










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActualidades Biológicas
Print version ISSN 0304-3584
Actu Biol vol.35 no.99 Medellín July/Dec. 2013
ARTÍCULOS DE INVESTIGACIÓN
DIVERSIDAD ARBÓREA Y PRIORIDADES DE CONSERVACIÓN DE LOS BOSQUES SECOS TROPICALES DEL SUR DEL DEPARTAMENTO DEL TOLIMA EN EL VALLE DEL RÍO MAGDALENA, COLOMBIA
TREE'S DIVERSITY AND CONSERVATION PRIORITIES OF TROPICAL DRY FORESTS OF THE SOUTHERN TOLIMA ON THE MAGDALENA RIVER VALLEY, COLOMBIA
Fernando Fernández-Méndez1,3; Jonny F. Bernate-Peña2; Omar Melo1,4
1 Grupo de Investigación en Biodiversidad y Dinámica de Ecosistemas, Tropicales, Facultad de Ingeniería Forestal, Universidad del Tolima, Bloque 26-12 Laboratorio de Dendrología. A. A. 546 Ibagué, Colombia.
2 Faculty of Forestry and Wood Sciences, Czech University of Life Sciences Prague. Prague, Czech Republic. jonferber@yahoo.com.
Correos electrónicos: 3 fmendez@ut.edu.co; 4 omelo@ut.edu.co.
Recibido: octubre 2012; aceptado: septiembre 2013.
Resumen
Se evaluó la diversidad florística en doce fragmentos de bosque seco tropical de siete municipios del sur del departamento del Tolima (Colombia), en la Ecorregión Estratégica de la Tatacoa y su área de influencia del Alto Magdalena. Se estableció una red de 12 parcelas de 50 x 50 m (0,25 ha), una en cada fragmento, registrándose todos los individuos arbóreos con diámetro superior a 5 cm. Se encontraron 121 morfoespecies agrupadas en 41 familias. La riqueza varió entre 15 y 34 especies por parcela, lo que permitió clasificar las coberturas vegetales en diferentes categorías de diversidad según los índices de riqueza y uniformidad. Se determinó la riqueza estimada, cuyos máximos valores variaron entre 44 y 46 especies. Las parcelas con mayor número de especies únicas alcanzaron valores entre 9 y 11; mientras que los valores mayores de especies únicas con abundancia unitarias estuvieron entre 3 y 5. El modelo de abundancia relativa que mejor describe los bosques evaluados corresponde a la serie logarítmica, sin embargo, cuando corresponden a sucesiones tempranas la serie geométrica mejora considerablemente su ajuste. Los modelos Log-Normal y Broken Stick, tienden a ajustes intermedios. Por otro lado, las relaciones de diversidad beta para estos bosques no superaron el 50% de similaridad. Los resultados permitieron priorizar los bosques en cuanto a su diversidad florística y rareza como potenciales para la futura conservación y restauración del bosque seco tropical del Alto Magdalena.
Palabras clave: Alto Magdalena, Colombia, diversidad alfa, diversidad beta, especies únicas, modelos de abundancia, rareza de especies.
Abstract
Floristic diversity was evaluated in twelve tropical dry forest fragments of seven municipalities in southern Tolima (Colombia) in the strategic Tatacoa ecoregion and its area of influence in the upper Magdalena. A network of 12 plots of 50 x 50 m (0.25 ha), one in each fragment, was established, registering all individuals with dbh greater than 5 cm. 121 morphospecies were found grouped into 41 families. The richness ranged from 15 to 34 species per plot, allowing a classification of vegetation cover in different categories of diversity indices such as richness and uniformity. We determined the estimated richness, whose maximum values varied between 44 and 46 species. The plots with the highest number of unique species reached values between 9 and 11, while higher values of unitary abundantly unique species were between 3 and 5. The relative abundance model that best described the studied forests was the logarithmic series; however, with early succession plots the geometric series considerably improved fit. Log-normal and Broken Stick models tended to yield intermediate fits. Furthermore, the beta diversity relationships for these forests exceeded 50% similarity. The results permit a prioritization of these forests based on plant diversity and rarity in ters of their potential for future conservation and restoration of tropical dry forest of the upper Magdalena.
Key words: abundance models, alfa diversity, upper Magdalena, beta diversity, Colombia, unique species, species rarity.
INTRODUCCIÓN
Los bosques secos tropicales (bs-T), corresponden a ecosistemas complejos y frágiles, que albergan una riqueza única de biodiversidad y en los cuales la evaporación excede a la precipitación, presentando uno o dos periodos de sequía que pueden durar entre 4 a 6 meses en el año (Janzen 1986, Luttge 2008, Murphy y Lugo 1996), lo que genera déficit de agua en el suelo, cuya consecuencia es la defoliación de la vegetación como respuesta al estrés hídrico (IAvH 2002, Lobo et al. 2003 Murphy y Lugo 1996). Esto genera mayor diversidad estructural y fisiológica en formas de vida (Mooney et al. 1996), mezclando especies caducifolias con siempre verdes, constituyendo patrones complejos de tipo ecofisiológico (Burnham 1997).
En general, los bs-T tienen alta presión antrópica a tal punto que se consideran como de los ecosistemas más degradados a causa del cambio de uso de la tierra, asociado a la agricultura intensiva, como consecuencia de la alta fertilidad de sus suelos (Quesada y Stoner 2004). Los bs-T presentan bajas tasas de crecimiento en comparación con la vegetación que crece en el trópico húmedo, además, presentan ciclos reproductivos restringidos que incrementan su fragilidad, haciéndolos más susceptibles a la perturbación (Murphy y Lugo 1996). Una característica de los bs-T es que desafían el patrón de incremento en la riqueza de especies a medida que se acerca a la latitud o línea del ecuador (Chazdon y Denslow 2002), puesto que los ecosistemas de este tipo con mayor riqueza de especies son los que más alejados están de la línea ecuatorial, tales como los que se encuentran en México, Bolivia, Paraguay y Argentina (Gentry 1995). Igualmente, en los bs-T las áreas de mayor riqueza de especies corresponden a las más secas, como sucede en México y Bolivia (Gentry 1995, Lobo et al. 2003). Además, se caracterizan también por el alto número de endemismos florísticos que pueden estar entre el 43 y 73%, solamente superado por los bosques amazónicos (Kalascka 2004).
En Colombia, los bs-T se ubican por debajo de los 1.000 m de altitud con precipitaciones inferiores a los 2.000 mm/año, temperaturas superiores a 24° C y climadiagramas unimodales o bimodales, de acuerdo con su ubicación en los paisajes de la llanura Caribe, la cual tiene la mayor proporción de este tipo de ecosistema, cuya extensión supera los 1.300 km2. Igualmente, otra área de gran importancia cubierta de bs-T, son los valles interandinos de los ríos Magdalena y Cauca, que tienen la característica de ser sometidos al efecto sombra de los flancos de las cordilleras y posiblemente sean las áreas más afectadas por la actividad antrópica de tipo agropecuario (Cavelier et al. 1996, Linares y Fandiño 2009, Repizo y Devia 2008).
Con relación a la extensión del bs-T, para el valle seco del río Magdalena solo se tienen datos de los remanentes del norte de Tolima donde existen cerca de 31 fragmentos con tamaño promedio de 155,5 ha (IAvH 1998), y en estudios realizados sobre la caracterización biofísica del Ecorregión Estratégica de la Tatacoa (Universidad del Tolima 2002). Se estableció que para dicha región, las áreas cubiertas por bosques naturales son entre 1,5 y 2% de la superficie original que cubría una extensión aproximada de 80.000 km2 (Etter 1993). El estado del conocimiento del bs-T en Colombia es pobre, dado que son pocos los lugares donde existen inventarios completos o parcelas de investigación permanente. La literatura informa de inventarios de pocos grupos de especies y de escaza información sobre la dinámica del bosque, su biodiversidad y de las estructuras subyacentes del mismo (Bernate y Fernández 2002, Cabrera y Galindo 2006, IAvH 2002, Linares y Fandiño 2009). En general, se encuentran menos estudios sobre los bosques lluviosos tropicales e incluso menos que sobre los bosques andinos (Montagnini y Jordan 2005, Prance 2006, Sánchez-Azofeifa et al. 2005).
Los estudios sobre biodiversidad han cobrado gran importancia para este tipo de comunidades de bs-T porque muestran la compleja forma como las especies aprovechan los recursos abióticos y bióticos, y por lo tanto, pueden ser utilizados como herramienta de evaluación y comparación del efecto de las actividades humanas en estos ecosistemas, sobre todo, por la simplificación de la estructura biótica. De esta manera, la medida de la diversidad biológica se traduce en términos de cantidad y proporción de los elementos biológicos a diferentes escalas (Flynn et al. 2011, Magurran 2011), más aún cuando el crecimiento de la frontera agrícola, el cambio de uso del suelo y la deforestación de los bosques han generado la pérdida progresiva de la cobertura vegetal aumentando la fragmentación y pérdida de conectividad, poniendo en peligro los recursos naturales, y en especial, la biodiversidad, al disminuir la resiliencia del bosque alterando su potencial de restauración (Lamb 2005).
Siendo este tipo de ecosistema una prioridad para la tarea investigativa de la comunidad científica (Sánchez-Azofeifa et al. 2005), se planteó el presente estudio en las áreas de bs-T ubicadas en el sur del departamento del Tolima (Colombia), en la Ecorregión Estratégica de la Tatacoa, en el Alto Magdalena. Los objetivos del presente artículo son: 1) conocer la composición florística del área de estudio; 2) evaluar la diversidad alfa y el gradiente de variación intercomunitario o diversidad beta; 3) explorar la presencia de especies con abundancia y frecuencia única, en la cual se fundamenta la rareza de especies que contribuye al conocimiento de los endemismos locales; 4) caracterizar las comunidades bajo los patrones de los modelos de abundancia de especies; 5) contribuir al conocimiento del bs-T; y 6) ofrecer lineamientos de manejo y conservación para este ecosistema vulnerable y crítico en la región del valle seco del río Magdalena.
MATERIALES Y METODOS
Área de estudio. El estudio se realizó en fragmentos naturales de bosques secos tropicales (bs-T), con diferentes estados sucesionales, distribuidos en los municipios de Alpujarra, Coyaima, Dolores, Natagaima, Ortega, Prado y Purificación en el sur del departamento del Tolima (Colombia), y que hacen parte de la Ecorregión Estratégica de la Tatacoa (figura 1; tabla 1). Dicha área corresponde a una de las zonas semiáridas de mayor vulnerabilidad del país. El área de estudio se enmarca dentro de las coordenadas planas: X = 770.000-950.000 y Y = 760.000-950.000, con cotas inferiores a 1.000 m de altitud y con una extensión aproximada de 641.000 ha (Universidad del Tolima 2002). El climadiagrama de la región tiene comportamiento bimodal con dos periodos marcados de lluvia anuales, el primero de marzo a mayo y el segundo, entre octubre a diciembre, con valores anuales promedios que oscilan entre los 400 a 2.000 mm/año, para un promedio anual de 1.463 mm/año. La temperatura promedio anual es de 27,2 °C, siendo agosto el mes más cálido con 28,3 °C y noviembre el más fresco con 26,3 °C. La humedad relativa oscila entre 58 y 76,4%, siendo agosto y noviembre los extremos. El brillo solar es alto, supera las 1.960 horas/ año (Rojas 2001). En la región predominan suelos muy susceptibles a procesos erosivos (69%), con baja a muy baja fertilidad natural (85%) y una zonificación ecológica como área de conservación y reserva en más del 60% (Perea 2001).
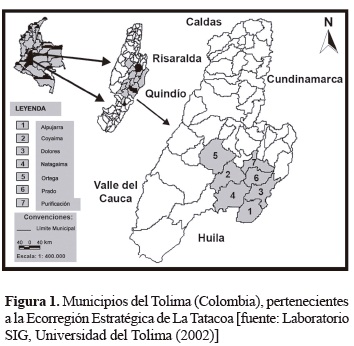
Metodología de campo. Se establecieron doce unidades de muestreo permanente en los fragmentos de bosque de 0,25 ha (50 x 50 m), con subparcelas de 10 x 10 m, teniendo como prioridad los individuos con habito arbóreo y arbustivo con diámetro normal superior a 5 cm (Bernate y Fernández 2002). Para cada individuo se recolectaron tres muestras botánicas, hasta donde fue posible en estado fértil. Posteriormente, se evaluaron de acuerdo con los descriptores morfológicos, el material fue organizado, empacado y prensado, para su traslado al Herbario de Dendrología de la Facultad de Ingeniería Forestal de la Universidad del Tolima y el Herbario Joaquín Antonio Uribe de Medellín (JAUM), donde fueron depositados y procesados con las técnicas convencionales de herborización. Las colecciones se identificaron por medio de comparaciones de herbario y catálogos de plantas vasculares.
Análisis de datos. En cuanto a índices de riqueza basados en la densidad de especies se seleccionó el índice de Margalef, se calculó el índice basado en la abundancia relativa de especies de Shannon (H') (Magurran 2004), por último el estimador de riqueza de especies basados en cobertura Chao 1 se seleccionó para estimar el potencial de riqueza. En cuanto a diversidad beta el gradiente de disimilaridad y complementariedad se determinó calculando la medida métrica de Jaccard y el índice de Sorensen para datos cuantitativos (Magurran 2004), en cuanto a rareza se hallaron las especies únicas, únicas unitarias, especies con abundancia uno y abundancia dos por sitio de muestreo, el procesamiento se llevó a cabo con el paquete informático EstimateS 6.0-b con especificación de límite superior de abundancia para especies raras e infrecuentes de 10 (Colwell 1997) y se revisó la información sobre estado de conservación de todas las especies encontradas y determinadas a nivel de especie. El estudio de diversidad beta se complementó con un análisis de conglomerados para las doce parcelas con las siguientes variables: número de individuos, número de especies, uniformidad de Shannon, recíproco de Simpson, índice alpha (α) y altura sobre el nivel del mar (Bernate y Fernández 2002), tales variables fueron escogidas tratando de recoger diferentes aspectos de la diversidad, relacionándolos a la vez con un aspecto propio pero independiente de los demás como es la altitud, para este análisis se utilizó el programa informático Infostat Profesional (Di Rienzo et al. 2008).
Las comunidades evaluadas, se caracterizaron utilizando las series propuestas por Magurran y McHill (2011), que corresponden a la serie Geométrica, serie Log, Serie Log-Normal Truncada y Broken Stick, con las respectivas pruebas de bondad de ajuste y de predicción (Flynn et al. 2011).
RESULTADOS
Composición florística. Se registraron 121 morfoespecies (Apéndice 1), cuyos valores variaron entre 8 y 34 especies por unidad de muestreo. Las parcelas ubicadas por debajo de los 400 m de altitud presentaron el mayor número de morfoespecies, correspondientes a los municipios de Ortega, Coyaima, Natagaima, Purificación y Prado (tabla 2).
Diversidad alfa (α) y beta (β). Para la riqueza de especies expresada en unidades de densidad por el índice de Margalef (Dmg) los valores mayores se generaron en las parcelas 2, 4, 7 y 10 con 6,44, 5,61, 5,86 y 5,90, respectivamente. En cuanto a índice de diversidad y heterogeneidad expresado por el índice de Shannon los valores menores se generaron en la parcela 6, definiendo en su orden las parcelas 2, 8, 9 y 10 como las más uniformes, es decir que las especies que conforman la comunidad biótica tienden a estar representadas por poblaciones de tamaño similar, mientras que las parcelas 1, 6 y 11 son descritas como las menos uniformes. De otro lado, Shannon muestra a las parcelas 1, 6 y 11 como las menos diversas, mientras que las parcelas 2, 4, 7 y 10 presentan la diversidad mayor. Teniendo en cuenta el posible estado de deterioro de los bosques, se determina en términos generales condiciones de diversidad pobre con respecto a las condiciones de diversidad óptimas que estos puedan presentar, se puede afirmar que los bosques descritos por las parcelas 1, 2, 6, 7, 10 y 11 tienen características importantes que les atribuyen prioridades de uso, manejo y conservación (tabla 2).
En cuanto a los valores de diversidad beta generados a partir de las comparaciones entre las diferentes parcelas, los valores de Jaccard clasifican las siete mejores comparaciones entre las parcelas 5-6, 2-4, 2-3, 5-7, 3-4, 7-12 y 6-9, respectivamente (tabla 3). Los valores para las medidas de Sorensen cuantitativo difieren un poco de Jaccard y define las mejores siete comparaciones entre las parcelas 5-6, 4-9, 2-3, 2-4, 3-4, 7-2 y 9-10, respectivamente (tabla 3), destacándose la primera de estas comparaciones por su valor alto (0,76), muy por encima del más cercano (0,36). Tal resultado muestra que estos dos sitios además de poseer un buen número de especies compartidas, también lo poseen en el número de individuos por especie, lo que las hace dos comunidades muy similares.
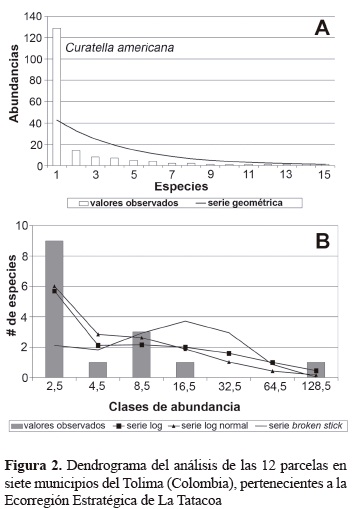
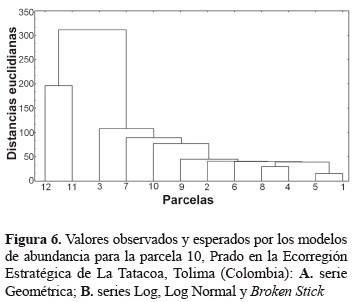
La figura 2 y la tabla 4 describen en forma tabulada y visual respectivamente las distancias euclidianas determinadas por el análisis de cluster, se encuentra que para las variables estudiadas existe gran similitud entre las parcelas 1 a 10, mientras que las parcelas 11 y 12 mantienen fuerte similitud entre sí y expresan diferencias marcadas con el primer grupo. Las parcelas más similares son 1 y 5, las cuales poseen valores idénticos para tres de las variables relacionadas como son número de individuos, número de especies e índice alpha. Posteriormente, se encuentran las parcelas 4 y 8, aunque con valores ya no tan semejantes y a este grupo luego se relacionan de una manera muy estrecha las parcelas 2, 6 y 9. Al relacionar los resultados anteriores con la altitud de las parcelas, se aprecia que el grupo que presenta mayor disimilaridad se ubica por encima de los 700 m de altitud, lo que sugiere coberturas que inician un ecotono al bosque premontano, por lo cual manifiestan la importancia de su conservación como hábitats sumideros para la conectividad entre el bs-T y el bosque premontano.
Rareza. En cuanto a las especies raras (tabla 2), las parcelas 7 y 10 manifiestan mayor cantidad de abundancias unitarias y presencia única, en contraste con la parcela 4 que posee el número mayor de especies con abundancia unitaria. En este aspecto, la parcela 6 también se destaca por la total ausencia de especies únicas. Se destaca la proporción de especies únicas y únicas con abundancia unitaria que alcanza el 47 y 17%, respectivamente de las 121 especies encontradas. Para el estimador de riqueza de especies Chao que se basa en las en la presencia de especies raras, se presentan los valores mayores en las parcelas 4 y 10.
Modelos de abundancia. Todas las parcelas se ajustan a las serie Log y Log Normal, en cuanto a la serie geométrica solamente cuatro parcelas (2, 8, 9 y 10) tienen similitudes con esta serie, la Broken Stick se ajusta a las parcelas 2, 9 y 10 (tabla 5). Al observar la serie geométrica se aprecia que en general las especies más abundantes en cada parcela son las que se interponen en su ajuste saliéndose abruptamente del patrón de incremento en abundancia, lo que las convierte en especies dominantes (figuras 3-6). El modelo de la serie Geométrica presentó los mayores ajustes en las parcelas 2, 9 y 10 (figuras 4-6; tabla 5). Según las pruebas de bondad de ajuste de Chi-cuadrado, la serie Log siempre logra ajustarse mejor que la Log Normal, alcanzando probabilidades de 0,72, 0,71 y 0,69 para las parcelas 6, 9 y 10, respectivamente (tabla 5).
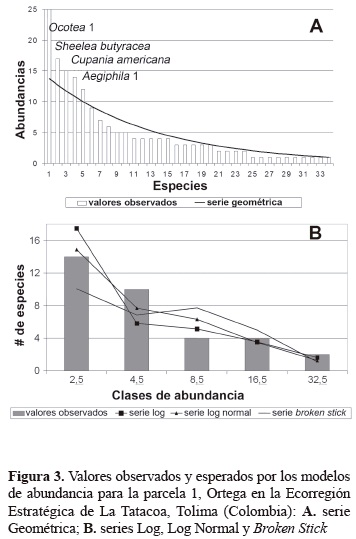
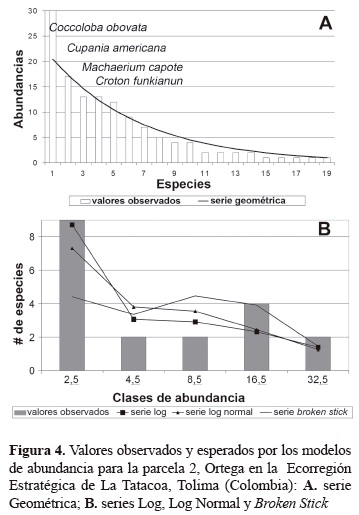
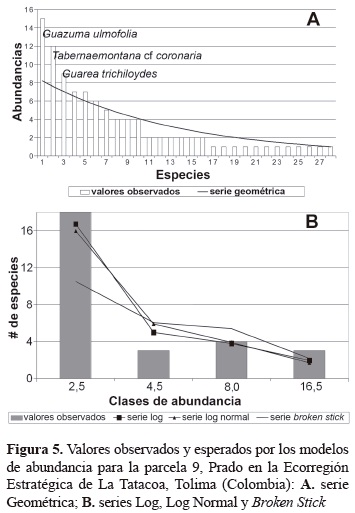
Las figuras 3 a 5, muestran el comportamiento de la serie Broken Stick, en el cual la tendencia de campana que esta suele tener difiere mucho con las distribuciones observadas, puesto que estas generalmente no poseen muchas especies en sus clases de abundancia intermedias. No obstante, para las parcelas 2, 9 y 10, la prueba de bondad de ajuste no encuentra diferencias significativas entre dicha serie (tabla 5). Las parcelas 2, 9 y 10 que brindan el mejor ajuste a las series Geométrica y Broken Stick, a su vez presentan un ajuste muy adecuado con las dos series restantes, sobre todo en la parcela 2, que obtiene la probabilidad mayor para la serie Broken Stick y para las series Log y Log Normal, obtiene probabilidades considerablemente buenas y muy similares entre sí (tabla 5). Tal coincidencia, para el caso de las series Log, Log Normal, y Broken Stick, se debe principalmente al número pequeño de clases de abundancia presentes en esta parcela, lo que hace el ajuste más fácil de realizar para todas las series (figura 3).
DISCUSIÓN
Composición florística. En este estudio se encontraron 121 morfoespecies agrupadas en 41 familias florísticas, lo cual supera ampliamente las 104 especies encontradas por Mendoza (1999) para el norte del departamento del Tolima (Colombia). El mismo autor informó para los bosques secos tropicales de la costa Caribe colombiana de 234 especies, pero con muestreos de individuos superiores a un centímetro de diámetro. Al discriminar estos resultados, las especies que corresponden a árboles y arbustos solo llegan a 52, cifra altamente superada por esta investigación, esto puede deberse en parte al tamaño del muestreo y al tipo de parcelas el cual es mucho más amplio en el presente estudio. Evaluaciones realizadas en bs-T en Puerto Rico por Murphy y Lugo (1986), encontraron 52 especies de árboles por hectárea con diámetro normal superior a 8 cm. Para el mismo bosque, dos estudios anteriores (Colon 2006, Quigley y Platt 2003) encontraron al género Coccoloba como uno de los más representativos de dichos bosques, el cual fue señalado en el presente estudio como dominante con una especie en un sitio de muestreo. En Jalisco (México), bs-T en áreas de conservación albergan más de 127 especies de árboles por hectárea con diámetros mayores a 10 cm (Burgos y Maass 2004), en los cuales los géneros Astronium, Cordia y Tabebuia son los más dominantes (Duran et al. 2006). En Nicaragua, Marín et al. (2009) encontraron 21 especies de árboles mayores a 10 cm de diámetro normal en parcelas de 0,7 ha, en las cuales el género Lonchocarpus fue uno de los más representativos. Powers et al. (2009), encontraron para un bs-T en Costa Rica 93 especies por hectárea para árboles mayores a 10 cm de diámetro normal, en los cuales Astronium, Brosimum y Spondias, son dominantes (Janzen 2002). Finalmente, estudios similares realizados por Parks et al. (2010) y Wishnie et al. (2007), en Panamá, encontraron de 28 especies de árboles por hectárea, en donde Guazuma es el género más representativo. Los géneros anteriores se encontraron en los bs-T de esta investigación y en algunas parcelas manifiestan comportamientos similares de dominancia, lo cual depende del estado de sucesión e intervención de cada sitio (Apéndice 1).
En cuanto a la diversidad de familia florísticas, Myrtaceae está representada por 12 morfoespecies, seguida de Fabaceae/ Mimosoide 8, Euphorbiaceae 7, posteriormente Anacardiaceae, Fabaceae/Papilionoide, Salicaceae, Moraceae y Rubiaceae con 5 especies. Estos resultados coinciden con lo señalado por Gentry (1995) y Mendoza (1999) para los bs-T colombianos, y Carrillo-Fajardo et al. (2007) para los bosques secos de Norte de Santander.
Diversidad alfa (α) y beta (β). Las parcelas 2 y 7 poseen la riqueza mayor de especies (34), al igual que los valores más altos para los índices de riqueza y uniformidad (tabla 2). La parcela 10 presenta el segundo valor más alto para diversidad, con dominancia mesurada, alto grado de uniformidad y riqueza estimada, además de valores excepcionales en cuanto a especies raras y únicas, que le permiten estimarle uno de los incrementos más altos en riqueza de especies según Chao 1 (tabla 2). Más del 50% de sus especies presentan abundancia menor o igual a 10, dentro de las cuales 9 son únicas (16%), lo cual vislumbra esta comunidad biótica como de interés para prioridades de conservación.
La diversidad beta expresada como la similaridaddisimilaridad entre los diferentes bosques evaluados, muestra tres grupos generales de coberturas, el primero lo constituyen los bosques representados en las parcelas 1, 2, 5, 6, 8 y 9, ubicados en los municipios de Ortega, Natagaima, Prado y Purificación. Estos bosques presentan la similaridad mayor tanto para Jaccard como Sorensen cuantitativo. Para el primer caso, la similaridad más alta alcanza valores superiores al 60% (parcelas 5 y 6), este conjunto de bosques se ubican entre 360 y 425 m de altitud. El segundo grupo, lo constituyen los bosques representados por las parcelas 3, 7 y 10 ubicadas en los municipios de Coyaima, Natagaima y Prado, con altitudes comprendidas entre 350 y 500 m. El tercer grupo, lo constituyen las parcelas 11 y 12, ubicadas en los municipios de Alpujarra y Dolores cuya altitud está por encima de 800 m (tabla 4). Esta clasificación (figura 2), vislumbra un complejo ecosistémico de complementariedad de comunidades vegetales que se hacen atractivas como prioridades de conservación debido a sus diferencias florísticas, consecuencia tanto de la heterogeneidad como por la rareza de sus especies, las cuales pueden ser utilizadas en procesos futuros de restauración como sugieren Burgos y Maass (2004) para paisajes naturales asociados a usos del suelo en bs-T ubicados en la costa oriental de México. Igualmente, los dos primeros conjuntos de bosques caracterizados tanto por la alta similaridad como ubicación geográfica adyacente, sugieren la ocurrencia de cierto grado de variación uniforme a través del espacio en la composición de especies y sus abundancias, lo que podría obedecer a su vez a condiciones de hábitat uniformemente variadas que quizá conduzcan a la determinación de una subregión biogeográfica específica, con características particulares de funcionalidad y dinámica (Butterfield 1995, Chazdon et al. 2010).
La combinación de las parcelas 5 y 7 según el índice de Jaccard muestra el valor mayor de similitud con el 50% de especies compartidas (tabla 4). Al comparar las relaciones entre las parcelas 5, 6 y 7, se puede deducir que la parcela 5 presenta características intermedias entre las otras dos, lo que en conjunto representa de una manera muy valiosa las diferentes etapas de desarrollo sucesivo que pueden ocurrir en un mismo bosque de poca extensión, ya que están ubicadas en la misma reserva indígena Anacarco. Dicha tendencia es muy similar con las parcelas 2, 3 y 4, que presentan el mismo comportamiento (tabla 4). Esta triple relación basa su similaridad tanto en el número de especies totales por parcela como en el número de especies y de individuos compartidos. Al hacer un reconocimiento de los medios por los que las medidas de diversidad beta definen las mejores relaciones, se revela una relación particular alterna para la parcela 4, que tiene similaridad con las parcelas 2, 3 y 9 menos apreciada por la medida cualitativa, pero valorada por la cuantitativa, al conservar una relación alta entre los individuos compartidos y el total de individuos de ambas parcelas, aunque solo comparte la mitad de la cantidad de especies que con las parcelas 2 y 3. Al tratar de relacionar y agrupar de manera consistente las parcelas restantes con base en la medida cualitativa y cuantitativa, solo las parcelas 9 y 10 presentan similaridad (tabla 3).
Las relaciones de similaridad dadas para las parcelas 5, 6 y 7, no deben ser tomadas muy en cuenta en comparación con las demás, ya que la corta distancia entre ellas sesga el resultado general de las comparaciones. Se sugiere en cambio que estas parcelas son una descripción amplia de un mismo sitio de muestreo, indudablemente requerida dadas las dimensiones de este bosque, las buenas condiciones presentes y la escasez de sitios de muestreo admisibles.
Si bien la importancia mayor en términos de biodiversidad indicadas por las medidas respectivas han sido destacadas, es preciso recalcar que todos los bosques estudiados poseen algún número de especies únicas, algunas de las cuales solo se registran con un individuo, y que dadas las reducidas extensiones de los bosques encontrados en la zona de estudio, les otorgan a estos toda la importancia que las medidas de diversidad no les reconozcan frente a otros ecosistemas como los andinos y los lluviosos tropicales.
Las variables utilizadas para producir el análisis de cluster son índices de diversidad alfa, complementadas por el número de individuos totales (usada por las medidas de riqueza, dominancia y uniformidad) y la altura sobre el nivel del mar. Lo anterior sugiere que la variable número total de individuos tiene gran importancia en la diferenciación de asociaciones vegetales (Bernate y Fernández 2002). De todas maneras, es un hecho que para este análisis las variables de altura sobre el nivel del mar y número total de individuos, consiguen modificar considerablemente las relaciones entre parcelas establecidas hasta el momento, al afirmar que las parcelas 5-1 y 8-4 poseen las similitudes más fuertes. El análisis cluster ratifica dos de las mayores similaridades establecidas por las medidas de diversidad beta (5-6 y 2-4). Esta confirmación obtenida es muy valiosa, si se tiene en cuenta que la influencia de las nuevas variables evaluadas, refuerza la declaración aceptada de la similitud existente entre estas parcelas. En términos prácticos, una apreciación más amplia de las agrupaciones observadas en el diagrama de árbol, permite rescatar las similitudes entre las parcelas 1, 2, 4, 5, 6, 8 y 9.
Si bien se acepta que los valores de riqueza y distribución de abundancias observados son en muchos casos similares a los encontrados en bosques en equilibrio estable, es un hecho que las labores de intervención como tala selectiva y pastoreo dentro de estos bosques han limitado constante y considerablemente su desarrollo hacia estados de diversidad mayor en toda la región neotropical (Portillo y Sánchez 2010).
Las relaciones de diversidad beta en su mayoría no superaron el 50% de similitud lo cual representa variabilidad alta entre comunidades. Sin embargo, los bosques de Ortega y Coyaima, caracterizados por las parcelas 2, 3 y 4, al encontrarse entre los menos disímiles tanto cualitativa como cuantitativamente, sugieren la ocurrencia de cierto grado de variación uniforme a través del espacio en la composición de especies y sus abundancias, lo que podría obedecer a su vez a condiciones de hábitat uniformemente variadas que quizá conduzcan a la determinación de otra subregión biogeográfica específica.
A pesar que las comunidades boscosas estudiadas se encuentran en un mismo rango altitudinal y por consiguiente en la misma zona de vida, existe una buena diferencia entre cada una de ellas en aspectos como dominancia, heterogeneidad, uniformidad, número y tipo de especies, lo que indica que estamos frente a una amplia diversidad de ecosistemas y etapas sucesionales del bosque. Este gradiente de asociaciones vegetales en bs-T no es muy conocido, como si lo es para ecosistemas de mayor humedad (Gentry 1988). Así lo investigaron Gentry (1995) y Mendoza (1999) en bosques secos del norte del Tolima y la costa Caribe, y Carrillo- Fajardo et al. (2007) en Cúcuta, los cuales aportan conocimientos de la amplia diversidad y endemismo dentro de cada región de bs-T en Colombia.
La descripción del bosque de la reserva indígena Anacarco en las parcelas 5, 6 y 7, es valiosa evidencia de la verdadera complejidad que pueden exhibir los bosques de la zona de estudio, si se tiene en cuenta la corta distancia entre tales parcelas para su relativa disimilitud encontrada. Lo anterior cobra mayor importancia si se considera la mínima proporción del bosque original representada por los relictos de bosque en los que se encuentra la totalidad de las parcelas, y por consiguiente la gran cantidad de las especies desaparecidas con los diferentes hábitat a través de los años. Este ejemplo a su vez justifica el valor de las estimaciones de riqueza y especies compartidas, al demostrar la existencia de especies adicionales no registradas, presentes fuera de las unidades de muestreo.
Rareza. La parcela 7 es de resaltar por sus siete abundancias dobles y diez abundancias unitarias dentro de las especies raras (más del 50% de sus especies con abundancia menor o igual a 10), dentro de las cuales nueve son únicas (16% del total de especies únicas). Los estudios revisados no presentan análisis de especies únicas, unitarias y únicas unitarias, esto no permite contrastar este importante dato para prioridades de conservación (tabla 2).
El listado de especies en peligro de extinción realizado por el Instituto Humboldt (Calderón 2010) incluye a las especies Anacardium excelsum, Astronium graveolens, Byrsonima crassifolia, Casearia megacarpa, Clorophora tintoria y Henrietella goudotiana, presentes en las parcelas del estudio y las categoriza en riesgo de extinción entre ''menor riesgo'' y ''en peligro''. Si bien la mayoría de tales especies en nuestro estudio se clasifican como muy raras ya sea por ocurrencia, abundancia, o por ambas características, solo representan un porcentaje pequeño del total de 57 especies únicas encontradas, dentro de las cuales 20 son de abundancia unitaria, sin contar otro número de especies de abundancia doble presentes en más de una parcela.
Lo anterior demuestra el nivel de desconocimiento que sobre estos bosques se tenía hasta el momento, sin mencionar la información perdida con la desaparición de gran parte de los mismos. Los sitios de rareza mayor registradas juegan un papel muy importante en el valor ecológico que estos bosques puedan tener, principalmente en consideraciones referentes al establecimiento de reservas naturales. De otro lado, especies raras en ocurrencia pero no así en abundancia, como es el caso de la especie Anaderananthera peregrina, que es de ocurrencia única y solo aparece en el bosque de Dolores (parcela 11), que constituye un relicto de muy limitada extensión, contrasta por su alta abundancia ubicándola como la especie con la segunda mayor población registrada en las 12 parcelas. Lo cual lo proyecta como un sitio de alto valor ecológico si se reconoce la gran cantidad de organismos tanto de fauna como de flora presentes en el mismo ecosistema que no solo se pueden beneficiar de su presencia, sino que pueden depender de ella, lo cual muestra un potencial para la revegetalización a partir de su regeneración natural o por plantación de material reproducido (Vieira y Scariot 2006).
Otras especies notificadas con algún grado de rareza son Attalea butyracea la cual se encuentra en categoría ''en preocupación menor nacional'' según UICN; Acanthocereus tetragonus incluida en el Apéndice II de CITES lo cual hace referencia a especies de las cuales se debe limitar su comercialización; Cedrela angustifolia la cual se encuentra en categoría VU A2 en la revisión de Pennington y Muellner (2010) y solo fue hallado un individuo en este estudio, el resto de especies únicas y unitarias no se encuentran en estudios de estado de conservación, lo cual alerta a tomar como prioridad estos factores de rareza, más que la simple riqueza de especies para definir sitios como prioridades de conservación.
Como en todo fenómeno o suceso natural los bosques sufren cambios a través del tiempo que por estar sujetos a innumerables factores son en general imposibles de predecir con precisión. Sin embargo, la necesidad de establecer acercamientos a estas predicciones obliga a realizar mediciones periódicas para crear modelos medianamente acertados sobre tales cambios en el tiempo. Las parcelas permanentes que se establecieron en este estudio fueron diseñadas para permitir la realización de estas mediciones a lo largo de los años. Entre más prolongado sea el estudio permanente en estos bosques, más precisas llegarán a ser las apreciaciones de su diversidad florística y aspectos funcionales como crecimiento y regeneración, que son en sí mismos los cambios que hacen variar tanto la riqueza como la heterogeneidad-dominancia de un bosque en sus procesos sucesionales.
Por tanto, es imprescindible pasar de una valoración o evaluación momentánea de la realidad como es la que presenta este estudio, a una valoración o evaluación dinámica, funcional y evolutiva que permita concebir, proyectar y predecir la realidad futura de estos bosques, por medio de la conservación de al menos las fracciones de bosques en las que se encuentran estas parcelas.
Modelos de abundancia. En cuanto a los modelos de distribución de abundancias, los bosques representados en las parcelas 2, 7, 9 y 10 son descritos bajo la serie geométrica (tabla 5), es decir que corresponden a comunidades donde unas pocas especies son dominantes y el resto son raras (Magurran 2004). Este modelo parte del principio ecológico que la abundancia de cada especie es equivalente a la proporción del recurso que esta utiliza (Krebs 1989). Igualmente, May (1981) afirmó que este tipo de modelo también cobija a las comunidades que estando en equilibrio estable sufren algún tipo de disturbio que estimula la dominancia de un grupo reducido de especies. Melo (2000), en fragmentos de bs-T en el norte del departamento del Tolima encontró que el modelo de la serie geométrica describe con buen ajuste a sucesiones tempranas dominadas por Cordia alliodora o Guazuma ulmifolia, que corresponden a especies dominantes en dos parcelas de este muestreo. Los demás bosques evaluados presentan buenos ajustes tanto al modelo logarítmico como log-normal (tabla 5). De acuerdo con Magurran (2004), estos modelos caracterizan comunidades donde las especies de abundancia intermedia son las más características (figura 3), es decir que las especies comunes son muy pocas, mientras que la mayoría de ellas son raras (Krebs 1989). En el norte del departamento del Tolima, estos modelos describen dentro de los bs-T a comunidades en estado de recuperación, al igual que bosques maduros con poca intervención antrópica (Melo 2000).
Al contrastar el ajuste de los modelos de abundancia para las parcelas 5, 6 y 7 de la reserva indígena Anacarco, es necesario anotar de manera general que para Magurran (2004), es frecuente encontrar buenos y confusos ajustes tanto de la serie Log como de la serie Log Normal, en parcelas cuyo tamaño no se considera suficientemente grande, lo que puede acomodarse en algunos casos a los resultados de este estudio, considerando que para todas las parcelas las pruebas de bondad de ajuste Chi-cuadrado encontraron diferencias no significativas. Con base en las pruebas de bondad de ajuste, las parcelas 5, 6 y 7 describen suficientemente bien las series Log y Log Normal (tabla 3), en donde a pesar de la diferencia entre cantidades totales de individuos, se guarda cierta proporción entre las clases de abundancia, de manera que la gran mayoría de las especies se encuentran en la primera clase, un número mucho menor de ellas en la segunda clase, un número mayor que el anterior en la tercera, y una cantidad de especies similar a la segunda en las clases superiores. Es de resaltar que la uniformidad de la parcela 7 le permite ajustarse casi perfectamente a un modelo geométrico en el que las especies más abundantes son varias y sus proporciones moderadamente diferenciadas. Atendiendo las apreciaciones de May (1981), quien afirmó que las comunidades que estando en un equilibrio estable sufren algún tipo de disturbio describen una distribución tipo serie Log a Geométrica, se puede afirmar entonces, que en general los bosques estudiados no se encuentran en su mejor estado de desarrollo sucesional y su distribución de abundancias sigue patrones Log y Log normal. Al igual que la parcela 2, la distribución de abundancias de la parcela 10 es descrita en forma casi perfecta por la serie Geométrica (figura 2), en parte debido a su gran uniformidad observada; uniformidad que al ser muy comparable con la de la parcela 9 (figura 4; tabla 3), la cual está ubicada a corta distancia de su siguiente le confiere a esta misma igual tipo de distribución de abundancias y con igual grado de ajuste. Una discusión y análisis gráfico más detallado de las distribuciones se puede observar en Bernate y Fernández (2002).
A manera de conclusión, los bosques evaluados comprenden una diversidad baja comparados con otros ecosistemas más húmedos, con tendencia a la homogeneidad y dominancia de algunas especies. Además, que presentan una diversidad beta alta en un gradiente de distancia horizontal y tres grupos marcados en el gradiente altitudinal.
El estudio cuantitativo de la rareza es el aporte mayor de este estudio y la recomendación de que es prioritario conservar bosques como los de la vereda el Diamante en Coyaima y los del resguardo indígena Anacarco. En cuanto a los modelos de abundancia, la mayoría son sitios de sucesiones intermedias altamente intervenidos que con un buen esquema de conservación podrían convertirse en refugios de la rareza alta de las especies encontradas.
Finalmente, los bosques de las parcelas 1, 2, 6, 7, 10 y 11 tienen características importantes que demandan acciones directas y gestión para su uso, manejo y conservación, por lo cual se recomienda que sean tenidos en cuenta para establecer sitios de conservación o áreas protegidas en cualquier categoría regional por parte del estado o reservas de la sociedad civil.
AGRADECIMIENTOS
A la Corporación Autónoma Regional del Tolima CORTOLIMA y la Facultad de Ingeniería Forestal de la Universidad del Tolima por la financiación del estudio por medio del proyecto ''Estudios de caracterización biofísica y socioeconómica de la Ecorregión Estratégica de la Tatacoa y su área de influencia''. A Jorge Enrique Céspedes por el apoyo y coordinación en el trabajo de campo de 15 meses. A Fernando Aly Huertas Gómez (q. e. p. d.) y Álvaro Cogollo del Herbario JAUM por el apoyo en determinación de especies. A los evaluadores anónimos de la revista Actualidades Biológicas quienes con sus observaciones permitieron consolidar un buen aporte al conocimiento de los bosques secos colombianos.
REFERENCIAS
Bernate-Peña JF, Fernández-Méndez F. 2002. Evaluación de la diversidad ecológica de las zonas áridas y semiáridas del área de influencia de la Ecorregión Estratégica de la Tatacoa en el Departamento del Tolima. [Tesis Ingeniero Forestal]. [Ibagué (Colombia)]: Facultad de Ingeniería Forestal, Universidad del Tolima. p. 115. [ Links ]
Burgos A, Maass JM. 2004. Vegetation change associated with land-use in tropical dry forest areas of Western Mexico. Agriculture, Ecosystems and Environment, 104: 475-481. [ Links ]
Burnham RJ. 1997. Stand characteristics and leaf litter composition of a dry forest hectare in Santa Rosa National Park, Costa Rica. Biotropica, 29: 387-395. [ Links ]
Butterfield RP. 1995. Promoting biodiversity: advances in evaluating native species for reforestation. Forest Ecology and Management, 75: 111-121. [ Links ]
Cabrera E, Galindo GA. 2006. Aproximación metodológica para la delimitación de ecosistemas de enclaves secos. Caso piloto: Cañones del río Dagua y del río Tuluá, Valle del Cauca-Colombia. Bogotá, D. C. (Colombia): Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. p. 45. [ Links ]
Calderón E. 2010. Listas rojas preliminares de plantas vasculares de Colombia, incluyendo orquídeas. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Fecha de acceso: 14 diciembre de 2011. Disponible en: http://www.humboldt.org.co/conservacion/plantas_amenazadas.htm. [ Links ]
Carrillo-Fajardo M, Rivera-Díaz O, Sánchez-Montaño R. 2007. Caracterización florística y estructural del bosque seco tropical del Cerro Tasajero, San José de Cúcuta. (Norte de Santander), Colombia. Actualidades Biológicas, 29 (86): 55-73. [ Links ]
Cavelier J, Ruiz A, Santos M, Quiñones M, Soriano P. 1996. El proceso de degradación y sabanización del valle alto del Magdalena. Neiva (Huila, Colombia): Fundación Alto del Magdalena. p. 92. [ Links ]
Chazdon RL, Denslow J. 2002. Floristic composition and species richness. In: Chazdon RL, Whitmore TC, editors. Foundation of tropical forest biology: classic papers with commentaries. Chicago (IL, U. S. A.): University of Chicago Press. p. 513-522. [ Links ]
Chazdon RL, Finegan B, Capers RS, Salgado-Negret B, Casanoves F, Boukili V, Norden N. 2010. Composition and dynamics of functional groups of trees during tropical forest succession in Northeastern Costa Rica. Biotropica, 41: 31-40. [ Links ]
Colon SM. 2006. Recovery of a subtropical dry forest after abandonment of different land uses. Biotropica, 38: 354-364. [ Links ]
Colwell RK. 1997. Estimates: statistical estimation of species richness and shared species from samples. Versión 5. User's guide and application. Fecha de acceso: 15 de diciembre de 2006. Disponible en: http://viceroy.eeb.uconn.edu/estimates. [ Links ]
Di Rienzo JA, Casanoves F, Balzarini MG, Gonzalez L, Tablada M, Robledo CW [Internet]. 2008. InfoStat, versión 2008. Argentina: Grupo InfoStat, Facultad de Ciencias Agropecuarias (Universidad Nacional de Córdoba). Fecha de acceso: 15 de diciembre de 2006. Disponible en: http://www.google.com.co. [ Links ]
Duran E, Meave JA, Lott EJ, Segura G. 2006. Structure and tree diversity patterns at the landscape level in a Mexican tropical deciduous forest. Boletín de la Sociedad Botánica de México, 79: 43-60. [ Links ]
Etter A. 1993. Diversidad ecosistémica en Colombia hoy. En: CEREC-Fundación Alejandro Ángel (FAA). Nuestra diversidad biológica. Bogotá (Colombia): CERECFAA. p. 43-61. [ Links ]
Flynn B, Mirotchnick N, Jain M, Palmer MI, Naeem S. 2011. Functional and phylogenetic diversity as predictors of biodiversity-ecosystem-function relationships. Ecology, 92 (8): 1573-1581. [ Links ]
Gentry AH. 1988. Changes in plant community diversity and floristic composition on environmental and geographical gradients. Annals of the Missouri Botanical Garden, 75: 1-34. [ Links ]
Gentry AH. 1995. Diversity and floristic composition of neotropical dry forest. In: Bullock S, Medina S, Mooney HA, editors. Tropical deciduous forest ecosystems. Cambridge: Cambridge UniversityPress. p. 116-194. [ Links ]
IAvH (Instituto Alexander von Humboldt). 1998. El bosque seco tropical (Bs-T) en Colombia. Programa de inventario de la biodiversidad, grupo de exploraciones y monitoreo ambiental GEMA. Villa de Leyva (Colombia): Instituto Alexander von Humboldt, IAVH. p. 24. [ Links ]
Armenteras D, Rodríguez N, Romero MH, Morales M, Cabrera E, Gottsman T, Franco C. 2002. Análisis preliminar de representatividad ecosistémica e identificación de vacíos de conservación y alternativas para el SIRAP del departamento del Valle del Cauca utilizando Sistemas de Información Geográfica. Informe de Resultados. Bogotá (Colombia): Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. p. 48. [ Links ]
Janzen DH. 1986. Tropical dry forests: the most endangered major tropical ecosystem. In: Wilson EO, editor. Biodiversity. Washington D. C.: National Academy Press. p. 130-137. [ Links ]
Janzen DH. 2002. Tropical dry forest: area de conservation Guanacaste, northwestern Costa Rica. In: Perrow M, Davy AJ, editors. Handbook of ecological restoration. Cambridge (England): Cambridge University Press. p. 559-583. [ Links ]
Kalascka M, Sánchez-Azofeifa GA, Calvo-Alvarado JC, Quesada M, Rivard D, Janzen DH. 2004. Species composition, similarity and diversity in three successional stages of a seasonally dry tropical forest. Forest Ecology and Management, 200: 227-247. [ Links ]
Lamb D, Erskine PD, Parrotta JA. 2005. Restoration of degraded tropical forest landscapes. Science, 310: 1628-1632. [ Links ]
Linares RJ, Fandiño MC. 2009. Estado del bosque seco tropical e importancia relativa de su flora leñosa, islas de la Vieja Providencia y Santa Catalina, Colombia, Caribe suroccidental. Revista de la Academia Colombiana de Ciencias, 33 (126): 1-12. [ Links ]
Lobo JA, Quesada M, Stoner KE, Fuchs EJ, Herrerias-Diego Y, Rojas-Sandoval J, Saborio-Rodriguez G. 2003. Factors affecting phenological patterns of bombacaceous trees in seasonal forests in Costa Rica and Mexico. American Journal of Botany, 90 (7): 1054-1063. [ Links ]
Luttge U. 2008. Physiological ecology of tropical plants. 2nd ed. Berlin (Germany): Springer-Verlag. p. 458. [ Links ]
Magurran AE. 2004. Measuring biological diversity. MA (U. S. A.): Blackwell Publishing Ltd. p. 256. [ Links ]
Magurran AE, McHill BJ. 2011. Biological diversity: frontiers in measurement and assessment. New York: Oxford University Press. p. 345. [ Links ]
Marin GC, Mulualem T, Gonzalez-Rivas B, Oden PC. 2009. A chronosequence analysis of forest recovery on abandoned agricultural fields in Nicaragua. Journal of Forestry Research, 20: 213-222. [ Links ]
May RM. 1981. Theoretical ecology: principles and applications. 2nd ed. Oxford: Blackwell Scientific Publications. p. 241. [ Links ]
Melo OA. 2000. Evaluación ecológica y silvicultural de los fragmentos de vegetación secundaria, ubicados en áreas de bosque seco tropical en el norte del departamento del Tolima. Ibagué (Tolima, Colombia): Facultad de Ingeniería Forestal, Universidad del Tolima. Informe técnico programa de investigación forestal en ecosistemas con tendencia a la aridez. p. 150. [ Links ]
Mendoza H. 1999. Estructura y riqueza florística del bosque seco tropical en la región Caribe y el valle del río Magdalena, Colombia. Caldasia, 21 (1): 70-94. [ Links ]
Montagnini F, Jordan CF. 2005. Tropical forest ecology: The basis for conservation and management. Berlin (Germany): Springer Verlag. p. 295. [ Links ]
Mooney HA, Bullock SH, Medina E. 1996. Introduction. In: Bullock SH, Mooney HA, Medina E, editors. 1996. Seasonally dry tropical forests. Cambridge (England): Cambridge University Press. p. 1-6. [ Links ]
Murphy PG, Lugo AE. 1986. Ecology of tropical dry forest. Annual Review of Ecology and Systematics, 17: 67-88. [ Links ]
Murphy PG, Lugo AE. 1996. Dry forests of Central America and the Caribbean. In: Bullock SH, Mooney HA, Medina E, editors. 1996. Seasonally dry tropical forests. Cambridge (England): Cambridge University Press. p. 9-34. [ Links ]
Parks A, van Breugel M, Ashton MS, Wishnie M, Mariscal E, Deago J, Ibarra D, Cedeno N, Hall JS. 2010. Local and regional environmental variation influences the growth of tropical trees in selection trials in the Republic of Panama. Forest Ecology and Management, 260: 12-21. [ Links ]
Perea JJ. 2001. Estado de los suelos de ecorregión estratégica de la Tatacoa. En: Caracterización biofísica y socioeconómica de la Ecorregión Estratégica de La Tatacoa y su área de influencia. Tomo III, Componente Suelos. Ibagué (Tolima, Colombia): Universidad del Tolima. p. 180. [ Links ]
Pennington TD, Muellner AN. 2010. A of monograph of Cedrela (Meliaceae). Milborne Port (England): DH Books. p. 112. [ Links ]
Portillo C, Sánchez-Azofeifa GA. 2010. Extent and conservation of tropical dry forests of the Americas. Biological Conservation, 143 (1): 144-155. [ Links ]
Powers JS, Becknell JM, Irving J, Perez-Aviles D. 2009. Diversity and structure of regenerating tropical dry forests in Costa Rica: environmental drivers and geographic patterns. Forest Ecology and Management, 258: 959-970. [ Links ]
Prance W. 2006. Tropical savannas and seasonally dry forests: an introduction. Journal of Biogeography, 33: 385-386. [ Links ]
Quesada M, Stoner KE. 2004. Threats to the conservation of the tropical dry forest in Costa Rica. In: Frankie GW, Mata A, Vinson SB, editors. Biodiversity conservation in Costa Rica: learning the lessons in a seasonal dry forest. Berkeley (CA, U. S. A.): University of California Press. p. 266-280. [ Links ]
Quigley MF, Platt WJ. 2003. Composition and structure of seasonally deciduous forests in the Americas. Ecological Monographs, 73: 87-106. [ Links ]
Repizo AA, Devia CA. 2008. Árboles y arbustos del valle seco del río Magdalena y la región Caribe colombiana: su ecología y usos. Bogotá, D. C. (Colombia): Facultad de Estudios Ambientales y Rurales, Pontificia Universidad Javeriana. p. 160. [ Links ]
Rojas PJ. 2001. Fauna y vida silvestre de la Ecorregión Estratégica de la Tatacoa. En: Caracterización biofísica y socioeconómica de la Ecorregión Estratégica de La Tatacoa y su área de influencia. Componente Fauna Tomo IV. Ibagué (Tolima, Colombia): Universidad del Tolima. p. 150. [ Links ]
Sánchez-Azofeifa A, Kalacska M, Quesada M, Calvo- Alvarado J, Nassa J, Rodríguez J. 2005. Need for integrated research for a sustainable future in tropical dry forests. Conservation Biology, 19 (2): 1-2. [ Links ]
Universidad del Tolima. 2002. Informe técnico proyecto ''Caracterización biofísica de la Eco-región Estratégica del desierto de la Tatacoa y su área de influencia''. Ibagué (Tolima, Colombia): Universidad del Tolima, Universidad Surcolombiana, Universidad de Cundinamarca, Cortolima, CAM. p. 250. [ Links ]
Vieira DLM, Scariot A, 2006. Principles of natural regeneration of tropical dry forests for restoration. Restoration Ecology, 14: 11-20. [ Links ]
Wishnie MH, Dent DH, Mariscal E, Deago J, Cedenoa N, Ibarra D, Condit R, Ashton, PMS. 2007. Initial performance and reforestation potential of 24 tropical tree species planted across a precipitation gradient in the Republic of Panama. Forest Ecology and Management, 243: 39-49. [ Links ]
