2004 [# = número de individuos; AB% = abundancia relativa (%); FO% = Frecuencia Ocurrencia mensual (%); BT = Biomasa Total; L = Longitud mínima, máxima, media; CV = Coefciente de variación]
Distribución espacio–temporal de la abundancia y de la biomasa. La biomasa no presentó diferencias signifcativas entre los factores analizados (mes: H(4,40) = 2.098; p = 0,72; sitio: H(3,40) = 7,84; p = 0,0492; ambiente: H(1,40) = 1.903 p = 0,17) (tabla 3).
Las capturas mayores en número y biomasa de íctiofauna en la Laguna de Cachimbero se registraron para P. blochii (0,693 g/m2 /h), C. magdalenae (0,193 g/m2 /h) y C. kraussii (0,183 g/m2 /h).
Infuencia de las variables ambientales sobre la conformación de la asociación de especies. Los tres primeros ejes reúnen el 99,5% de la variabilidad acumulada entre las especies y el ambiente (figura 3 y tabla 4). No se encontró una relación signifcativa entre las características ambientales de la Laguna de Cachimbero y la asociación de especies de peces (F = 1,8; p = 0,16; tabla 4). Dentro del análisis, C. magdalenae, la profundidad y la temperatura del agua fueron las variables que aportan más información al componente uno.
DISCUSIÓN
La distribución del número de especies de acuerdo con su orden coinciden con la tendencia general en los ecosistemas limnéticos del neotrópico, que se caracterizan por un predominio de formas de los grupos Characiformes y Siluriformes (Lowe– McConnell, 1987). La totalidad de las especies identifcadas durante el estudio son típicas de la cuenca del Magdalena, presentan una amplia distribución en este sistema y han sido reportadas por Mojica (1999) y Dahl (1971).
A pesar de presentar cambios espacio–temporales en sus abundancias Cyphocharax magdalenae, P. blochii, C. kraussii y T. insignis, siempre fueron las especies más importantes en las capturas. Excepto P. blochii, estas especies no migran por el cauce principal del río Magdalena, por lo que las variaciones en su abundancia pueden estar relacionadas con la infuencia que tiene la expansión y contracción del cuerpo de agua sobre el movimiento de los individuos y sobre la efectividad de las redes de pesca.
La laguna de Cachimbero presentó un número bajo de especies y una comunidad de peces poco diversa en comparación con los reportes para otras ciénagas del río Magdalena (Ríos, 2005;
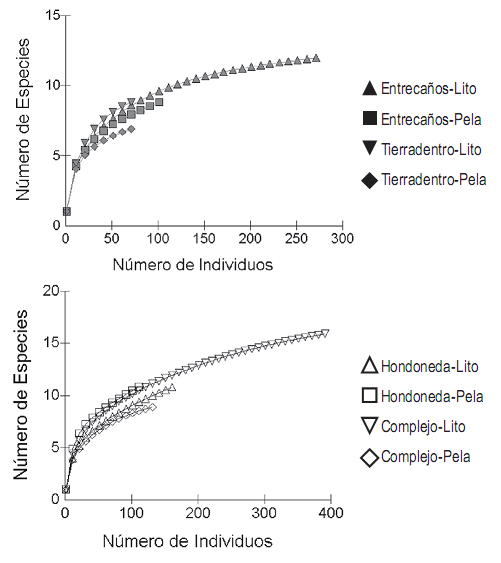
Figura 2. Número de especies esperadas para las estaciones de muestreo en los diferentes momentos de colecta en la Laguna de Cachimbero, Cimitarra (Santander), Colombia. A. Estaciones: Entrecaños y Tierradentro. B. Estaciones: Hondonada y Complejo. Ambientes (Lito = litoral, Pela = pelágica)
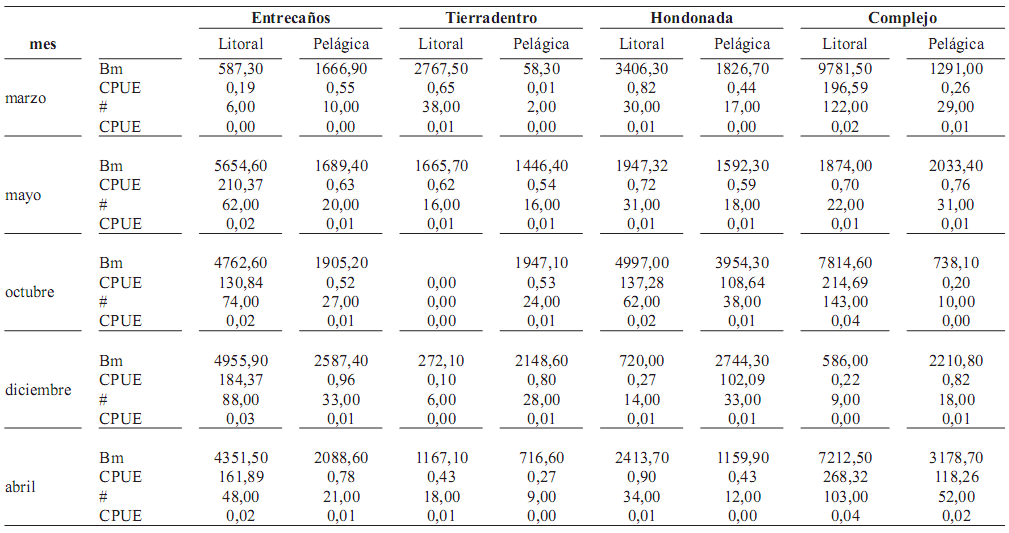
Tabla 3. ''Captura por unidad de esfuerzo'' (CPUE) en número (#) y biomasa (Bm, gramos) con redes de espera en Laguna de Cachimbero, Cimitarra (Santander), Colombia durante marzo, mayo, octubre y diciembre de 2003 y abril de 2004
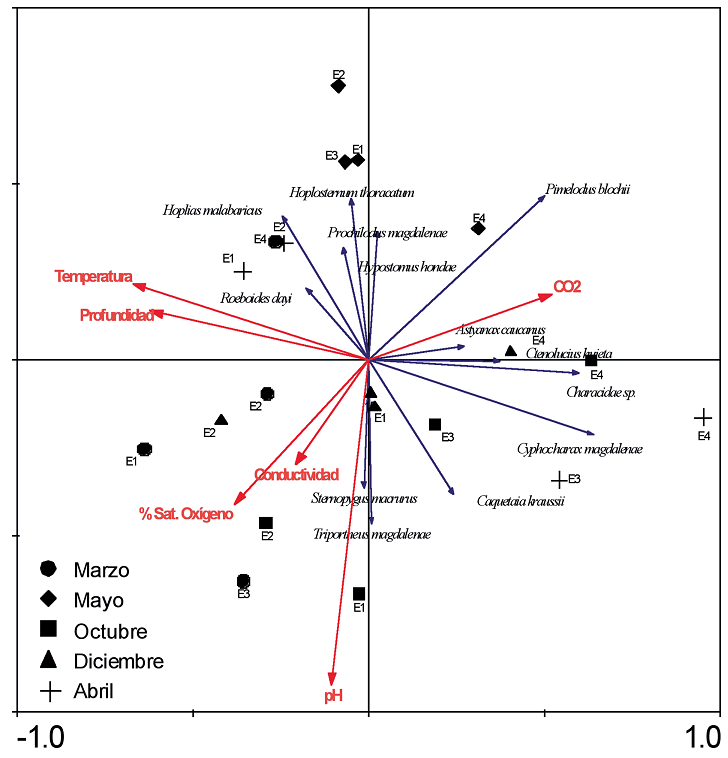
Figura 3. Representación gráfca del Análisis de Redundancia basado en la interacción de las variables ambientales y la asociación de especies en la Laguna de Cachimbero, Cimitarra (Santander), Colombia. Factores de análisis: meses; estaciones (E1 = Entrecaños; E2 = Tierradentro; E3 = Hondonada; E4 = Complejo)
Valderrama y Zárate, 1989). Lowe–McConnell (1975) afirmó que los ambientes donde no se presentan fluctuaciones drásticas en el nivel del agua, la fauna íctica es menos diversa que en los ecosistemas que ofrecen condiciones diferenciales de hábitat y diferentes posibilidades de explotación de los recursos y una mayor coexistencia de los organismos. Por otro lado, la perdida de conectividad con el afluente principal reduce el efecto que tiene el flujo hidráulico y biológico sobre la dinámica propia del sistema generando asociaciones con pocas especies y abundancias poco contrastantes, particularmente en el tiempo.
La estructura de la asociación de especies en la laguna de Cachimbero no tuvo grandes variaciones entre periodos pluviométricos debido a la poca influencia de las especies migratorias (leves cambios en la abundancia de algunas especies, v. g.: Prochilodus magdalenae, Curimata mivartii o bien, con la ausencia de otras, v. g.: Sorubim cuspicaudus y Pseudoplatystoma magdaleniatum). A pesar de que el nivel del agua es el factor abiótico regulador del comportamiento espacial y temporal de la oferta pesquera en el sistema del río Magdalena (Valderrama y Zárate,1989; Zárate y Cubides, 1977), la reducción en la conectividad
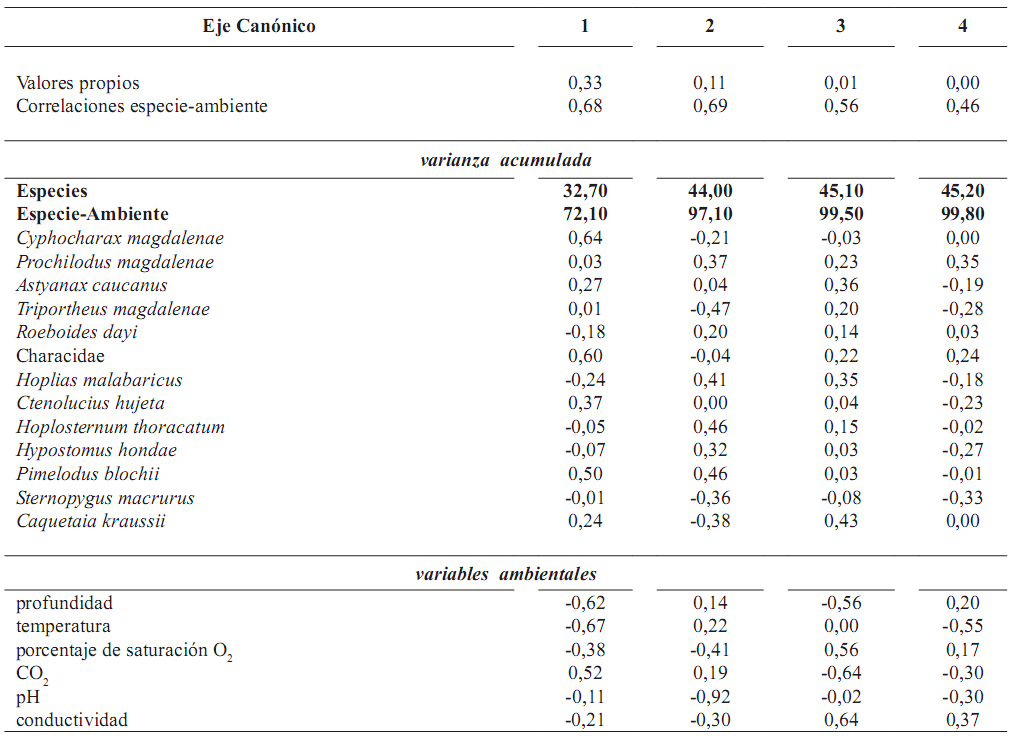
Tabla 4. Análisis de Redundancia realizado con la información de la asociación de especies de peces y las características ambientales en la ciénaga en Laguna de Cachimbero, Cimitarra (Santander), Colombia durante marzo, mayo, octubre y diciembre de 2003 y abril de 2004
hidráulica entre el río Magdalena y la laguna de Cachimbero, ha llevado a la reducción del fujo biótico entre estos ambientes.
La asociación de especies en la laguna de Cachimbero fue particular al hábitat muestreado. La mayor riqueza, abundancia, CPUE y diversidad se observó en la zona litoral y en aquellos sitios próximos a los tributarios y estas características no cambiaron entre los periodos de lluvias y estiaje. La intensidad de la inundación (Junk y Fursh, 1993), la presencia de tributarios, la estructura del suelo y de la vegetación de borde son determinantes en la disponibilidad de hábitats pues potencian la concentración de la biota acuática cerca de las orillas debido a la alta heterogeneidad de hábitat y mayores posibilidades de nichos (Lowe–McConnell, 1987; Machado–Allison, 1987; Machado–Allison y Moreno, 1993).
A manera de conclusión, podemos afrmar que la paulatina perdida de conectividad hidrológica y biológica entre laguna de Cachimbero y el río Magdalena tiene una fuerte infuencia sobre la dinámica de la asociación de especies de peces y que, a pesar de que las lluvias locales provocan cambios en el nivel del agua, este factor ambiental no fue un elemento limitante en la conformación temporal de la comunidad íctica.
AGRADECIMIENTOS
Los autores agradecen el apoyo económico y logístico del Club Rotario–Medellín, a la Universidad de Antioquia, al Fondo CODI–Universidad de Antioquia, a la Fundación Idea Wild, a la Corporación Académica Ambiental y a los integrantes del grupo de Gestión y Modelación Ambiental de la Universidad de Antioquia.
REFERENCIAS
1. Barthem RB, Goulding M. 1997. Os bagres balizadores: ecologia, migração e conservação de peixes amazônicos. Sociedad Civil Mamirauá, Tefé–AM., CNPq. Brasília. Brazil. [ Links ]
2. Bayley PB. 1996. Riverine fishes. Pp. 251–271. En: Petts G, Calow P (eds.). River Restoration. Blackwell Science. Oxford, United Kingdom. [ Links ]
3. Dahl G. 1971. Los peces del Norte de Colombia. Ministerio de Agricultura, INDERENA. Bogotá, Colombia. [ Links ]
4. Eigenmann C. 1912. The freshwater fishes of British Guyana, including a study of species and the relations of the fauna of the plateau to that of the Lowlands. Memoirs of the Carnegie Museum, 5(1):1–578. [ Links ]
5. Eigenmann C. 1922. The fishes of Northwestern South America, including Colombia, Panamá and the Pacific slopes of the Ecuador and Peru, together with an appendix upon the fishes of the río Meta in Colombia. Memoirs of the Carnegie Museum, 9(1):1–346 [ Links ]
6. Galvis G, Mojica JI. 2007. The Magdalena River fresh water fishes and fisheries. Aquatic Ecosystem Health and Management, 10(2):127–139. [ Links ]
7. Gery J. 1977. Characoides of the world. T.H.F. Public. Neptune (NJ), USA. [ Links ]
8. Jiménez–Segura LF. 2007. Ictioplancton y reproducción de los peces en la Cuenca media del Río Magdalena a la altura de Puerto Berrio. Tesis de doctorado en Biología. Universidad de Antioquia. Medellin, Colombia. [ Links ]
9. Junk WJ, Bayley PB, Sparks RE. 1989. The flood pulse concept in river–flooplain system. Canadian Fisheries and Aquatic Sciences (Proceedings of the international Large River Symposium. Special Publication), 106:110–127. [ Links ]
10. Junk W, Fursh K. 1993. A general review of tropical South America floodplains. Wethlenas. Ecology and Management, 2(4):231–238. [ Links ]
11. Lowe–McConnell RH. 1975. Fish communities in Tropical Freshwater: Their distribution, Ecology and evolution. Longman, London. [ Links ]
12. Lowe–McConnell RH. 1987. Ecological studies in tropical fish communities. Cambridge University Press. Cambridge, UK. [ Links ]
13. Machado–Allison A. 1987. Los peces de los llanos de Venezuela un ensayo sobre su historia natural. Universidad Central de Venezuela. Caracas, Venezuela. [ Links ]
14. Machado–Allison A, Moreno H. 1993. Estudio de la comunidad de peces del río Orituco. Estado Guárico, Venezuela. Parte I. Inventario, abundancia relativa y diversidad. Acta Biológica Venezuelica, 14(4):77–94. [ Links ]
15. Mojica JI. 1999. Lista Preliminar de las especies de peces dulceacuícolas de Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 23(Suplemento especial):548–566. [ Links ]
16. Nelson J. 1994. The fishes of the World. 3ra. ed. Jhon Wiley & Sons. NJ, USA. [ Links ]
17. Pielou EC. 1969. An introduction to mathematical ecology. Jhon Willey & Sons. New York, USA. [ Links ]
18. Reis RE, Kullander SO, Ferraris CJ. 2003. Checklist of the freshwater fishes of South and Central America. EdiPucrs. Porto Alegre, Brasil. [ Links ]
19. Restrepo JC, Toro JL. 1997. Reserva Natural Laguna de Cachimbero: Caracterización biofísica. Informe final. Corantioquia, Corporación Laguna de Cachimbero. Medellin, Colombia [ Links ]
20. Ríos MI. 2005. Variación espacio temporal de la comunidad de peces de la ciénaga de Ayapel, Colombia. Tesis de Maestría, Universidad de Antioquia. Medellín (Antioquia), Colombia. [ Links ]
21. Shannon CE, Weaver W. 1948. The mathematical theory of communication. Press Illinois University. Urbana (IL), USA. [ Links ]
22. Schultz LP. 1944. The catfishes of Venezuela, with descriptions of thirty–eight new forms. Smithsonian Institution United States National Museum, 94:173–338. [ Links ]
23. Simpson E. 1949. Measurement of diversity. Nature, 163:688. [ Links ]
24. Valderrama M, Zárate M. 1989. Some ecological aspects and present state of the fishery of the Magdalena river basin, Colombia, South America. Canadian Fisheries and Aquatic Sciences (Proceedings of the international Large River Symposium. Special Publication), 106:209–421. [ Links ]
25. Vari R. 1989. A phylogenect study of the Neotropical characiform family Curimatidae (Pisces Ostariophysis). Smithsonian Contribution to Zoology, 471:1–71. [ Links ]
26. Welcomme RL. 1979. Fisheries ecology of floodplain rivers. London, UK. [ Links ]
27. Zárate M, Cubides A. 1977. Estudio ecológico de las ciénagas del plano inundable del río Magdalena. INDERENA–FAO, Col/71/552. Cartagena (Bolívar), Colombia. [ Links ]
Recibido: julio 2008
Aceptado para publicación: noviembre de 2008

