










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActualidades Biológicas
Print version ISSN 0304-3584
Actu Biol vol.31 no.1 Medellín July/Dec. 2009
ARTÍCULOS DE INVESTIGACIÓN
PATRONES ESPACIO-TEMPORALES DE LA ESTRUCTURA Y COMPOSICIÓN DE LA HOJARASCA EN LAS QUEBRADAS DEL PARQUE NACIONAL NATURAL GORGONA
SPATIO-TEMPORAL PATTERNS OF STRUCTURE AND COMPOSITION OF LEAF LITTER IN STREAMS OF THE GORGONA NATURAL NATIONAL PARK
Sandra M. Valencia-G.1; Giovanny A. Pérez-Z.2; Pilar X. Lizarazo-M.3; Juan F. Blanco4
1 Instituto de Biología, Universidad de Antioquia. A. A. 1226. Medellín, Antioquia, Colombia. smvalencia29@gmail.com
2 Instituto de Biología, Universidad de Antioquia. A. A. 1226. Medellín, Antioquia, Colombia.
3 Docente. Instituto de Biología, Universidad de Antioquia. A. A. 1226. Medellín, Antioquia, Colombia. pixilime@gmail.com
4 Docente. Instituto de Biología, Universidad de Antioquia. A. A. 1226. Medellín, Antioquia, Colombia. jfblanco73@yahoo.com, blanco@matematicas.udea.edu.co.
Resumen
Este trabajo estima la estructura y composición de la tafocenosis de la hojarasca acumulada en el lecho de nueve quebradas de la isla Gorgona en abril y junio de 2008. La pácora (Cespedesia macrophylla) fue la especie dominante en términos de biomasa total en toda la isla, aunque fue más abundante en las quebradas cuyos bosques ribereños fueron deforestados antes de la declaración del parque nacional o están sometidos a disturbios naturales (e.g., caída de árboles y los derrumbes) que frecuentemente abren el dosel. Sin embargo, se registraron más de cien morfoespecies, veintidós de las cuales se registraron en ambos muestreos. La mayor parte de las especies aporta poca biomasa y número de hojas, y por lo tanto no parecen ser funcionalmente importantes. Además de las especies dominantes, la biomasa total, abundancia total (número de hojas) y riqueza de la tafocenosis difiere significativamente entre las quebradas posiblemente como consecuencia del estado sucesional del bosque ribereño. Durante la época de lluvias se redujo significativamente la biomasa de la hojarasca acumulada.
Palabras clave: bosques ribereños, Cespedesia macrophylla, diversidad florística, hojarasca, Isla Gorgona, sucesión secundaria
Abstract
This paper estimates the structure and species composition of the leaf litter standing stock in nine streams at Gorgona Island during April and June, 2008. The pácora (Cespedesia macrophylla) contributed most of the biomass island wide, although it was restricted to riparian areas either deforested before the designation of the island as a protected area or to forest gaps created by treefall and bank failure. Nonetheless, >100 morphospecies were recognized, and 22 occurred during both sampling dates. Most of the species made a minor contribution in number of leaves and total biomass to the leaf litter standing stock and are not seemly important for ecosystem function. Riparian forests in different succesional stages differ in terms of dominant species, total biomass, total abundance, and richness. Leaf litter biomass was significantly diminished during the rainy season.
Key words: Cespedesia macrophylla, floristic diversity, Gorgona Island, leaf litter, riparian zones, secondary succession
INTRODUCCIÓN
Colombia posee extensos bosques húmedos tropicales que se consideran de gran importancia debido a que representan gran parte de la riqueza y biodiversidad mundial, concentrando una enorme cantidad de especies de prácticamente todos los grupos taxonómicos (Comité Técnico Interagencial del Foro de Ministros de Medio Ambiente de América Latina y el Caribe 2000).
Dentro de la región biogeográfica del Chocó sobresale la isla Gorgona, única área insular en el pacífico oriental colombiano, cubierta en más del 80% por bosque húmedo tropical (Aguirre y Rangel 1993, Rangel 2004). Estos bosques del Chocó biogeográfico se caracterizan por su gran biodiversidad y alto grado de endemismo de plantas y otros organismos, pero no han sido extensamente estudiados, siendo poco el conocimiento que se tiene sobre la diversidad florística tanto continental como insular (Rangel 2004, Yockteng y Cavalier 1998).
Se propone que durante el pleistoceno, al bajar el nivel del mar, isla Gorgona se unió al continente (Aguirre y Rangel 1993). Esto explica la similitud de flora y fauna entre la costa y la isla, haciendo de ésta última un área que puede considerarse rica. Sin embargo, por su carácter insular presenta menor número de especies en comparación con los bosques continentales del Chocó biogeográfico (Yockteng y Cavelier 1998). Para la región se han estimado más de 100 y 250 especies por 10.000 m2, y en la isla se estiman 100 especies para un área de 8.000 m2, siendo un área que presenta menor riqueza (Yockteng y Cavelier 1998).
Aunque actualmente es un parque nacional natural, la historia de uso de la isla se divide en tres periodos desde el punto de vista agronómico (Bernal 1989, Arango et al. 1991, manuscritos sin publicar). 1) Colonización (1830-1959): durante este periodo se establecieron cultivos en el sur (principalmente cítricos, coco, maíz, caña de azúcar, plátano y pastos). Hacia el centro y norte las siembras fueron menos intensas, principalmente cocoteros. Se considera que durante este periodo la agricultura fue localizada, de subsistencia familiar y con pocos efectos degradativos. La explotación maderera fue una actividad secundaria. La población en el centro y sur fue ocasional principalmente con fines recreativos. 2) Isla-prisión (1959-1984): durante este periodo hubo gran impacto en los suelos del centro (sitio del penal y ''El Poblado'') y nororiente de la isla. Se incrementó el número de plantas de cultivo por parte de los guardianes y personal administrativo. Se expandió el cultivo de coco y se introdujo yuca, piña, plátanos, coca, café, papaya, palma africana y algunas plantas ornamentales (e.g., crotón, antorcha). Durante este periodo se expandieron las malezas. 3) Parque Nacional Natural (1984- presente): Este periodo se ha caracterizado por: a) la ausencia de colonos (solo existe personal administrativo del parque y de las concesiones eco-turísticas); b) la regeneración natural en el sur de la isla, y la antigua colonia penal de Huisitó y Playa Blanca; c) la reforestación con especies de árboles nativos (machare, tangare, caimito, virola) y palma naidí (introducida) en zonas sensibles a la erosión y derrumbes alrededor de la isla y a lo largo de la quebrada Ilú (= Trinidad) desde la desembocadura (e.g., antigua cancha de fútbol y marranera); d) la concentración de la actividad humana en El Poblado y los senderos ecológicos.
Durante los primeros años posteriores a la declaración del parque se propuso que existía una vegetación intervenida en etapas de sucesión o ruderal que correspondía a: a) los cultivos en el costado oriental y suroriental y cerca a la playa detrás de los cocales y árboles frutales, b) las áreas ''aclaradas'' para la entresaca de madera, principalmente en los alrededores de Yundigua (= Pablo VI) (Rangel 1990). La vegetación natural ''tipo selvática'' (con dos estratos arbóreos) estaba conformada por la alianza Cespedesio-Symphonion, dentro de la cual se encontraban tres asociaciones: a) Ossaeo sessilifoliae-Anaxagoretum phaeocarpae; b) Malpigioglabrae-Cespedesietummacrophyllae y Cassipoureo ellipticae-Ryabetum speciosae. Las dos primeras asociaciones se establecen entre los 40 y 200 m y la otra por encima.
Estas diferencias en la composición de especies podrían generar una contribución diferencial de hojarasca sobre las quebradas de la isla y en consecuencia podría tener efecto sobre las tasas de descomposición de ésta y sobre las condiciones del hábitat y refugio para algunas comunidades biológicas. En los sistemas lóticos de bosques tropicales, la principal fuente de energía está representada por el ingreso de material alóctono, constituido por diferentes tipos de desechos, entre los cuales el material vegetal representa un porcentaje importante (Swan y Palmer 2006), siendo la hojarasca sumergida una fuente de energía y carbono, que queda disponible para macroinvertebrados y otros organismos, posterior a la actividad de la microbiota asociada a ésta (Das et al. 2007). Los modelos de la dinámica del procesamiento de la hojarasca, que establecen que éste se puede ver afectado por varios factores incluyendo la estacionalidad de la caída de hojas (Wantzen et al. 2008), han sido bien documentados y sustentados en estudios para zonas templadas. Sin embargo, dichos modelos pueden no ser aplicables para el trópico, debido a que el ingreso de hojarasca a las quebradas puede ser constante a lo largo del año. Se ha sugerido que en quebradas tropicales y principalmente en las de pequeño orden el papel de la hojarasca en la transferencia de energía hacia los diferentes niveles tróficos puede ser más significativo. La relevancia de la hojarasca en el ecosistema se fundamenta en que esta representa la mayor parte del ingreso de materia orgánica a partir de los bosques ribereños (Abelho 2001).
Por otra parte, la retención y tiempos de residencia de la hojarasca dentro de las quebradas, así como los cambios en la contribución a lo largo del cauce, son factores que afectan de manera directa la dinámica de los procesos de descomposición, a pesar de lo cual estas variables no han sido consideradas en los estudios. Adicionalmente, el procesamiento de la hojarasca no ocurre en un contexto espacio-temporal equilibrado sino más bien en puntos y tiempos específicos (''hot spots'' y ''hot moments'') (Wantzen et al. 2008), lo que hace necesario ampliar el conocimiento que se tiene acerca de los diferentes factores implicados en estos procesos y que en consecuencia, conduzca a un mejor entendimiento de éstos, particularmente para zonas tropicales.
Este trabajo evalúa la variación espacio- temporal de la composición y estructura de la hojarasca sumergida en las quebradas del Parque Nacional Natural Gorgona, lo que se constituye en el primer paso hacia la identificación y entendimiento de algunas de las variables involucradas en la dinámica de estos ecosistemas acuáticos y que pueden condicionar su diversidad funcional y estructural.
MATERIALES Y MÉTODOS
Área de estudio. Isla Gorgona (2° 47' - 3° 06' N, 78° 06' - 78° 18' O) está localizada en el pacifico oriental colombiano a 30 kilómetros del continente (Díaz et al. 2001; ver mapa en Blanco et al. 2009). La parte terrestre de la isla ocupa 1.568 ha, de las cuales el 90% presentan topografía escarpada con pendiente de 50-75%. La temperatura promedio anual del aire es de 26 °C, con una humedad relativa promedio superior al 90% (Aguirre y Rangel 1993). El promedio anual de precipitación es 6.778 mm, con un régimen unimodal [época de menor precipitación entre diciembre y abril, y época lluviosa entre mayo y noviembre (Blanco 2009a)].
Colecta y procesamiento de muestras. Para evaluar la variación en la composición de la hojarasca en las quebradas de la isla, se realizaron muestreos durante la época seca (abril) y la época lluviosa (junio) de 2008. Se optó por muestrear en abril y junio ya que son representativos de cada época y también por limitaciones logísticas y presupuestales para muestrear a lo largo del año. Se eligieron para los muestreos nueve quebradas de las veinticinco permanentes que presenta la isla. Siete quebradas se ubicaron en el costado oriental (Iguapoga, Ilú, Muelle, Aeropuerto Sur, Chorro del Cura, Pizarro y El Roble) y dos en el costado occidental (La Camaronera y Cocal Norte). Para ver una completa descripción de las características ambientales de las quebradas consultar a Gómez et al. (2009, en este volumen). En cada una de las quebradas se muestreó un tramo bajo, de 10 a 25 msnm. El muestreo cualitativo consistió en identificar en cada quebrada un tramo de 10 m de largo (pero el ancho del cauce fue variable) dentro del cual se colectaron manualmente paquetes mixtos de hojarasca sumergida realizando un recorrido en zigzag, con el fin de abarcar la mayor área posible.
El procesamiento de las muestras incluyó el prensado, secado y posterior separación de la hojarasca por morfotipos. Se realizó la cuantificación de número de hojas y de biomasa en peso seco por morfotipo en cada quebrada. A cada morfotipo se le tomó una fotografía digital y se le adjudicó un código. Para la identificación de morfotipos se siguió el listado de plantas de la isla de Gorgona publicado por Rangel (1990) y posteriormente se buscaron imágenes en catálogos digitales en línea del Instituto Smithsoniano de Investigaciones Tropicales en Panamá (STRI 2009 a, b, c: Herbario, Flora de Barro Colorado, Flora de Bocas del Toro) y del Jardín Botánico de Missouri (MOBOT 2009 a, b, c: Flora Tropical, Lista de chequeo del río Escalerete-Valle del Cauca, Lista de chequeo del Bajo Calima-Valle del Cauca). También se consultó a los botánicos del Herbario de la Universidad de Antioquia.
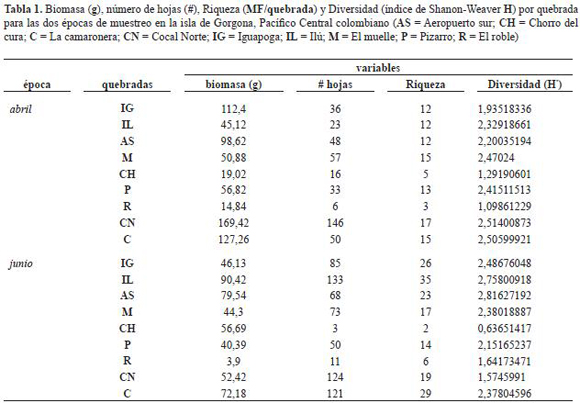
Procesamiento de la información. A partir de la información anterior se construyeron matrices de datos que fueron utilizadas para la realización de los gráficos y análisis estadísticos. Utilizando la biomasa (peso en gramos del paquete de hojas) y el número de hojas totales para cada morfotipo para toda la isla, se construyó una curva de especies ranqueadas para determinar las más importantes con respecto a estas dos variables. Posteriormente se realizó una regresión lineal para establecer si el aporte en número de hojas era proporcional al aporte en biomasa. Para las especies dominantes se graficó la biomasa y número de hojas en todas las quebradas para establecer patrones espaciales de composición. Finalmente, se calculó la diversidad utilizando el índice de Shannon-Weaver (H') para cada quebrada, y junto con la biomasa total (gramos), abundancia total (número de hojas), riqueza (número de especies) y la biomasa y abundancia de los morfotipos más importantes (4, 5, 10, 15, 19, 21), con estos datos se construyó una matriz para explorar patrones espacio-temporales de la estructura y composición de la tafocenosis. Se incluyó la información de los muestreo en tabla 1. Esta matriz se utilizó para poder ordenar las quebradas basadas en las características de la tafocenosis, por medio de un Escalamiento Multidimensional no-Métrico (NMDS), utilizando como distancia de medida Sorensen (Bray-Curtis). Las coordenadas de ordenación fueron correlacionadas con la matriz de datos para determinar las variables que mayor relación guardan con los ejes. Estos análisis fueron realizados utilizando el programa PCORD (versión 4.25).
RESULTADOS
A partir de las colectas de hojarasca sumergida en las quebradas de la isla Gorgona, en el muestreo realizado en abril se obtuvieron 70 morfotipos y en junio 101; 22 morfotipos fueron comunes a las dos épocas. En abril el peso seco total de hojarasca, de los morfotipos colectados fue de 694,4 g, de los cuales el 35,0% correspondió al morfotipo 2 (pácora o pacó; Ochnaceae: Cespedesia macrophylla) (figura 1), seguida por el morfotipo 14 (indeterminado) con 6,5% y los morfotipos 29 (yarumo; Moraceae: Cecropia garciae), 26 (indeterminado) y 15 (Ficus tonduzii) con valores inferiores. En junio la biomasa total fue de 486,0 g, de los cuales C. macrophylla contribuyó con el 35,0%, el morfotipo 4 (higuerón; Moraceae: Ficus sp.) con el 13,7% y el morfotipo 10 (mantequillo; Euphorbiaceae: Sapium sp.) con el 6,2%. Los morfotipos 11 (árbol del pan; Moraceae: Artocarpus altilis) (figura 1) y el 101 contribuyeron seguidamente con menos de este valor. El morfotipo más importante en relación al aporte en biomasa en las dos épocas estudiadas fue C. macrophylla, los otros morfotipos presentaron aportes menores o cambiaron con la época de estudio (figura 2).


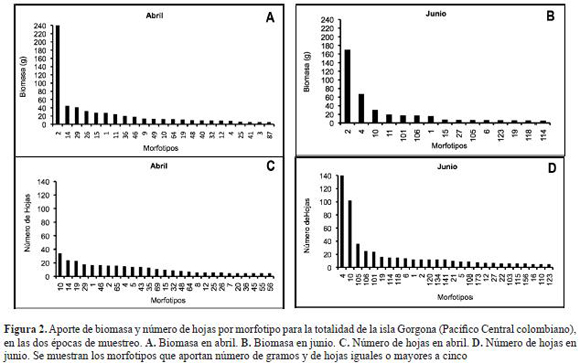
Con respecto al número de hojas, en abril se colectaron en total 417 hojas de las cuales el morfotipo 10 (mantequillo: Sapium) contribuyó con 8,2%, el morfotipo 14 con 5,8%, el morfotipo 19 (Heliocarpus) con 5,2% y el morfotipo 29 con 4,3%. En junio se colectaron 668 hojas, el mayor aporte lo representó el morfotipo 4 (higuerón: Ficus sp.) con 21,0%, seguido del morfotipo 10 (mantequillo: Sapium) con 15,26%, el morfotipo 105 con 5.38% y los morfotipos 106 y 101 con menos del 4%. El mantequillo (Sapium) fue el representante común, además con un porcentaje mayor de contribución en número de hojas en las dos épocas muestreadas (figura 2). Otras especies secundariamente importantes en términos de biomasa, (que pudieron ser identificadas), pero poco frecuentes y abundantes son: Majagua (Malvaceae: Hibiscus glabrata), Otobo (Myristicaceae: Dialyanthera), Bauhinia (Fabaceae), Terminalia (Combretaceae), Pienemono, Balso o Corcho (Tiliaceae: Heliocarpus), Sajo (Anacardiaceae: Camnosperma panamensis), Cuángare (Myristicaceae: Virola), Machare (Clusiaceae: Symphonia globulifera), Tángare (Meliaceae: Carapa guianensis), Caimitillo (Sapotaceae: Pouteria euguiifolia), Lechero, Lecheperro o Sandé (Brosimun utile), Guayabillo (Myrtaceae), Miconia (Melastomataceae).
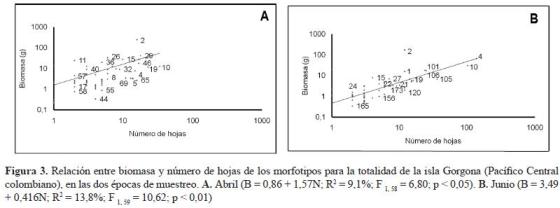
Se observó una correlación significativa (pero con una alta dispersión) entre el número de hojas y la biomasa para las especies en ambas épocas (figura 3). La mayoría de las especies aportan poca biomasa aunque el número de hojas/especie sea variable. En la figura 3 se observa que tanto en abril como en junio C. macrophylla (morfotipo 2) aportó la mayor biomasa a pesar de que no aportó muchas hojas.
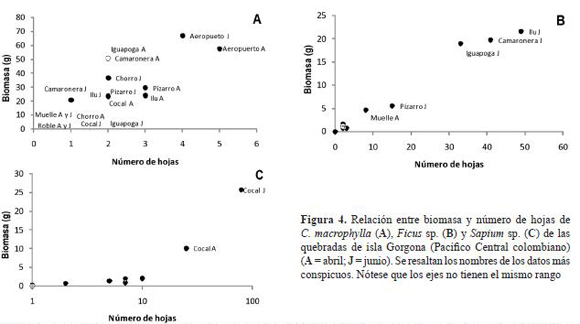
La figura 4 muestra la distribución espacial de la biomasa y número de hojas de las tres especies dominantes. Cespedesia macrophylla contibuyó con la mayor biomasa y número de hojas en el Aeropuerto Sur, seguida por un grupo conformado por La Camaronera, Pizarro, Ilú, Igauapoga y El Chorro del Cura. En El Roble y El Muelle no se encontró la especie. Ficus presentó la mayor abundancia y biomasa en el mes de junio en Ilú, La Camaronera e Iguapoga. Sapium tuvo la mayor abundancia y biomasa en Cocal Norte.
La ordenación de las quebradas en las dos épocas basadas en la composición de morfotipos más abundantes y frecuentes (biomasa y número de hojas de los morfotipos 1, 2, 4, 5, 10, 15, 19, 21) y los índices de estructura de la tafocenosis (tabla 1) mostró 3 grupos a lo largo del eje 2 (figura 5). Este eje se correlacionó con la biomasa, número de hojas, riqueza y diversidad. Con el eje 2 también se correlacionaron las biomasas de Vismia baccifera, Ficus sp., Heliocarpus y el morfotipo 21. El eje 1 corresponde a la biomasa de dos especies de Ficus.

DISCUSIÓN
Patrones de riqueza. Este estudio demuestra que la riqueza de morfotipos de la tafocenosis de la hojarasca es alta en las dos épocas de estudio (abril: 70; junio: 101). Esto es reflejo de la alta riqueza de especies de árboles estimada para los bosques de la isla [> 100/8.000 m2: Yockteng y Cavelier (1998)]. Lo que también es soportado por el hecho de que solamente 22 morfotipos fueran comunes entre las dos épocas, sugiriendo que hay un alto recambio temporal en las especies que aportan hojarasca a las quebradas, aumentando significativamente la riqueza. De hecho hemos encontrado alrededor de 180 morfotipos considerando partes bajas y altas de las nueve quebradas estudiadas (datos sin publicar). Estudios en el Bosque Experimental de Luquillo en la isla de Puerto Rico han mostrado un gran recambio temporal de especies en la hojarasca debido a que cada especie de árbol tiene una fenología particular de caída de hojas, flores y frutos (Zalamea y González 2008). Aunque el estudio de morfotipos puede sobre- o sub-estimar la riqueza taxonómica de la flórula de una región, el valor encontrado en la isla Gorgona se encuentra dentro del rango reportado para bosques del Chocó Biogeográfico (Rangel 2004). Los índices de diversidad (H = 3,9 bits/individuo) y de equidad (J = 0,86) de árboles son muy altos en la isla Gorgona, siendo similares a los encontrados en otros bosques continentales del pacífico colombiano y ecuatoriano (Yockteng y Cavelier 1998). Esto se debe a que las especies raras contribuyen a aumentar la diversidad. Para reducir este efecto de las especies raras, al considerar solamente las especies más abundantes en la hojarasca se encuentra que la diversidad es menor (H > 2 bits/individuo).
Finalmente, aunque se han realizado varios inventarios florísticos en la isla desde 1983 éstos no han sido intensivos, ni extensos (Cabrera 1983, Fernández-Pérez 1985, Murillo y Lozano 1989, Palta 1986). El inventario más extenso y detallado fue el realizado por Rangel (1990) y Rangel et al. (1990a) en 1986 y 1987. Hasta el momento no se han muestreado sistemáticamente los bosques ribereños los cuales podrían contener especies exclusivas como se ha encontrado en otras partes del mundo (Heartsill-Scalley et al. 2009, Naiman et al. 2005). Una fase posterior de este estudio considerará la visita a los herbarios donde se mantiene la mayor cantidad de muestras botánicas de la isla Gorgona (Herbario Nacional, Herbario de la Universidad del Valle, de la Universidad del Cauca y de la Fundación Universitaria de Popayán).
Composición específica. La pácora (C. macrophylla) fue la especie más frecuente y con mayor aporte de biomasa en ambas épocas (35%), aunque aporta pocas hojas. El primer patrón puede ser el reflejo de su dominancia en estados sucesionales tempranos y que puede alcanzar el estrato arbóreo en los claros de bosques maduros. Es dominante en los rastrojos y bosques secundarios jóvenes de la vertiente oriental. También es común (aunque no abundante) en los bosques maduros de la vertiente occidental donde han ocurrido caídas de árboles o deslizamientos de los bancos de las quebradas. Su gran aporte en biomasa se debe al gran tamaño (hasta 1 m de longitud) que alcanzan sus hojas. Anteriormente se había reportado que ésta era una especie pionera dominante en los rastrojos jóvenes (< 5 años; Cabrera 1983) cercanos a El Poblado poco después de la declaración de la isla como parque nacional natural. Actualmente representa la principal especie arbórea emergente en las zonas donde antiguamente estuvieron el aeropuerto, el muelle, el cementerio, la cancha de futbol (actual estación meteorológica) y al norte de ''El Chorro del Cura'' a ambos lados del camino a Playa Pizarro (figura 6). Funcionalmente, ésta especie podría ser muy importante no solo porque contribuye con la mayor cantidad de hojarasca a las quebradas, sino que por su tamaño contribuye a retener hojas de especies más pequeñas y a formar ''paquetes''. En general, el lecho de los rápidos almacena poca hojarasca posiblemente por el régimen de disturbio (JF Blanco observaciones personales) y el mayor almacenamiento se da en las piscinas o charcos. Nuestros resultados, aunque no cuantificaron el almacenamiento por unidad de área, si muestran que la biomasa total se redujo en un 43%, a pesar de que el número de hojas incrementó en un 37%. Esto también sugiere que el aporte de especies de hojas pequeñas (con poca biomasa) no alcanza a compensar el arrastre de la hojarasca dentro del cauce durante la época de lluvias, ya que no se observó una correlación entre el número de hojas totales y la biomasa total por especie. En términos generales, las especies aportan igual proporción de hojas, pero entre ellas la más abundante fue el mantequillo (Euphorbiaceae: Sapium), dominante en ambas épocas en las quebradas Pizarro y Cocal Norte. El higuerón (Ficus) fue secundariamente importante en número y biomasa en varias quebradas y se relaciona directamente con el desarrollo del bosque ya que aunque es una especie pionera en los claros de bosque permanece en estados sucesionales tardíos. Vismia y Heliocarpus tienen una estrategia similar. El árbol del pan (Moraceae: Artocarpus) aunque está restringida al área de El Poblado aporta hojas muy grandes al cauce de la quebrada Ilú.

Todas las especies secundarias que pudieron ser determinadas son características de los bosques del pacífico colombiano (Rangel 1990, 2004). Sorprendentemente, el yarumo (Moraceae: Cecropia) tuvo una baja abundancia y frecuencia de árboles y por lo tanto un aporte insignificante de hojarasca comparado con otros bosques neotropicales (Foster y Brokaw 1992). Esta especie es considerada la pionera por excelencia en esta región pero podría estar siendo excluida por la pácora en los bosques de la isla Gorgona, posiblemente debido al tipo de suelo arcilloso pobre en materia orgánica (Palta 1986, JF Blanco observaciones personales).
Patrones espaciales. Este estudio sugiere que las diferencias en composición de especies que presentan los bosques ribereños se reflejan en la hojarasca acumulada en las quebradas. Además tres variables estructurales separan los sitios: biomasa total, abundancia total y riqueza. Los sitios con mayor riqueza además tienen mayor abundancia. La baja biomasa en la quebrada El Roble puede ser explicada por el alto régimen de disturbio hidrológico, lo cual también se refleja en las características físico-químicas del agua y el ensamblaje de macroinvertebrados (Blanco 2009b, Gómez et al. 2009, en este volumen). La zona ribereña está dominada por árboles de Buchenavia sp. conocida localmente como ''roble''.
El Chorro del Cura también presentó poca biomasa probablemente porque está rodeada por un rastrojo con dosel abierto en el que Cecropia garciae y Miconia spp. sobresalen entre la densa capa rastrera de helechos marraneros (Pteridium aquilinum) (figura 6). Los funcionarios del parque mencionan anecdóticamente, que en esta zona se establecieron cultivos bajos durante la época de la prisión y según Cabrera (1983) era un rastrojo de aproximadamente 5 años al momento de la declaración de la isla como parque.
Aunque la biomasa total es la principal variable que separa las quebradas estudiadas, esta fue dominada por diferentes especies. C. macrophylla fue dominante en El Aeropuerto, Iguapoga, Ilú y La Camaronera. Ficus fue dominante en junio en Iguapoga, Ilú y La Camaronera. Terminalia fue dominante en Iguapoa e Ilú. Las dos primeras especies son consideradas como pioneras que pueden permanecer en los bosques secundarios (Palta 1986). En Cocal Norte, Sapium y Heliocarpus tuvieron la mayor biomasa. El Muelle y Pizarro tuvieron biomasas totales intermedias pero diferentes especies dominantes. En Pizarro la pácora fue secundaria en términos de biomasa.
De acuerdo a la literatura gris, algunas publicaciones (Cabrera 1983, Palta 1986) y a anécdotas de los funcionarios más antiguos del parque se hipotetiza que algunos bosques son secundarios debido a la intervención humana. Se menciona que la vertiente oriental fue intervenida con tala rasa o entresaca desde unos 50-60 m desde la línea de marea hasta los 50-100 msnm dependiendo de la zona (Palta 1986). En Ilú e Iguapoga es evidente el legado de la deforestación pasada, alrededor del penal y de El Poblado, por la presencia de especies exóticas tales como cocoteros (Cocos nucifera), aguacate (Persea americana), totumo o calabazo (Crescentia cujete), naranja (Citrus sinensis), mamey (Mammea americana), mango (Manguifera indica), guamo (Inga edulis) y árbol del pan (Artocarpus altilis). En el área hubo una cancha de futbol, una marranera, una plantación de árboles frutales, rosas y un camino de piedra para subir a los ''montes'' a extraer madera (Rangel et al. 1990a). En Pizarro, se presume que hubo explotación de maderables como el sajo (Campnosperma panamensis), el sande (Brosimum utile) y el tángare (Carapa guianensis) durante la época del penal. La zona ribereña de la quebrada El Aeropuerto tiene una vegetación de crecimiento secundario (rastrojo bajo-medio), donde se observan especies introducidas, mezcladas con helecho marranero y árboles emergentes de especies pioneras (pácora y yarumo), ''helecho arbóreo'' (Cyathea sp.) emergente y cocoteros. En la quebrada El Muelle, la cual bordea por detrás a la ladera que corre paralelamente el área del antiguo muelle y el cementerio, la zona ribereña presenta especies introducidas (frutales y cocoteros) y son dominantes el helecho marranero y pácoras emergentes. También se menciona que en los primeros años de operación del parque se realizaron esfuerzos para reforestar algunas áreas con especies nativas.
Los bosques primarios antes de la declaración del parque se encontraban en las partes altas (>100 msnm) y la vertiente occidental, según Palta (1986), quien hizo colectas a lo largo de nueve transectos altitudinales cruzando la isla, recorridos alrededor de la franja costera y a lo largo de algunas quebradas entre 1975 y 1976. Según Cabrera (1983) algunos de los árboles más gruesos (diámetro a la altura del pecho) de estos bosques eran: Manilkara (Sapotaceae) con 200 cm, Apeiba (Tiliaceae) con 100 cm, Cordia (Boraginaceae) con 80 cm, Tapirira (Anacardiaceae) con 45 cm y Aspidosperma (Apocynaceae) 40 cm, y alcanzaban alturas superiores a los 30 m. Las cabeceras de todas las quebradas en nuestro estudio presentaron características similares en la altura del dosel y diámetro de los árboles, pero además notamos una alta incidencia de derrumbes y volcamiento de árboles lo que produce muchas áreas de claros, lo cual fue evidente en Pizarro y La Camaronera, probablemente incrementado por las rocas sedimentarias, suelos poco desarrollados y pendientes fuertes. Palta (1986) ya había observado que en los bosques maduros, a pesar de la ausencia de actividad antrópica, la acumulación de hojarasca, los suelos anegados y las raíces superficiales producían volcamiento de arboles de 35 m de alto, ayudado por la acción de los vientos.
Implicaciones funcionales. Los anteriores patrones de composición y distribución de las especies de hojarasca en el lecho de las quebradas pueden tener unas implicaciones importantes para el funcionamiento ecosistémico. Fue posible observar paquetes de hojarasca monoespecíficos (e.g.,: pácora, higuerón, mantequillo, árbol del pan, sandé) o mixtos (> 5 especies). Estas diferencias no representan únicamente composiciones diferentes sino diferentes calidades. Por ejemplo, la pácora es de baja calidad (C: 41%, N: 1%, P: < 0,1%; JF Blanco, datos sin publicar), es muy dura y permanece intacta durante mucho tiempo en las quebradas (recalcitrante) (figura 1). Adicionalmente, según Rangel et al. (1990a, b) las hojas son predominantemente esclerófilas independientemente del tamaño (mesófilas o macrófilas), lo que significa que son densas y con poco contenido de agua. Por lo tanto, incluso los paquetes mixtos podrían tener mala calidad desde el punto de vista físico. Entre los paquetes monoespecificos también sobresalen los del árbol del pan (especie introducida) la cual tiene una biomasa foliar especifica similar a la de la pácora y es dominante en la parte baja de Iguapoga y aunque se desconoce su calidad podría alterar el procesamiento de la hojarasca en las quebradas. Aunque desconocemos cual podría ser la consecuencia para el ecosistema acuático de los cambios cuantitativos y cualitativos de la hojarasca, Kominoski et al. (2007, 2009) encontraron recientemente en estudios experimentales en Coweeta (Carolina del Norte, EUA) que paquetes de hojarasca que contenían una mayor riqueza de especies se descomponían más rápido y no se observaron efectos aditivos. La presencia de especies particulares también tenía un efecto sobre las tasas. Por otra parte, la riqueza de especies en los paquetes de hojarasca también condicionó los macroinvertebrados fragmentadores presentes y aunque la composición de especies de la hojarasca no tiene un efecto fuerte, la presencia de algunas especies de mala calidad puede alterar significativamente la composición de los macroinvertebrados.
En conclusión, la deforestación pasada en la isla Gorgona dejó legados en los nuevos bosques a través de: a) reducción de la diversidad de hojarasca, b) el aumento de la dominancia de especies pioneras tales como pácora, balso, higuerón y Miconia (posiblemente de baja calidad físico-química), y c) el reemplazo de especies nativas por especies exóticas tales como el árbol del pan y otros frutales. Lo anterior podría tener implicaciones sobre el funcionamiento de las quebradas. Para avanzar en el entendimiento de la relación funcional bosque-quebrada en el Chocó biogeográfico y particularmente en isla Gorgona recomendamos la realización de estudios sobre: a) la caracterización florística de los bosques ribereños en función de actividades humanas presentes o pasadas, b) fenología de la caída de la hojarasca, c) la calidad físico-química y tasas de descomposición de la hojarasca de especies nativas y exóticas dominantes, d) fauna asociada al proceso de descomposición de la hojarasca, e) manipulaciones experimentales para determinar el papel de propiedades particulares de los paquetes de hojarasca (e.g., calidad, diversidad taxonómica, identidad/origen) sobre las comunidades de macroinvertebrados y el funcionamiento de los ecosistemas.
AGRADECIMIENTOS
Agradecemos al Comité Central de Apoyo a la Investigación (CODI) de la Universidad de Antioquia por la financiación del Proyecto Gorgona Island Stream Bio-Assessment (GISbIO). A la Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales por permitirnos realizar esta investigación en la isla Gorgona. A los integrantes del grupo de trabajo del Proyecto GISBIO. A Juan Carlos Arias, Magnolia Longo, Ana María Gómez Aguirre, Justino Bonilla y Héctor Javier Montaño quienes participaron en el trabajo de campo. Agradecemos también al Herbario de la Universidad de Antioquia por el procesamiento del material vegetal y al Laboratorio de Micología por el préstamo de sus instalaciones para la determinación de los morfotipos. Finalmente, agradecemos los comentarios de dos evaluadores anónimos que contribuyeron a mejorar el manuscrito.
REFERENCIAS
1. Abelho M. 2001. From litter fall to breakdown in streams: a review. The Scientific World, 1: 656-680. [ Links ]
2. Aguirre J, Rangel O. 1993. La isla Gorgona y sus ecosistemas. En: Leyva P, editor. Colombia Pacífico. Tomo I. Bogotá (Colombia): Fondo para la Protección del Medio Ambiente -FEN- Colombia. p. 106-170. [ Links ]
3. Blanco JF. 2009a. The hydroclimatology of Gorgona Island: seasonal and ENSO-related patterns. Actualidades Biológicas, 31(91): 111-121. [ Links ]
4. Blanco JF. 2009b. Características físico-químicas de las quebradas del Parque Nacional Natural Gorgona. Actualidades Biológicas, 31(91): 123-140. [ Links ]
5. Cabrera I. 1983. Inventario florístico preliminar de la isla de Gorgona. Boletín CAE (Corporación para la Acción Ambiental, Universidad del Valle), 1983: 11-20. [ Links ]
6. Comité Técnico Inter-Agencial del Foro de Ministros del Medio Ambiente de América Latina y el Caribe. 2000. Programa de las Naciones Unidas para el Medio Ambiente. Fecha de consulta: 18 de agosto 2009. Disponible en: http://www.pnuma.org [ Links ]
7. Das M, Royer T V, Leff LG. 2007. Diversity of fungi, bacteria and actinomycetes on leaves decomposing in a stream. Applied and Environmental Microbiology, 73: 756-767. [ Links ]
8. Díaz JM, López M, Barrios LM. 2001. Introducción. En: Barrios LM, López M, editores. Gorgona marina: contribución al conocimiento de una isla única. Serie Publicaciones Especiales No. 7. Santa Marta (Colombia): INVEMAR. p. 13-16. [ Links ]
9. Fernández-Pérez A. 1985. Primer catálogo de las plantas de las islas de Gorgona y Gorgonilla. En: Inderena- Cauca, editores. Gorgona: Parque Nacional Natural, No. 1. Programa la universidad en los parques. Popayán (Cauca): INDERENA-Cauca y Fundación Universitaria de Popayán. [ Links ]
10. Foster R, Brokaw NV. 1992. Estructura e historia de la vegetación de la isla de Barro Colorado. En: Leigh E, Rand AE, Windsor DM, editores. Ecología de un bosque tropical. Ciclos estacionales y cambios a largo plazo. Balboa (Panamá): Smithsonian Tropical Research Institute. p. 113-127. [ Links ]
11. Gómez-Aguirre AM, Longo-Sánchez MC, Blanco JF. 2009. Macroinvertebrate assemblages in Gorgona Island streams: spatial patterns during two contrasting hydrologic periods. Actualidades Biológicas, 31(91):161-178. [ Links ]
12. Heartsill-Scalley T, Crowl TA, Thompson J. 2009. Tree species distributions in relation to stream distance in mid-montane wet forest, Puerto Rico. Caribbean Journal of Science, 45: en prensa. [ Links ]
13. Kominoski JS, Pringle CM. 2009. Resource-consumer diversity: testing the effects of leaf litter species diversity on stream macroinvertebrate communities. Freshwater Biology, 54: 1461-1473. [ Links ]
14. Kominoski JS, Pringle CM, Ball BA, Bradford MA, Coleman DC, Hall DB, Hunter MD. 2007. Non-additive effects of leaf litter species diversity on breakdown dynamics in a detritus-based stream. Ecology, 88: 1167-1176. [ Links ]
15. MOBOT (Missouri Botanical Garden). 2009a. Botanical Inventory of the Río Escalerete Reserve. Fecha de consulta 17 de agosto 2009. Disponible en: http://www.mobot.org/mobot/Research/colombia/escalerete [ Links ]
14. MOBOT (Missouri Botanical Garden). 2009b. Guide to the Plants of the Bajo Calima Region. Fecha de consulta 17 de agosto 2009. Disponible en: http://www.mobot. org/mobot/Research/colombia/bajocalima [ Links ]
15. MOBOT (Missouri Botanical Garden). 2009c. Flora tropical. Fecha de consulta 17 de agosto 2009. Disponible en:http://www.tropicos.org [ Links ]
16. Murillo MT, Lozano G. 1989. Hacia la realización de una flórula del Parque Nacional Natural islas de Gorgona y Gorgonilla, Cauca, Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 28: 277-304. [ Links ]
17. Naiman R, Décamps H, McClain ME. 2005. Riparia. Ecology, conservation, and management of stream side communities. Burlington (MA): Elsevier. p. 430. [ Links ]
18. Palta J. 1986. Aspectos ecológicos y de vegetación. En: Prahl H, Alberico M, editores. Isla de Gorgona. Cali (Colombia): Universidad del Valle-Banco de la República. p. 213-221. [ Links ]
19. Rangel O. 1990. Tipos de Vegetación. En: Aguirre J, Rangel O, editores. Biota y Ecosistemas de Gorgona. Bogotá (Colombia): Fondo para la protección del medio ambiente -FEN- Colombia. p. 109-126 [ Links ]
20. Rangel O, Aguirre J, Sánchez R. 1990a. Aspectos estructurales, dinámicos y fisionómicos de la vegetación (una aproximación preliminar). En: Aguirre J, Rangel O, editores. Biota y Ecosistemas de Gorgona. Bogotá (Colombia): Fondo para la protección del medio ambiente -FEN- Colombia. p. 127-141. [ Links ]
21. Rangel O, Fuentes C, Uribe J. 1990b. Aspectos biotipológicos y morfoecológicos de la vegetación. En: Aguirre J, Rangel O, editores. Biota y Ecosistemas de Gorgona. Bogotá (Colombia): Fondo para la protección del medio ambiente -FEN- Colombia. p. 152-169 [ Links ]
22. Rangel O. 2004. La vegetación del Chocó biogeográfico de Colombia y zonas cordilleranas aledañas-Síntesis. En: Rangel O, editor. Colombia, diversidad biótica IV: El Chocó biogeográfico/ Costa Pacífica. Bogotá: Instituto de Ciencias Naturales, Universidad Nacional de Colombia. p. 769-815. [ Links ]
23. Smithsonian Tropical Research Institute's Herbarium (STRI). 2009a. Flora de Barro Colorado. Fecha de consulta 17 de agosto 2009. Disponible en: http://biogeodb.stri.si.edu/bioinformatics/croat/home [ Links ]
24. Smithsonian Tropical Research Institute's Herbarium. 2009b. Flora Bocas del Toro. Fecha de consulta 17 de agosto 2009. Disponible en: http://biogeodb.stri. si.edu/bocas_database [ Links ]
25. Smithsonian Tropical Research Institute's Herbarium. 2009c. Herbario. Fecha de consulta 17 de agosto 2009. Disponible en: http://biogeodb.stri.si.edu/herbarium [ Links ]
26. Swan CM, Palmer MA. 2006. Composition of speciose leaf litter alters stream detritivore growth, feeding activity and leaf breakdown. Oecologia, 147:469-478. [ Links ]
27. Wantzen K, Yule C, Tockner K, Junk W. 2008. Riparian Wetlands of Tropical Streams. En: Dudgeon D, editor. Tropical Stream Ecology. (United States): Elsevier. p. 209-281. [ Links ]
28. Yockteng R, Cavelier J. 1998. Diversidad y mecanismos de dispersión de árboles de la isla Gorgona y de los bosques húmedos tropicales del Pacífico colombo-ecuatoriano. Revista de Biología Tropical 46: 45-53. [ Links ]
29. Zalamea, M, González G. 2008. Leaf fall phenology in a subtropical wet forest in Puerto Rico: from species to community patterns. Biotropica, 40: 295-304. [ Links ]
Recibido: mayo 2009;
Aceptado: noviembre 2009.

