










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActualidades Biológicas
Print version ISSN 0304-3584
Actu Biol vol.33 no.94 Medellín Jan./June 2011
ARTÍCULOS DE INVESTIGACIÓN
RESPUESTA DE TRES GENOTIPOS DE TOMATE AL CULTIVO IN VITRO Y AISLAMIENTO DE PROTOPLASTOS
RESPONSE OF THREE GENOTYPES OF TOMATO TO IN VITRO CULTURE AND ISOLATION OF PROTOPLASTS
Catalina Botero-Giraldo1; Catalina Restrepo-Osorio2; Aura I. Urrea-Trujillo3
1 Instituto de Biología, Universidad de Antioquia. A. A. 1226. Medellín (Antioquia), Colombia. catabuffy@gmail.com.
2 Corporación para Investigaciones Biológicas (CIB), Universidad de Antioquia. A. A. 1226. Medellín (Antioquia), Colombia. cataclooney85@hotmail.com.
3 Docente, Instituto de Biología, Universidad de Antioquia. A. A. 1226. Medellín (Antioquia), Colombia. aurrea@matematicas.udea.edu.co.
Recibido: octubre 2009; aceptado: abril 2010.
Resumen
El objetivo de esta investigación fue evaluar la mejor fuente de material vegetal para el aislamiento de protoplastos, suspensiones celulares o foliolos en tres genotipos de tomate con diferente grado de resistencia a Phytophthora infestans: Solanum habrochaites (503) resistente, S. lycopersicum (507) susceptible y el híbrido parcialmente resistente (508). Luego de lograr el establecimiento in vitro a partir de semilla certificada en los genotipos 503 y 507 y de estacas del genotipo 508, las hojas de las vitroplantas desarrolladas y multiplicadas se usaron para la inducción de callo. Se evaluaron cuatro combinaciones de reguladores de crecimiento y dos condiciones lumínicas por genotipo, encontrándose los porcentajes más altos de formación de callo en oscuridad. Las combinaciones de reguladores con mayor efecto sobre el crecimiento y friabilidad de los callos fueron BAP (1 ppm) + AIA (4 ppm) para el genotipo 503 y BAP (4 ppm) + AIA (1 ppm) para los genotipos 507 y 508. Estas combinaciones de reguladores fueron utilizadas para el establecimiento de las suspensiones celulares y el trazado de la curva de crecimiento. El aislamiento de los protoplastos a partir de suspensiones celulares se realizó en la fase exponencial, evaluando diferentes mezclas enzimáticas y tiempos de incubación; sin embargo, no se logró respuesta para ningún genotipo. A partir del mesófilo, se obtuvo 1,2 x 105 protoplastos/ml con alta viabilidad para el genotipo 503 (celulasa 0,6% y pectinasa 0,1%, 8 h de incubación); 1,9 x 105 protoplastos/ml para el genotipo 507 (celulasa 4% y pectinasa 0,4%, 8 h de incubación); y para el genotipo 508, 1,1 x 105 protoplastos/ml (celulasa 2% y pectinasa 0,2%, 7 h de incubación).
Palabras clave: callo, mesófilo, suspensión celular, Solanum lycopersicum, Solanum habrochaites
Abstract
The goal of this study was to evaluate the best plant material source for protoplast isolation, cell suspensions or leaf mesophyll cells in three tomato genotypes with different levels of resistance to Phytophthora infestans: Solanum habrochaites (503) resistant, S. lycopersicum (507) susceptible, and the hybrid partially resistant (508). After the in vitro establishment from certified seeds of the 503 and 507 genotypes, and cuttings of the 508 genotype, the leaves from the developed and multiplied plantlets were used for callus induction. Four growth regulator combinations per genotype and two light conditions were evaluated, with the highest percentages of callus formation under dark conditions. The combinations of regulators with greatest effect on growth and callus friability were BAP (1 ppm) + IAA (4 ppm) for the 503 genotype and BAP (4 ppm) + IAA (1 ppm) for the 507 and 508 genotypes. These combinations of regulators were used for the establishment of cell suspensions and plotting of the cell growth curve. Protoplast isolation from cell suspensions was conducted in the exponential phase, evaluating different enzymatic mixtures and incubation times; however, no response was obtained for any genotype. From mesophyll, 1.2 x 105 protoplasts/ml with high viability were obtained for the 503 genotype (0.6% cellulose and 0.1% pectinase, 8 h incubation); 1.9 x 105 protoplasts/ml for the 507 genotype (4% cellulose and 0.4% pectinase, 8 h incubation); and 1.1 x 105 protoplasts/ml for the 508 genotype (2% cellulose and 0.2% pectinase, 7 h incubation).
Key words: callus, mesophyll, suspension cells, Solanum lycopersicum, Solanum habrochaites
INTRODUCCIÓN
El tizón tardío, causado por el oomycete Phytophthora infestans Mont. De Bary, es la enfermedad de mayor importancia económica del tomate y de otras solanáceas (Solanum tuberosum L., Solanum phureja, Physalis peruviana L., Solanum quitoense Lamarck) (Carreño et al. 2007, Lourenço-Júnior et al. 2006), esto ha llevado a diversos estudios para su control. Específicamente para el tomate (Solanum lycopersicum L.), debido a la carencia de cultivares comerciales con resistencia efectiva, efectiva, los cultivadores requieren aplicaciones intensivas de fungicidas, provocando problemas para el ambiente, la salud de las personas y generándose además resistencia a los fungicidas por parte de las plantas (Lourenço-Júnior et al. 2006). Los métodos convencionales de mejoramiento presentan como principales limitaciones el largo tiempo para producir una nueva variedad, la dificultad de aprovechar la variabilidad genética de especies o géneros relacionados debido al alto grado de incompatibilidad y la baja eficiencia de la selección (Dixon 2004, Krishna-Ghosh y Guha-Sarkar 2011, Sharma et al. 2002). El cultivo de células y tejidos comprende una amplia gama de técnicas que permiten regenerar plantas completas a partir de tejidos embrionarios, fragmentos de tejido, callos, células aisladas o protoplastos. Esta tecnología conocida como propagación clonal o micropropagación tiene entre otros usos potenciales en los cultivos agrícolas: la producción a gran escala de genotipos heterocigóticos, de genotipos autoincompatibles, un progenitor que presente esterilidad masculina en un programa de mejoramiento genético, la propagación de material fitogenético libre de enfermedades y conservación e intercambio internacional de germoplasma (Poehlman y Allen 2003). Una selección temprana puede ser realizada en plantas in vitro, callos, suspensiones celulares y protoplastos, cuando se presenta respuesta diferencial entre especies y variedades resistentes y susceptibles al patógeno al entrar en contacto con los esporangios, el micelio, el cultivo filtrado del hongo, entre otros (Rodríguez et al. 2007, Urrea et al. 2001, Veitía et al. 2009). La selección in vitro tiene la ventaja de permitir tratar con facilidad gran número de individuos con un determinado agente selectivo, pues utiliza a la célula como unidad básica. Disponer de un método de selección in vitro que permita la reducción de las poblaciones, eliminando la mayor parte de las formas susceptibles en la primera etapa, es de gran importancia, además permite lograr mayor eficiencia en la obtención de una nueva variedad ya que la selección se realiza a nivel celular directa o indirectamente, ya sea en callos o células en suspensión (Urrea 2000).
Es conocido que los protoplastos son un material experimental ampliamente utilizado para investigaciones biológicas. En adición a su rol en la generación de nuevas plantas, los protoplastos proporcionan un único sistema de células individuales para estudios de ultraestructura celular, genética, fisiología e interacciones planta-patógeno (Davey et al. 2005). Protoplastos aislados frescos han sido empleados en estudios relacionados con la síntesis de la pared celular, propiedades de la membrana e infección de virus, mejoramiento de plantas a través de técnicas de manipulación genética, tales como hibridación somática, cibridización, transformación de plantas o transferencia directa de genes y selección in vitro, para estudios bioquímicos y moleculares fluctuando entre investigaciones en las propiedades de crecimiento de células individuales hasta transporte a través de la membrana (Anjum 1998, Bhojwani y Razdan 1996, Hurtado y Merino 1987, Ling et al. 2009, Mühlbach 1980, Rao y Prakash 1995, Szczerbakowa et al. 2000, Tylicki et al. 2002, Zapata et al. 1977).
Muchos protocolos sobre aislamiento y purificación de protoplastos han sido publicados quedando claro que tanto el tipo de explante como el genotipo son determinantes en los resultados obtenidos en cuanto a producción y reproducibilidad. El objetivo de este trabajo fue evaluar la eficiencia en el aislamiento de protoplastos a partir de mesófilo y suspensiones celulares de genotipos de tomate con diferente grado de resistencia a P. infestans, para su posterior utilización en experimentos de selección in vitro.
MATERIALES Y MÉTODOS
Semillas y estacas de las especies de tomate Solanum habrochaites, S. lycopersicum var. cerasiforme y el híbrido producto de las anteriores, denominadas en adelante genotipos 503, 507 y 508, respectivamente, fueron suministradas por el banco de germoplasma del Centro de Investigación ''La Selva'', de Corpoica, Rionegro (Antioquia), Colombia.
Establecimiento y multiplicación in vitro. Para la introducción in vitro de los tres genotipos, las yemas apicales del genotipo 508 y semillas de los genotipos 503 y 507 desinfectadas exitosamente, fueron sembradas en el medio de cultivo basal MS (Murashige y Skoog 1962), suplementado con sacarosa (30 g/l) y agar (7%). El pH se ajustó a 5,7 antes de ser esterilizado a 120 libras de presión y temperatura de 121 ºC durante 15 min.
Las plantas obtenidas fueron transferidas a medio de cultivo fresco de la misma composición descrita anteriormente para ser multiplicadas y contar así con suficientes hojas, explantes requeridos para los experimentos siguientes. El material se mantuvo en una cámara climatizada a una temperatura de 24 ± 2 °C, humedad relativa de 55% y con un fotoperiodo de 16 h/ luz y 8 h/oscuridad.
Inducción y multiplicación de callos. Con el objetivo de obtener callos friables adecuados para el establecimiento de suspensiones celulares, se utilizaron porciones de hojas de las plantas cultivadas in vitro. Las hojas se cortaron en segmentos de 1 cm2 aproximadamente y se colocaron con el envés en contacto con el medio de cultivo. Se evaluaron cuatro combinaciones de reguladores de crecimiento como se describe a continuación y dos condiciones lumínicas (fotoperiodo 16/8 y oscuridad continua). El medio de cultivo basal MS fue adicionado con: 1) BAP (1 ppm) + AIA (0,5 ppm); 2) BAP (4 ppm) + AIA (1 ppm); 3) BAP (1 ppm) + AIA (4 ppm); 4) BAP (1 ppm) + GA3 (1 ppm); y 5) Control (exento de reguladores de crecimiento), de acuerdo con la experiencia en el laboratorio y con lo descrito y con lo reportado por Fuentes et al. (1998) y Brasileiro et al. (1999).
El tamaño de la muestra fue de 5 cajas de Petri por tratamiento, cada una con 4 discos de hoja y para cada genotipo. Los indicadores de respuesta fueron la presencia o no de callo y porcentaje y tipo de callo (friable o compacto), 20 días después de la siembra.
Para la etapa de multiplicación, los callos obtenidos en cada tratamiento se dividieron en porciones iguales de aproximadamente 1 cm y fueron subcultivados en medio fresco de la misma composición. Las condiciones de cultivo fueron las mismas para el crecimiento de las plantas pero en oscuridad continua. Se evaluó el crecimiento del callo a partir del peso ganado (en gramos) 20 días después del subcultivo, así como la friabilidad de los mismos.
Establecimiento de las suspensiones celulares. El establecimiento de las suspensiones celulares para cada genotipo se inició con los callos que presentaron mejor respuesta en la etapa de multiplicación, es decir, con crecimiento activo y de textura friable. Para su disgregación, entre 1,5 y 3,0 g de callo fueron adicionados a 50 ml de medio de cultivo en Erlenmeyers de 250 ml, los cuales se ubicaron en un agitador horizontal a 90 rpm, bajo condiciones de oscuridad y a una temperatura de 24 ± 2 ºC. Después de 8 semanas, se inició el trazado de la curva de crecimiento para determinar el tiempo en alcanzar la etapa exponencial. El experimento se realizó por triplicado. El indicador de crecimiento celular utilizado fue el volumen de células sedimentadas (VCS), para lo cual cada 7 días se tomó una alícuota de 5 ml de cada suspensión, se llevaron a centrifugación a 600 g durante tres minutos para la sedimentación de las células y luego se determinó el volumen, expresándolo en ml de células por ml de cultivo.
Aislamiento de protoplastos. Para determinar la fuente de protoplastos más adecuada por la cantidad y la viabilidad de los mismos, se compararon aquellos provenientes del mesófilo y los obtenidos a partir de las suspensiones celulares, para los 3 genotipos.
Los enzimas utilizadas para el aislamiento fueron: celulasa (SIGMA, a partir de Aspergillus niger), pectinasa (SIGMA, a partir de Rhizopus sp.) y hemicelulasa (SIGMA, a partir de Aspergillus niger).
A partir de mesófilo. Para la obtención de protoplastos a partir de mesófilo se evaluaron los protocolos descritos por Mei-Lie et al. (1987) y Zapata et al. (1977, 1981). El protocolo base usado para el aislamiento de protoplastos se modificó únicamente en la cantidad de gramos en peso fresco de hojas empleada (0,6 g). El número de protoplastos para cada genotipo se estimó a partir de tres conteos bajo un microscopio de luz en un hemocitómetro (cámara de Neubauer Boeco-Germany, profundidad 0,1 mm) y registrado como el número de protoplastos/ml.
Para determinar la viabilidad de los protoplastos se utilizó el método de tinción con el colorante azul de Evans al 0,025%, el cual permite diferenciar las células viables (no teñidas) de las no viables (teñidas).
A partir de suspensiones celulares. Para la obtención de protoplastos a partir de suspensiones celulares se evaluaron los protocolos descritos por Blewett et al. (2000) y Lei et al. (2003).
A partir de los resultados obtenidos en ensayos preliminares, se realizaron modificaciones al protocolo original como se describe a continuación: 0,5 g de células provenientes de suspensiones celulares en crecimiento activo (etapa exponencial en la curva), fueron sometidas a una solución que contenía 0,4 M de manitol y 5 mM de MES, a pH de 5,8. Luego fueron incubadas en una mezcla enzimática que contenía celulasa 2% y pectinasa 0,5% y mantenidas en oscuridad a 25 ºC por 4 h. Después de este tiempo, la mezcla fue diluida con 5 ml de solución de lavado; conteniendo las sales CPW [NaCl (1,0 mM), KCl (0,2 mM) y CaSO4 (0,2 mM)] y KCl 20g/l, seguidamente se llevó a centrifugación a 600 revoluciones por tres minutos. El pellet obtenido se resuspendió por dos veces en 10 ml de medio de lavado y por último se centrifugó dos veces a 600 r/min, la primera durante 3 min y la segunda por 7 min.
Los ensayos se llevaron a cabo con suspensiones celulares de cada genotipo en etapa de multiplicación. El experimento fue repetido al menos 3 veces para cada genotipo. Para determinar la densidad de población y la viabilidad de los protoplastos se utilizaron los métodos descritos para el aislamiento a partir del mesófilo.
Análisis estadístico. Los datos obtenidos en la etapa de inducción y multiplicación de los callos, fueron analizados mediante un ANOVA multifactorial y unifactorial, respectivamente. Para determinar las diferencias significativas entre los tratamientos por el porcentaje de inducción de callo se utilizaron las pruebas de Tamhane, Dunnett T3 o Games-Howell para los análisis no paramétricos y para los datos con distribución normal se emplearon la prueba de Tukey. Las pruebas se consideraron significativas para un error α = 0,05. Los datos se procesaron en el paquete estadístico SPSS versión 13.0.
RESULTADOS
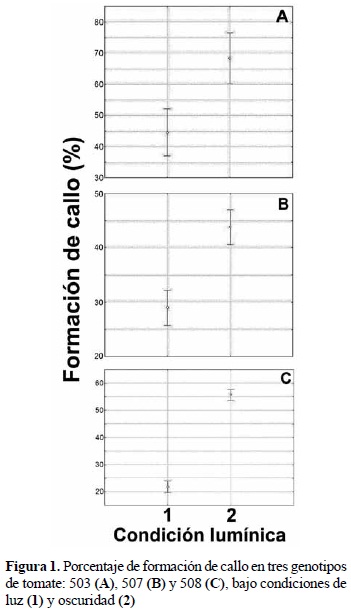
En la etapa de inducción de callo para los tres genotipos se encontraron diferencias significativas, con un valor p ≤ 0,05, entre las condiciones de cultivo (fotoperiodo y oscuridad) en todos los tratamientos (figura 1). Bajo condiciones de fotoperiodo (16/8) los porcentajes de inducción oscilaron entre 37 y 53, 26 y 33, 20 y 24%, en los genotipos 503, 507 y 508, respectivamente. Mientras que en oscuridad los valores oscilaron entre 60 y 77, 41 y 47, 54 y 58% de los respectivos genotipos.
El tratamiento cuatro (BAP 1 ppm + GA3 1 ppm) tuvo un efecto marcado en la regeneración indirecta de brotes para los genotipos 503 y 507 en las dos condiciones lumínicas. El genotipo 508 solamente presentó un encorvamiento de la hoja, similar a la respuesta obtenida en el tratamiento control.
Los callos formados en condiciones de fotoperiodo en los genotipos 503 y 507, respondieron formando brotes y yemas adventicias y en algunos casos raíces (regeneración indirecta), principalmente en los tratamientos con reguladores. En el genotipo 508, no se formaron brotes o yemas en ningún tratamiento, no obstante mostró muy buena capacidad de regeneración de raíces en el tratamiento tres (BAP 1 ppm + AIA 4 ppm).
Bajo condiciones de oscuridad continua, en el genotipo 503 hubo proliferación de callo sin regeneración de órganos. Los genotipos 507 y 508, solo en el tratamiento uno (BAP 1 ppm + AIA 0,5 ppm) presentaron regeneración de raíces en baja cantidad, mientras que en los tratamientos dos (BAP 4 ppm + AIA 1 ppm) y tres (BAP 1 ppm + AIA 4 ppm) se presentó abundante formación de callo.
El color de los callos también se vio afectado por las condiciones lumínicas, en fotoperiodo se tornaron verde y café oscuro en los genotipos 503 y 508, respectivamente. Mientras que en oscuridad para estos dos genotipos el callo fue café claro. Para el genotipo 507 tanto en condición de fotoperiodo como en oscuridad, el callo se tornó de color café claro con porciones blanquecinas.
Efecto de diferentes combinaciones de reguladores de crecimiento y condiciones lumínicas sobre la formación de callo en los genotipos de tomate 503, 507 y 508. Con el objetivo de lograr la formación de suficiente callo friable, se evaluaron diferentes combinaciones y concentraciones de reguladores de crecimiento, utilizando como medio basal el MS. Para el genotipo 503 en condiciones de fotoperiodo (16/8), en el tratamiento dos, se alcanzó el mayor porcentaje de inducción (X: 86%) con diferencia significativa respecto a los demás tratamientos (tabla 1); sin embargo, presentaron regeneración de brotes, lo cual no era el objetivo. Por el contrario, en condiciones de oscuridad, los mayores porcentajes se alcanzaron en los tratamientos uno, dos y tres, sin diferencias significativas entre ellos, no obstante teniendo en cuenta que el tratamiento tres, BAP (1 ppm) + AIA (4 ppm), además de lograr mayor porcentaje de callo, las características del mismo tales como la friabilidad y coloración fueron las adecuadas, se seleccionó como la mejor combinación de reguladores para la inducción de callo.
Para el genotipo 507 en condiciones de fotoperiodo, los tratamientos dos y tres alcanzaron los mayores porcentajes de inducción (X: 38, X: 42, respectivamente) con diferencia significativa respecto a los otros dos tratamientos (tabla 1). Los callos formados en los tratamientos uno y tres a su vez dieron origen a raíces. Por el contrario en condiciones de oscuridad, el mayor porcentaje de callo se alcanzó con el tratamiento dos (BAP 4 ppm + AIA 1 ppm) con diferencia estadística altamente significativa con respecto a los demás tratamientos y por lo tanto fue seleccionado para etapas posteriores.
Para el genotipo 508 en condiciones de fotoperiodo, con el tratamiento dos se alcanzó el mayor porcentaje de inducción de callo (X: 45%) con diferencia significativa respecto a los demás tratamientos (tabla 1). En condiciones de oscuridad los mayores porcentajes de callo se alcanzaron en los tratamientos dos y tres, sin diferencias significativas entre ellos, sin embargo el tratamiento dos, BAP (4 ppm) + AIA (1 ppm), alcanzó mayor porcentaje de callo (95%) y la friabilidad y coloración permitieron seleccionarlo como el más adecuado.
Multiplicación de callos. De acuerdo con los resultados obtenidos en la etapa de inducción, para la multiplicación de los callos se seleccionó la oscuridad continua y se evaluaron los mismos tratamientos de la etapa de inducción, excepto el tratamiento cuatro (BAP 1 ppm + GA3 1 ppm) debido a que este indujo regeneración de brotes en los genotipos 503 y 507, y el genotipo 508 no presentó respuesta.
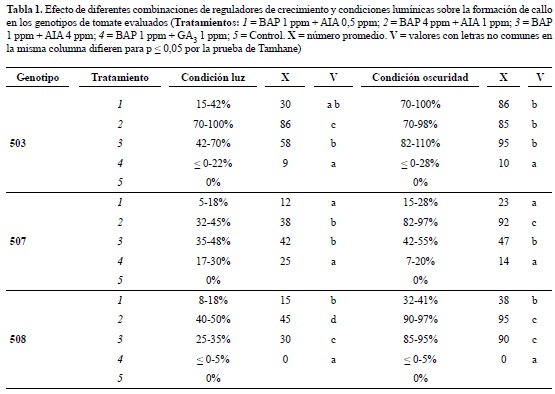
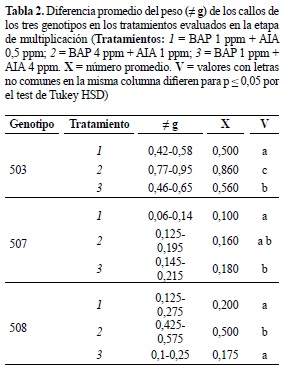
Para los genotipos 503 y 508, en el análisis de comparaciones múltiples realizado, se encontró que el mayor valor de peso fresco (0,86 y 0,5 g de callo, respectivamente) se alcanzó en el tratamiento dos, pero al evaluar las características del callo en el genotipo 503 se comprobó su consistencia compacta y aspecto nodular, por el contrario en el tratamiento tres luego de la segunda multiplicación, el callo presentó mayor friabilidad y por lo tanto se seleccionó para posteriores ensayos (figura 2A; tabla 2). Para el genotipo 508 el tratamiento dos favoreció tanto el incremento en peso como las características de interés (figura 2C; tabla 2). En el genotipo 507 no se encontraron diferencias entre los tres tratamientos, sin embargo dada la mayor friabilidad en el tratamiento dos, se escogió este para la etapa de multiplicación (figura 2B; tabla 2).
Establecimiento de suspensiones celulares. Con los callos que en la etapa de multiplicación presentaron mejor respuesta en cuanto a friabilidad y crecimiento se llevó a cabo la disgregación celular y el establecimiento de las suspensiones celulares. El volumen de células sedimentadas (VCS) registrado a partir de 5 ml de suspensión cada 7 días para los tres genotipos, se presenta en la tabla 3.
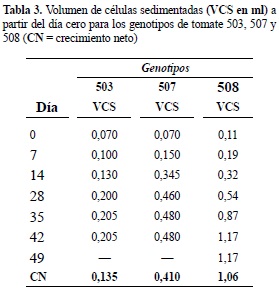
Para los 3 genotipos se presentó crecimiento exponencial desde el día cero, con la continua división celular. Para el genotipo 503 y 507, al día 28 comenzó la fase estacionaria, mientras que para el genotipo 508, inició el día 42 (figura 3). El mayor crecimiento neto se registró para el genotipo 508 (1,06 ml), seguido del genotipo 507 (0,41 ml) y 503 con el menor crecimiento neto (0,135 ml) (tabla 3).
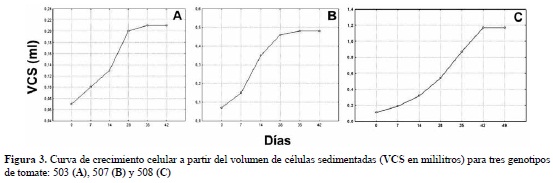
Teniendo en cuenta las curvas de crecimiento celular obtenidas, se decidió realizar los experimentos de aislamiento de protoplastos entre el día 14 y el 28 para 503, entre el día 7 y el 28 para 507 y para 508 entre los días 14 y 42, correspondientes a las fases exponenciales de cada genotipo. Esta etapa, según lo descrito por Pierik (1990), es la recomendada para obtener mayor cantidad de protoplastos viables.
Aislamiento de protoplastos. A partir de mesófilo. Para los tres genotipos el tiempo de incubación y la concentración de enzimas evaluados inicialmente no favorecieron el aislamiento de protoplastos viables. Después de 18 h de incubación, se encontró alto porcentaje de células dañadas, al parecer las enzimas degradaron tanto las proteínas de la pared celular como de la membrana celular. A partir de lo anterior, se procedió a realizar modificaciones al protocolo, principalmente en la concentración de las enzimas y tiempo de exposición del tejido a la mezcla enzimática, con el propósito de lograr mayor cantidad y viabilidad por encima del 80%.
Para el genotipo 503, la mezcla enzimática y tiempo de incubación más apropiadas para el aislamiento de protoplastos fue celulasa 0,6% y pectinasa 0,1% durante 8 h, obteniéndose viabilidad del 81,5% y concentración de protoplastos de 1,2 x 105 (figura 4A, tabla 4).
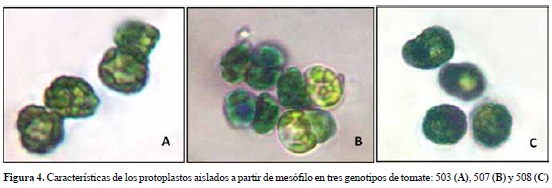
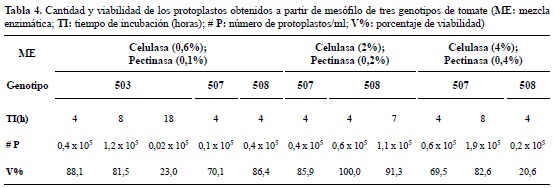
En el genotipo 507, con la mezcla enzimática celulasa 4% y pectinasa 0,4%, y un tiempo de incubación de 8 h, se alcanzó 1,9 x 105 protoplastos/ ml con una viabilidad del 82,6% (figura 4B; tabla 4).
Para el genotipo 508, de las 3 mezclas enzimáticas evaluadas, la que consiguió un mayor número de protoplastos fue celulasa 2% y pectinasa 0,2% con 7 horas de incubación, obteniendo 1,1 x 105 protoplastos/ml con una viabilidad del 91,3% (figura 4C; tabla 4).
El número de protoplastos viables alcanzado en cada uno de los genotipos coincide con la cantidad recomendada para el cultivo en trabajos de mejoramiento a través de la fusión de protoplastos.
A partir de suspensiones celulares. Se tomaron suspensiones celulares en fase exponencial para los ensayos de aislamiento de protoplastos. En el primer ensayo realizado usando celulasa 2% y pectinasa 0,5% y un tiempo de incubación de 4 h no se obtuvieron protoplastos en ningún genotipo, observándose al microscopio plasmólisis incipiente de la membrana celular. A este protocolo inicial se le hicieron varias modificaciones en cuanto a la concentración de enzimas y tiempo de incubación, sin embargo no se logró el aislamiento de protoplastos en ninguna de estas, para los genotipos en estudio.
Con la mezcla enzimática de celulasa 3% y pectinasa 1,5% a las 4 h de incubación se alcanzó un porcentaje de viabilidad del 60%, mientras que con 8 h y la misma mezcla enzimática se redujo a 52,5%. Cuando se evaluó la mezcla enzimática de celulasa 3%, pectinasa 1,5% y hemicelulasa 1% con 12 horas de incubación el porcentaje de viabilidad fue de solo el 35,2%.
De las dos fuentes de protoplastos evaluadas, el mesófilo fue la mejor opción para los tres genotipos, el establecimiento de las suspensiones celulares además de tomar más tiempo, se dificultó el aislamiento de protoplastos ya que las células tienden a agruparse formando agregados que son difíciles de separar aun con las enzimas.
DISCUSIÓN
Inducción y multiplicación de callos. Se ha documentado que en miembros de la familia Solanaceae, las auxinas solas o en combinación con una citoquinina en el medio de cultivo favorecen la formación de callo. Los resultados obtenidos en este trabajo coinciden con lo descrito por Harish et al. (2010) y Jatoi et al. (2001), ya que en condiciones de oscuridad continua en los diferentes tratamientos con la combinación BAP y AIA, se alcanzaron los mayores porcentajes de formación de callo para los tres genotipos de tomate.
Teniendo en cuenta que la formación de los callos tenía como finalidad el establecimiento de suspensiones celulares, tanto el crecimiento como la friabilidad fueron los aspectos más importantes que se deben tener en cuenta. En este estudio el genotipo, la condición lumínica y la concentración de reguladores fueron determinantes en la friabilidad. Se ha reportado que son relevantes además para esta característica el número de subcultivos y la consistencia del medio (Carron et al. 1998, Cetz 2005).
Los tratamientos que generaron callos con mayor friabilidad fueron las combinaciones de BAP a 4 ppm + AIA a 1 ppm y BAP a 1 ppm + AIA a 4 ppm en condiciones de oscuridad.
Es relevante anotar que las concentraciones de estos dos reguladores tuvieron un efecto sobre la formación de callos con mayor friabilidad dependiendo del genotipo, así 4 ppm de BAP y 1 ppm de AIA generó la mejor respuesta para 507 y 508 (híbrido parcialmente resistente a P. infestans), por el contrario con 4 ppm de AIA y 1 ppm de BAP, solo el genotipo 503 respondió favorablemente.
Se ha reportado que el éxito para la regeneración in vitro en tomate varía con el medio nutritivo, concentraciones y combinaciones de reguladores de crecimiento, luz y regímenes de temperatura en el cuarto de incubación, genotipo y factores relacionados al explante (Bhatia et al. 2004, Devi et al. 2008). Los resultados obtenidos en este trabajo mostraron que la regeneración indirecta de brotes y yemas adventicias obtenidos en los genotipos 503 y 507 con el tratamiento cuatro (BAP 1 ppm + GA3 1 ppm) en condiciones de fotoperiodo y oscuridad se puede atribuir tanto a la combinación de reguladores como al genotipo directamente. De igual manera, para estos genotipos se logró la regeneración indirecta de pequeños brotes y raíces en los demás tratamientos pero solo bajo condiciones de fotoperiodo. Aunque la combinación de reguladores BAP + AIA ha sido reportada por varios autores en protocolos de regeneración de plantas de tomate (Álvarez et al. 1999, Fuentes et al. 1998, Mohamed et al. 2010, Rzepka-Plevneš et al. 2006, Soniya et al. 2001), en este trabajo se logró la formación y multiplicación de callo principalmente cuando los cultivos se mantuvieron en oscuridad.
Establecimiento de suspensiones celulares. Es conocido que una de las fuentes para obtener protoplastos viables son las suspensiones celulares. El éxito del establecimiento de estas a partir de fragmentos de callo depende en gran medida de la friabilidad del tejido calloso (Chawla 2002, Chin 2008, Godoy-Hernández y Vásquez-Flota 2006).
La diferencia del crecimiento celular neto, como volumen de VCS, encontrado para los tres genotipos, se puede explicar primero por la influencia que obviamente tiene el genotipo y segundo por la cantidad de inóculo inicial, la cual no fue igual para todos. El genotipo 508 demoró 1 semana más respecto a los otros genotipos para alcanzar la fase estacionaria, debido probablemente a que la cantidad de inóculo inicial fue la menor (1,85 g/50 ml). El tamaño del inóculo inicial determina el periodo lag de la curva de crecimiento celular, por lo tanto, con una alta densidad del inóculo inicial se reduce la longitud de la fase lag (Chawla 2002). En ninguno de los genotipos evaluados se obtuvo esta fase, lo cual sugiere que el inóculo inicial fue suficiente para que las células comenzaran la fase exponencial desde el día cero con una continua división celular y sin alcanzar la saturación tempranamente.
Aunque el medio seleccionado para el establecimiento de las suspensiones fue el de mejor respuesta en la etapa de multiplicación de los callos, no se puede asegurar si la consistencia líquida del medio, los requerimientos de sales minerales, hormonas y otros nutrientes o por el contrario suprimir algunos de estos puedan contribuir al mejor crecimiento celular en estos genotipos.
Al inicio del establecimiento de las suspensiones celulares, éstas fueron mantenidas en condiciones de luz, sin embargo, debido al pobre crecimiento en estas condiciones, fueron transferidas a oscuridad continua, logrando mayor crecimiento en esta condición, lo cual coincide con lo descrito por Latif et al. (1993), quienes encontraron que bajo condiciones de luz, la división celular temprana fue inhibida.
Por otro lado, Brownleader y Dey (1997) y Encina et al. (2001), recomiendan de manera general que los callos seleccionados para iniciar el establecimiento de las suspensiones celulares permanezcan en la oscuridad o a muy bajos niveles de iluminación, así como también las suspensiones celulares.
Aislamiento de protoplastos. A partir de mesófilo. Para el aislamiento de protoplastos a partir del mesófilo, las plantas in vitro son preferidas a las de maceta, debido a que sus hojas poseen cutícula y pared más delgada, lo cual facilita el proceso de digestión de la pared celular (Borgato et al. 2007). Esto se corroboró en los ensayos preliminares realizados con hojas de plantas mantenidas en la estación biológica (Universidad de Antioquia, Medellín, Colombia), con las cuales fueron muy pocos los protoplastos obtenidos.
Diferentes autores afirman que el aislamiento y obtención de protoplastos es específico para cada especie (Divakaran et al. 2008, Rizkalla et al. 2007, Schum et al. 2001, Szabados 1991, Szczerbakowa et al. 2000); en el presente trabajo, igualmente cada genotipo respondió de manera diferente a las concentraciones enzimáticas y al tiempo de incubación del tejido foliar. Mientras que para el genotipo resistente a P. infestans, 503, se requirió las menores concentraciones enzimáticas de celulasa y pectinasa, 0,6 y 0,1%, respectivamente, para el genotipo susceptible a P. infestans, 507, fue necesaria mayor concentración enzimática, celulasa 4% y pectinasa 0,4%.
El aislamiento de protoplastos a partir del mesófilo fue eficiente de acuerdo con la literatura consultada, ya que Kulawiec et al. (2003) obtuvieron para L. esculentum una eficiencia entre 2,5 a 5 x 105 por un gramo de hoja, no obstante, Dorion et al. (1999) registraron para Lycopersicon cheesmanii el aislamiento altamente eficiente de protoplastos con una producción media de 3 x 107 por gramo de hoja fresca.
Davey et al. (2005) afirman que, generalmente, la densidad óptima de protoplastos es de 5 x 102-1,0 x 106 protoplastos/ml para un cultivo exitoso, dependiendo de la especie/cultivar.
A partir de suspensiones celulares. Las células predominantes cuando las suspensiones celulares se encuentran en fase exponencial son las meristemáticas, éstas al ser células que están en continua división, las paredes son más delgadas siendo teóricamente más fácil el aislamiento de los protoplastos, sin embargo en el presente estudio aunque en los tres genotipos el aislamiento se evaluó exclusivamente durante la fase exponencial, no se logró la liberación de protoplastos. Contrario a lo reportado por Lei et al. (2003), en el presente trabajo la mezcla enzimática aplicada inicialmente, celulasa (2%) y pectinasa (0,5%) no resultó efectiva para el aislamiento de protoplastos a partir de suspensiones celulares en los tres clones de tomate, no obstante la viabilidad celular se mantuvo por encima del 80%. Con el incremento de la concentración de ambas enzimas y la exposición a diferentes tiempos de incubación, además de la adición de hemicelulasa en uno de los tratamientos, tampoco se obtuvieron resultados favorables al aislamiento y la viabilidad de las células disminuyó a medida que aumentaba el tiempo de exposición a la mezcla enzimática.
Aunque se puede lograr mayor producción de protoplastos usando altas concentraciones enzimáticas, en muchos casos la viabilidad se ve afectada ya que se puede producir lisis de estos tan pronto son liberados. Aunque inicialmente con 4 h de incubación y con baja concentración de enzimas no se logró el aislamiento de protoplastos, la viabilidad de las células se mantuvo alta, no obstante con el mismo tiempo de incubación y mayor concentración enzimática tampoco se logró el aislamiento, sin embargo se presentó alto porcentaje de daño celular.
La producción de compuestos que inhiben la digestión de la pared y subsecuentemente la liberación de protoplastos a partir del explante, ha sido reportada por Rao y Prakash (1995). Rethmeier et al. (1991), relacionaron además la baja viabilidad y frecuencia de regeneración a partir de protoplastos con los altos niveles de etileno inducidos por las enzimas durante el aislamiento. De otro lado, Kennedy y De Filippis (2004) identificaron entre algunas estrategias para disminuir el efecto degradativo deletéreo durante el aislamiento, un control estricto del pH, evaluar el número de enzimas y la adición de antioxidantes e inhibidores de proteasas. Este último aspecto fue evaluado por Santos et al. (2010), quienes encontraron acumulación de proteínas extracelulares en el medio de cultivo debido a cambios estructurales o modificaciones bioquímicas a nivel celular y podría estar relacionada con la lisis celular. Teniendo en cuenta que el daño celular incrementó con el tiempo de incubación, este es otro factor crítico a considerar, como lo informan Davey et al. (2005), Horváth (2009), Ishikawa et al. (2010), Ling et al. (2010) y Pongchawee et al. (2006).
AGRADECIMIENTOS
Se desea agradecer al Laboratorio de Fisiología y Cultivo de Tejidos Vegetales, a la Universidad de Antioquia por su apoyo financiero y a Corpoica por el suministro de material vegetal.
REFERENCIAS
1. Álvarez M, Rodríguez J, Plana D, Santana N. 1999. Radiosensibilidad a rayos gamma 60Co en callos de tomate (Lycopersicon esculentum, Mill.) variedad AMALIA. Cultivos Tropicales, 20 (4): 35-39. [ Links ]
2. Anjum MA. 1998. Effect of protoplast source and media on growth regenerability of protoplast-derives calluses of Solanum tuberosum L. Acta Physiologiae Plantarum, 20 (2): 129-133. [ Links ]
3. Bhatia P, Ashwath N, Senaratna T. 2004. Effect of cytokinins on organogenesis and callus induction in cotyledonary explants of tomato (Lycopersicon esculentum Mill.). En: Islam AS, editors. In vitro culture, transformation and molecular markers for crop improvement. Enfield (U. S. A.): Science Publishers. p. 17-24. [ Links ]
4. Bhojwani SS, Razdan MK. 1996. Plant Tissue Culture: Theory and practice. A revised edition. Amsterdam: Elsevier Science. p. 767. [ Links ]
5. Blewett J, Burrows K, Thomas C. 2000. A micromanipulation method to measure the mechanical properties of single tomato suspension cells. Biotechnology Letters, 22: 1877-1883. [ Links ]
6. Borgato L, Pisani F, Furini A. 2007. Plant regeneration from leaf protoplasts of Solanum virginianum L. (Solanaceae). Plant Cell, Tissue and Organ Culture, 88: 247-252. [ Links ]
7. Brasileiro ACR, Willadino L, Carvalheira GG, Guerra M. 1999. Callus induction and plant regeneration of tomato (Lycopersicon esculentum CV. IPA 5) via anther culture. Ciência Rural (Santa Maria), 29 (4): 619-623. [ Links ]
8. Brownleader MD, Dey PM. 1997. Plant cell biotechnology. En: Dey PM, Harborne JB, editors. Plant Biochemistry. San Diego (California, U. S. A.): Academic Press. p. 517-529. [ Links ]
9. Carreño N, Vargas A, Bernal AJ, Restrepo S. 2007. Problemas fitopatológicos en especies de la familia Solanaceae causados por los géneros Phytophtora, Alternaria y Ralstonia en Colombia. Una revisión. Agronomía Colombiana, 25 (2): 320-329. [ Links ]
10. Carron MP, Lardet L, Dea BG. 1998. Micropropagation de l'Hevea par embryogenèse somatique. Plantations Recherche Développement, 5 (3): 187-194. [ Links ]
11. Cetz J. 2005. Micropropagación de chile dulce (Capsicum annuum L. var. Najera.) y chile habanero (Capsicum chinense Jacq.) con miras al mejoramiento genético del cultivo. [Tesis de maestría]. [Turrialba (Costa Rica)]: Centro Agronómico Tropical de Investigación y Enseñanza (CATIE). Programa de educación para el desarrollo y la conservación. Escuela de Posgrado. p. 73. [ Links ]
12. Chawla HS. 2002. Introduction to plant biotechnology. Second edition. Enfield, New Hampshire (U. S. A.): Science Publishers. p. 538. [ Links ]
13. Chin Ch-K. 2008. Plant cell culture as source of valuable chemicals. En: Havkin-Frenkel D, Belanger FC, editors. Biotechnology in flavor production. Chennai (India): Blackwell Publishing Ltd. p. 104-114. [ Links ]
14. Davey MR, Anthony P, Power JB, Lowe K.C. 2005. Plant protoplast technology: Current status. Review. Acta Physiologiae Plantarum, 27 (1): 117-129. [ Links ]
15. Devi R, Dhaliwal MS, Kaur A, Gosal SS. 2008. Effect of growth regulators on in vitro morphogenic response of tomato. Indian Journal of Biotechnology, 7: 526-530. [ Links ]
16. Divakaran M, Pillai GS, Nirmal-Babu K, Peter KV. 2008. Isolation and fusion of protoplasts in Vanilla species. Current Science, 94 (1): 115-120. [ Links ]
17. Dixon A [Internet]. 2004. Conventional Breeding: lessons learned. International Center for Tropical Agriculture. Fecha de acceso: 15 de diciembre de 2010. Disponible en: http://webapp.ciat.cgiar.org/webciat/biotechnology/cbn/sixth_international_meeting/pdf_presentations/dia%2012/Alfred_Dixon2.pdf. [ Links ]
18. Dorion N, Wies N, Burteaux A, Bigot C. 1999. Protoplast and leaf explants culture of Lycopersicon cheesmanii and salt tolerance of protoplast-derived calli. Plant Cell, Tissue and Organ Culture, 56: 9-16. [ Links ]
19. Encina CL, Constantin M, Botella J. 2001. An easy and reliable method for establishment and maintenance of leaf and root cell cultures of Arabidopsis thaliana. Plant Molecular Biology Reporter, 19: 245-248. [ Links ]
20. Fuentes A, Soto N, Alfonso D, Oramas P. 1998. Estudio de las condiciones de cultivo para la regeneración de plantas de tomate (Lycopersicon esculentum MILL), a partir de cotiledones y hojas de la variedad Campbell 28. Biotecnología aplicada [Internet], 15 (4): 242-245. Fecha de acceso: 12 de noviembre de 2010. Disponible en: http://www.bioline.org.br/request?ba98039. [ Links ]
21. Godoy-Hernández G, Vázquez-Flota FA. 2006. Growth measurements. Estimation of cell division and cell expansion. En: Loyola-Vargas VM, Vásquez-Flota F, editors. Plant cell culture protocols. 2nd edition. New Jersey (U. S. A.): Humana Press. p. 51-58. [ Links ]
22. Harish MC, Rajeevkumar S, Sathishkumar R. 2010. Efficient in vitro callus induction and regeneration of different tomato cultivars of India. Asian Journal of Biotechnology, 2 (3): 178-184. [ Links ]
23. Horváth E. 2009. Protoplast isolation from Solanum lycopersicum L. leaf tissue and their response to shortterm NaCl treatment. Acta Biologica Szegediensis, 53 (2): 83-86. [ Links ]
24. Hurtado MD, Merino MM. 1987. Cultivo de tejidos vegetales. México, D. F. (México): Editorial Trillas. p. 232. [ Links ]
25. Ishikawa FH, Barcelos Q de L, Souza EA de, Dias ES. 2010. Factors affecting the production and regeneration of protoplasts from Colletotrichum lindemuthianum. Ciência Agrotécnica (Lavras), 34 (1): 74-79. [ Links ]
26. Jatoi SA, Sajid GM, Sappal HU, Baloch MS, Quraishi A, Anwar R [Internet]. 2001. Differential in vitro response of tomato hybrids against a multitude of hormonal regimes. OnLine Journal of Biological Sciences [Internet], 1 (12): 1141-1144. Fecha de acceso: 28 de octubre de 2010. Disponible en: http://scialert.net/qredirect.php?doi=jbs.2001.1141.1144linkid=pdf. [ Links ]
27. Kennedy BF, De Filippis LF. 2004. Tissue degradation and enzymatic activity observed during protoplast isolation in two ornamental Grevillea species. In Vitro Cellular and Developmental Biology-Plant, 40 (1): 119-125. [ Links ]
28. Krishna-Ghosh S, Guha-Sarkar Ch [Internet]. 2011. Monsanto company. Fecha de acceso: 18 de diciembre de 2010. Disponible en: http://www.monsanto.co.uk/news/ukshowlib.phtml?uid=4930. [ Links ]
29. Kulawiec M, Tagashira N, Plader W, Bartoszewski M, Kuc D, Sniezko R, Malepszy S. 2003. Chromosome number variation in somatic hybrids between transgenic tomato (Lycopersicon esculentum) and Solanum lycopersicoides. Journal of Applied Genetics, 44 (4): 431-447. [ Links ]
30. Latif M, Mumtaz N, Davey MR, Power JB. 1993. Plant regeneration from protoplasts isolated from cell suspension cultures of the wild tomato, Lycopersicon chilense Dun. Plant Cell, Tissue and Organ Culture, 32: 311-317. [ Links ]
31. Lei X-Y, Zhu R-Y, Zhang G-Y, Dai Y-R. 2003. Possible involvement of the mitochondrial alternative pathway in ethylene-induced apoptosis in tomato protoplasts. Plant Growth Regulation, 41: 111-116. [ Links ]
32. Ling APK, Ling CP, Siang C, Hussein S. 2009. Establishment of protoplast isolation protocols of Orthosiphon staminues. American-Eurasian Journal of Sustainable Agriculture, 3 (3): 587-596. [ Links ]
33. Ling APK, Phua GAT, Tee CS, Hussein S. 2010. Optimization of protoplast isolation protocols from callus of Eurycoma longifolia. Journal of Medicinal Plants Research, 4 (17): 1778-1785. [ Links ]
34. Lourenço-Júnior V, Maffia LA, da Silva-Romeiro R, Mizubuti ESG. 2006. Biocontrol of tomato late blight with the combination of epiphytic antagonists and rhizobacteria. Biological Control, 38: 331-340. [ Links ]
35. Mei-Lie MC Tan, Boerrigter HS, Kool AdJ. 1987. A rapid procedure for plant regeneration from protoplasts isolated from suspension cultures and leaf mesophyll cells of wild Solanum species and Lycopersicon pennellii. Plant Science, 49: 63-72. [ Links ]
36. Mohamed A-a N, Ismail MR, Rahman MH. 2010. In vitro response from cotyledon and hypocotyls explants in tomato by inducing 6-benzylaminopurine. African Journal of Biotechnology, 9 (30): 4802-4807. [ Links ]
37. Mühlbach HP. 1980. Different regeneration potentials of mesophyll protoplasts from cultivated and a wild species of tomato. Planta, 148: 89-96. [ Links ]
38. Murashige T, Skoog F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Plant Physiology, 15: 473-497. [ Links ]
39. Pierik RLM. 1990. Cultivo in vitro de las plantas superiores. Madrid (España): Ediciones Mundi-Prensa. p. 326. [ Links ]
40. Poehlman JM, Allen D. 2003. Aplicación de la biología molecular al fitomejoramiento. En: Poehlman JM, Allen D. Mejoramiento genético de las cosechas. 2.a ed. México, D. F. (México): Editorial Limusa, S. A. p. 145-168. [ Links ]
41. Pongchawee K, Na-Nakorn U, Lamseejan S, Poompuang S, Phansiri S. 2006. Factors affecting the protoplast isolation and culture of Anubias nana Engler. International Journal of Botany, 2 (2): 193-200. [ Links ]
42. Rao KS, Prakash AH. 1995. A simple method for the isolation of plant protoplasts. Journal of Biosciences, 20 (5): 645-655. [ Links ]
43. Rethmeier NOM, Jansen CE, Snel EAM, Nijkamp HJJ, Hille J. 1991. Improvement of regeneration of Lycopersicon pennellii protoplasts by decreasing ethylene production. Plant Cell Reports, 9: 539-543. [ Links ]
44. Rizkalla AA, Badr-Elden AM, Nower AA. 2007. Protoplast isolation, salt stress and callus formation of two date palm genotypes. Journal of Applied Sciences Research, 3 (10): 1186-1194. [ Links ]
45. Rodríguez NV, Kowalski B, Rodríguez LG, Caraballoso IB, Suárez MA, Pérez PO, Quintana CR, González N, Ramos RQ. 2007. In vitro and ex vitro selection of potato plantlets for resistance to early blight. Journal of Phytopathology, 155: 582-586. [ Links ]
46. Rzepka-Plevne D, Kulpa D, Grabiec M, Kowalczys K, Kurek J. 2006. The effect of growth regulators and culture conditions on the callus induction in tomato Lycopersicon sp. Acta Scientiarum Polonorum, Hortorum Cultus, 5 (2): 23-34. [ Links ]
47. Santos ALW, Silveira V, Steiner N, Maraschin M, Guerra MP. 2010. Biochemical and morphological changes during the growth kinetics of Araucaria angustifolia suspension cultures. Brazilian Archives of Biology and Technology, 53 (3): 497-504. [ Links ]
48. Schum A, Hofmann K, Ghalib N, Tawflk A. 2001. Factors affecting protoplast isolation and plant regeneration in Rosa spp. Gartenbauwissenschaft, 66 (3): 115-122. [ Links ]
48. Sharma HC, Crouch JH, Sharma KK, Seetharama N, Hash CT. 2002. Applications of biotechnology for crop improvement: prospects and constraints. Review. Plant Science, 163: 381-395. [ Links ]
50. Soniya EV, Banerjee NS, Das MR. 2001. Genetic analysis of somaclonal variation among callus-derived plants of tomato. Current Science, 80 (9): 1213-1215. [ Links ]
51. Szabados L. 1991. Protoplastos: aislamiento, cultivo y regeneración de plantas. En: Roca WM, Mroginski LA, editores. Cultivo de tejidos en la agricultura: Fundamentos y aplicaciones. Cali (Colombia): Publicación CIAT, Centro Internacional de Agricultura Tropical. p. 240-270. [ Links ]
52. Szczerbakowa A, Borkowska M, Wielgat B. 2000. Plant regeneration from the protoplasts of Solanum tuberosum, S. nigrum and S. bulbocastanum. Acta Physiologiae Plantarum, 22 (1): 3-10. [ Links ]
53. Tylicki A, Burza W, Malepszy S, Kulawiec M, Kuras. 2002. Structural and ultrastructural analysis of Solanum lycopersicoides protoplasts during diploid plant regeneration. Annals of Botany, 90: 269-278. [ Links ]
54. Urrea AI. 2000. Efecto de Phytophthora infestans (Mont.) de Bary y sus metabolitos en la selección de somaclones de papa (Solanum tuberosum L.) var. Diacol Capiro obtenidos por mutagénesis in vitro y variación somaclonal. [Tesis de doctorado]. [Santa Clara (Cuba)]: Universidad Central Marta Abreu de las Villas, Facultad de Ciencias Agropecuarias, Instituto de Biotecnología de las Plantas. p. 90. [ Links ]
55. Urrea AI, Veitía N, Bermúdez I. 2001. Selección in vitro de callos de papa (Solanum tuberosum) var. Diacol Capiro empleando el filtrado crudo de Phytophtora infestans (MONT) DE BARY. Actualidades Biológicas, 23 (74): 5-13. [ Links ]
56. Veitía N, García LR, Bermúdez-Caraballoso I, Acosta-Suárez M, Leiva-Mora M, Alvarado-Capó Y, Torres D, Padrón Y. 2009. Protocolo para la selección in vitro de plantas de papa resistentes al filtrado de cultivo de Alternaria solani Sor. Biotecnología Vegetal, 9 (1): 27-32. [ Links ]
57. Zapata FJ, Evans PK, Power JB, Cocking EC. 1977. The effect of temperature on the division of leaf protoplasts of Lycopersicon esculentum and Lycopersicon peruvianum. Plant Science Letters, 8: 119-124. [ Links ]
58. Zapata FJ, Sink KC, Cocking EC. 1981. Callus formation from leaf mesophyll protoplasts of three Lycopersicon species: L. esculentum C. V. Walter, L. pimpinillifolium and L. hirsutum, F. glabratum. Plant Science Letters, 23 (1): 41-46. [ Links ]

