










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActualidades Biológicas
Print version ISSN 0304-3584
Actu Biol vol.36 no.100 Medellín Jan./June 2014
ARTÍCULOS DE INVESTIGACIÓN
Relaciones entre el peso seco y la longitud total de los géneros de invertebrados acuáticos Helobdella (Hirudinea: Glossiphoniidae) y Asellus (Crustacea: Asellidae) de un humedal andino de Colombia
Length–mass relationships in the aquatic invertebrate genera Helobdella (Hirudinea: Glossiphoniidae) and Asellus (Crustacea: Asellidae) of an Andean wetland of Colombia
John J. Rivera–Usme1, Gabriel A. Pinilla–Agudelo2, Diana L. Camacho–Pinzón3, María I. Castro–Rebolledo4, Jesús O. Rangel–Churio5
1 Calle 50, # 22–01. Armenia (Quindio), Colombia. jaderrivera@yahoo.com.ar.
2 Docente, Departamento de Biología, Universidad Nacional de Colombia. Bogotá, Colombia. gapinillaa@unal.edu.co.
3 Calle 50, # 22–01. Armenia (Quindio), Colombia. dianacamachop@hotmail.com.
4 Calle 22B, # 64–27, Torre 1, Apto. # 102. Bogotá, Colombia. micastro40@hotmail.com.
5 Instituto de Ciencias Naturales, Universidad Nacional de Colombia. Bogotá, Colombia. jorangelc@unal.edu.co.
Recibido: agosto 2013; aceptado: marzo 2014.
Resumen
Existen dificultades para determinar el peso de los macroinvertebrados y conocer así su importancia energética dentro de un ecosistema acuático. Por lo tanto, el objetivo de este estudio fue hallar relaciones matemáticas entre la longitud y la biomasa en dos géneros de invertebrados, representativos en el humedal Jaboque (Engativá) Bogotá D. C., Colombia, que faciliten hallar el peso de los individuos. Para ello, se realizaron cuatro muestreos desde abril de 2009 a enero de 2010. Se obtuvieron ecuaciones para estimar el peso seco a partir de medidas de la longitud total del cuerpo, para los géneros Helobdella y Asellus. Las relaciones halladas entre longitud total y peso seco fueron significativas y explicaron al menos el 69% de la varianza, expresada en los coeficientes de correlación (r2 = 0,69 y 0,85). Estos modelos permitirán calcular la biomasa para determinar el crecimiento y la producción secundaria de estos taxones en posteriores estudios de laboratorio o de campo. También podrían ayudar a conocer la importancia de estos organismos en el flujo de energía en los ecosistemas acuáticos, principalmente en los humedales andinos urbanos, donde son muy abundantes.
Palabras clave: Asellus, biomasa, Colombia, Helobdella, humedal andino Jaboque, invertebrados acuáticos, relación peso seco–longitud total.
Abstract
A major difficulty in the study of macroinvertebrates is the determination of their weight as a measure of the energetic importance of these organisms in an aquatic ecosystem. Therefore, the goal of this study was to find mathematical relationships between the length and biomass of two representative genera of aquatic invertebrates in the Jaboque (Engativá) wetland located in Bogotá, Colombia, to facilitate estimation of the mass of individuals. Four surveys were conducted from April 2009 to January 2010. Equations were derived to estimate dry weight from measurements of total body length for the genera Asellus and Helobdella. The length–mass relationships obtained were significant and explained at least 69% of the variance, expressed as the coefficients of correlation (r2 = 0.69 and 0.85). These models will allow for the calculation of biomass, and thereby help determine growth and secondary production of these taxa in subsequent laboratory or field studies. In addition, these equations will help to understand the importance of these organisms to energy flow through consumer pathways in aquatic ecosystems, particularly in urban Andean wetlands, where these organisms are very abundant.
Key words: Asellus, Helobdella, biomass, length–mass relationships, aquatic invertebrates, Andean wetland Jaboque, Colombia.
INTRODUCCIÓN
Los macroinvertebrados acuáticos se han usado ampliamente en estudios de bioindicación mediante la valoración de su composición y su diversidad (Rosemberg y Resh 1993). Sin embargo, en los últimos años se ha hecho mayor énfasis en su aporte en biomasa, lo cual permite establecer cuál es la contribución de esta comunidad en los flujos energéticos del sistema y la importancia que tienen para los demás eslabones tróficos del cuerpo de agua (Huryn y Wallace 2000). La biomasa es por lo tanto una medida que cuantifica la influencia de estos organismos sobre los procesos de transferencia de materia y energía en el ecosistema (Benke et al. 1999). Además, se reconoce que las estimaciones de biomasa son parte de los métodos para determinar la tasa de crecimiento y la producción secundaria de los macroinvertebrados acuáticos, así como para entender las historias de vida y las relaciones tróficas entre los grupos funcionales de alimentación (Benke 1996, Cyr et al. 1997).
Existen varios métodos para determinar la biomasa, tales como la medición del peso de forma directa en organismos frescos o preservados, la estimación del biovolumen y la elaboración de ecuaciones que relacionen la biomasa seca con la longitud de los organismos (Burgherr y Meyer 1997). La estimación indirecta de la biomasa en peso seco a partir de la longitud total del cuerpo es mucho más rápida que la determinación directa por gravimetría, particularmente para pequeños invertebrados como los macroinvertebrados acuáticos. Por supuesto, este tipo de técnicas indirectas dan una aproximación al peso seco, pero permiten evaluar la biomasa sin someter a los organismos a estrés y sin sacrificar toda la comunidad (Becker et al. 2009). Las determinaciones directas requieren una gran cantidad de muestras o especímenes recolectados, con lo cual, si hay muchos muestreos, pueden llegar a ser destructivos y nocivos para las poblaciones. A menudo esto hace que las mediciones directas de biomasa de invertebrados sean difíciles o imposibles (Sabo et al. 2002). Además, si se utilizan ejemplares preservados y se secan para pesarlos, es factible que se deterioren, lo que impide dejarlos en un museo o en una colección. Las técnicas más ampliamente aceptadas para la estimación de la biomasa involucran las regresiones entre el peso seco y la longitud del cuerpo de los organismos (Benke et al. 1999). Son numerosos los autores que han empleado esta relación entre morfología y peso de los invertebrados acuáticos bentónicos, por lo que solo se mencionan algunos como Meyer (1989), Nolte (1990), Smock (1980) y Towers et al. (1994). Miserandino (2001) obtuvo ecuaciones a nivel de órdenes de insectos (Ephemeroptera, Diptera, Plecoptera y Trichoptera) y relacionó la longitud total–peso seco con transformación logarítmica para datos de peso seco vs. largo del cuerpo de especímenes preservados. Aunque se dispone de algunos trabajos sobre insectos acuáticos del trópico (Cressa 1999, Tomanova et al. 2006), las investigaciones de este tipo son escasas en esta región del mundo, por lo que se hace necesario hallar ecuaciones basadas en las dimensiones corporales para dichas latitudes.
La información generada en el presente estudio constituye una primera aproximación para hallar, a partir del tamaño, la biomasa en grupos de hirudíneos e isópodos del humedal Jaboque. Se llena así un vació de conocimiento, ya que no se disponía de este tipo de ecuaciones para organismos de humedales andinos urbanos tropicales colombianos. De esta manera, las ecuaciones propuestas pueden remplazar las que se han usado hasta ahora, basadas en la literatura de zonas templadas, lo cual puede mejorar la exactitud en los cálculos y las posteriores interpretaciones que se deriven de ellos.
MATERIALES Y MÉTODOS
Descripción del área de estudio. El humedal Jaboque se ubica en la localidad de Engativá (Cundinamarca), Colombia, cerca del río Juan Amarillo (4° 43' 35,33'' N y 74° 08' 45,11'' O) y posee una extensión de 151,9 ha. Es un área inundable de la cuenca del río Bogotá, conformada por zonas permanentemente anegadas con espejos de agua y con vegetación flotante compuesta por Eichhornia crassipes Solms, Lemna gibba L., Limnobium laevigatum Humb. y Spirodela intermedia Koch. También se encuentran macrófitas emergentes (helófitas) como Juncus effusus L., Polygonum punctatum Elliott, y Typha latifolia L. y macrófitas sumergidas enraizadas (hidrófitas) como Myriophyllum aquaticum Verdc. (Rangel et al. 2005). Este humedal se localiza a 2.546 m. s. n. m., la precipitación media anual es de 792,8 mm y la humedad relativa es del 80%, con una temperatura media anual de 13,4 °C (Rangel y Orjuela 2003).
En general, las características físicas y químicas del humedal Jaboque se pueden resumir de la siguiente manera (Rivera 2011): es un sistema eutrófico, de aguas neutras a ligeramente ácidas (pH 6,44), bastante mineralizadas (conductividad 316 μScm–1), con temperaturas superiores a 15 °C en promedio, por lo cual se considera de aguas cálidas tropicales (Salas y Martino 2001). La demanda biológica de oxígeno (DBO5) es muy alta (92,9 mgl–1 O2 en promedio) e indica un elevado contenido de materia orgánica (MO); a su vez estos valores tan altos ocasionan que el oxígeno disuelto sea extremadamente bajo (0,89 mgl–1 en promedio, es decir el 12% de saturación). Los compuestos nitrogenados y el fósforo también son altos (promedios de 11,8 mgl–1 de NO3 y 1,78 mgl–1 de PO4). Los datos registrados para la mayor parte de las variables analizadas en el humedal superan los valores reglamentarios para consumo humano y para preservación de la biota acuática.
Todas estas características físicas y químicas son típicas de los humedales urbanos con altos aportes de aguas residuales. En el caso de Jaboque, estas aguas provienen de las actividades domésticas e industriales de la ciudad de Bogotá.
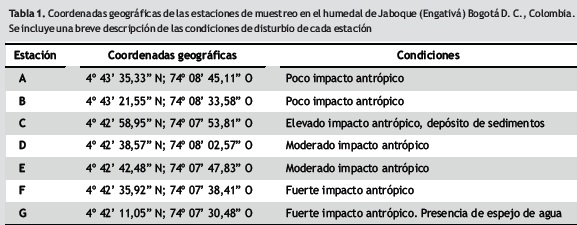
Muestreos y análisis de datos. Los muestreos se realizaron en abril y octubre de 2009 (meses de mayor precipitación) y en agosto de 2009 y enero de 2010 (meses de menor precipitación) en siete sitios del humedal Jaboque, seleccionados de acuerdo con el trabajo previo de Rangel (2005). Dos de estas estaciones estuvieron en áreas menos perturbadas (A, B), dos en zonas de impacto moderado (D, E) y tres en el sitios muy afectados por la urbanización y adecuación en su perímetro de canales de recolección de aguas residuales (C, F y G) (tabla 1). La recolección de los macroinvertebrados se hizo en zonas de macrófitas flotantes mediante una red de mano de 0,5 mm de ojo de malla. El área total muestreada fue de 0,126 m2 en cada zona. Esta red se ubicó bajo las plantas acuáticas, las cuales se extrajeron y se colocaron en bolsas plásticas. Las plantas se lavaron posteriormente para remover los organismos allí contenidos. Los invertebrados se fijaron con alcohol al 70% y se determinaron en el Laboratorio de Palinología del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia (ICN–UNAL); para la identificación de los géneros considerados en este trabajo se utilizaron las claves taxonómicas de Merritt y Cummins (1996) y Pennak (1978). Las mediciones de longitud corporal total de los individuos de los géneros Helobdella Blanchard 1896 y Asellus Geoffroy 1762 (familias Glossiphoniidae y Asellidae, respectivamente) se realizaron bajo estereoscopio con reglilla graduada en décimas de mm (precisión de 0,1 mm) en el Laboratorio de Ecología del Departamento de Biología (DB–UNAL). En los dos casos se midieron 50 individuos que incluyeron todos los tamaños capturados. Para hallar la biomasa en ''peso seco'' (PS) los organismos se secaron en una estufa (24 horas a 60 °C, Monzón et al. 1991) y luego se pesaron en una balanza de 0,0001 g de precisión en el Instituto de Genética Universidad Nacional (IG–UNAL). Los datos de biomasa se transformaron a logaritmo natural para su normalización y posterior análisis estadístico. Con el fin de obtener las ecuaciones para estimar el peso seco a partir de las medidas de la longitud total del cuerpo, se utilizaron modelos de regresión lineal teniendo en cuenta las consideraciones de Burgherr y Meyer (1997) y Miserendino (2001). Se empleó la ecuación general Y = a + b * X (donde: a y b son constantes, Y es el peso seco y X es la longitud). La versión linealizada del modelo se ajustó a los datos transformados a logaritmo mediante técnicas estándar de regresión. Para estos análisis se empleó el programa Statgraphics Centurión XV (StatPointTechnolgies 2006).
RESULTADOS Y DISCUSIÓN
Los ejemplares medidos de los dos grupos taxonómicos estuvieron comprendidos entre 3,4 y 13 mm de longitud total. El resumen estadístico de los datos de peso seco y longitud total de los organismos se muestra en la tabla 2. La regresión lineal simple entre la longitud total y la biomasa de las sanguijuelas del género Helobdella de la familia Glossiphoniidae (figura 1) tuvo un valor predictivo del 69% (r2 = 0,69; p < 0,05). La ecuación del modelo ajustado fue
Y = 7,49626 + 0,306511 * X
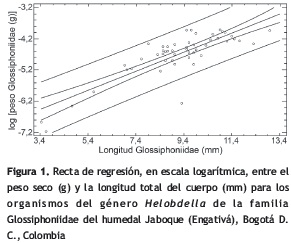
Learner y Potter (1974) desarrollaron una regresión para Helobdella stagnalis en un reservorio del sur de Gales en Inglaterra; emplearon el diámetro de la ventosa posterior y la biomasa en peso seco y obtuvieron un correlación alta (r = 0,97). Edwards et al. (2009) elaboraron ecuaciones para tres especies de sanguijuelas del norte de Europa, las cuales describen la relación entre la longitud del cuerpo y la biomasa (tanto en peso seco como húmedo); cuantificaron los efectos de la preservación en alcohol de estos especímenes y encontraron una reducción en la longitud del cuerpo y en la masa. Sin embargo, hallaron que los efectos sobre las relaciones de masa y longitud eran mínimos. Los especímenes más grandes de las tres especies tuvieron menos afectación por estas pérdidas. Estos autores recomiendan que las mediciones en muestras conservadas en alcohol se hagan después del primer mes de preservación, a fin de lograr la estabilidad en las variables de longitud y peso. Para el presente estudio, las mediciones se hicieron en muestras preservadas al menos 6 meses después de las capturas; es posible entonces que la preservación haya incidido en que la regresión solo llegara al 69% de explicación.
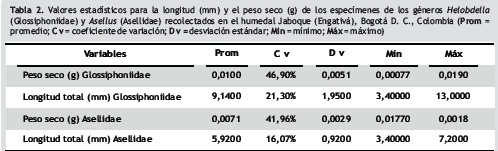
Los hirudíneos glosifónidos del humedal Jaboque se catalogan como depredadores y detritívoros (Rivera et al. 2013), ya que en sus contenidos estomacales se presentó un alto porcentaje de detritus orgánicos. Fueron muy numerosos en este ecosistema (1.587 ind/m2 en promedio, Rivera 2011) debido a que abunda la vegetación acuática, principalmente Eichhornia crassipes. Esto coincide con lo registrado por De Marco et al. (2001), quienes afirman que en zonas cubiertas por abundante vegetación hay predominio de este grupo de individuos. Autores como Haas et al. (2007) hallaron que el porcentaje de biomasa de hirudíneos en un humedal alemán fue de 1,6% de toda la fauna bentónica. En Jaboque, estos organismos constituyeron el 62% de la biomasa aportada por la comunidad de macroinvertebrados acuáticos (Rivera 2011), quizá debido a la gran cantidad de materia orgánica presente en este humedal. En el río Lambourn (Inglaterra), Tod y Schmid– Araya (2009) midieron la biomasa de los hirudíneos Glossiphonia complanata (0,01–0,05 gm–2), G. heteroclita (0,002–0,01 gm–2) y Erpobdella octoculata (0,001–0,004 gm–2). Los datos de dicho río son mucho menores a los registrados para Jaboque, donde el promedio de biomasa de Helobdella fue de 3,2 gm–2 (Rivera 2011). Los pesos secos de los glosifónidos en este humedal fluctuaron entre 0,00077 y 0,019 g (tabla 2).
Las diferencias en las abundancias y biomasas de los hirudíneos de zonas templadas con respecto a los registros en Jaboque quizá se deban a las explicaciones planteadas por autores como Tillman y Barnes (1973), quienes afirman que las oscilaciones en los ciclos de vida (y por ende en la biomasa) de Helobdella están fuertemente influenciadas por la temperatura. En las latitudes medias y altas los cambios de temperatura en el ciclo anual son muy fuertes, mientras que en la zona tropical esta variable tiene menos influjo, y es la variación en la precipitación la principal causa de las fluctuaciones ecológicas. Esto enfatiza la necesidad de desarrollar ecuaciones para los organismos de los ambientes tropicales, los cuales pueden responder de manera distinta a aquellos de los ecosistemas de la zona templada.
Para los isópodos de la familia Asellidae (figura 2) el valor predictivo de la regresión fue del 85% (r2 = 0,85; p < 0,05). En consecuencia, la ecuación resultante (Y = 7,72475 + 0,46904 * X) tiene una capacidad predictiva elevada. La correlación entre la longitud y el peso hallada en el presente trabajo (r = 0,92) es similar a la encontrada por Basset (1993) para Asellus en ríos de Inglaterra (r = 0,96). Este autor afirma que la disponibilidad de energía contenida en los isópodos, entendida como producción secundaria, está altamente relacionada con la polución química y con las elevadas concentraciones de nutrientes, las cuales influyen positivamente sobre las poblaciones de Asellus, tal como parece suceder en Jaboque.
La gran cantidad de individuos (283 inds/m2) y la importante biomasa de isópodos en Jaboque (11% de toda la biomasa de macroinvertebrados, Rivera 2011), parecen deberse a las condiciones claramente eutróficas de este humedal. En un humedal de Georgia (Estados Unidos) la biomasa de Asellidae llegó a 0,24 gm–2 (Martien y Benke 1977), mientras que Jaboque fue de 0,56 gm –2 . Chará et al. (2010) afirmaron que los isópodos pertenecen al grupo trófico de los fragmentadores (organismos que se alimenta de materia orgánica particulada gruesa) y que son muy importantes en términos de biomasa, ya que por tratarse de individuos de gran tamaño (en Jaboque sus pesos secos fluctuaron entre 0,0018 y 0,0177 g; tabla 2) contribuyen en mayor proporción al consumo del material vegetal, posiblemente en descomposición o detritus. Rivera et al. (2013) confirmaron esta apreciación en Jaboque y hallaron abundantes restos orgánicos y vegetales en los contenidos estomacales de estos organismos, que participan de esta manera con el reciclaje de nutrientes en el ecosistema. Isópodos e hirudíneos tienen un gran potencial ecológico para transformar los detritos de baja calidad y poca energía en biomasa de mejor calidad, representada en los cuerpos mismos de los organismos (Anderson y Sedell 1979, Gonçalves et al. 2004). Esta biomasa queda disponible para otros niveles tróficos o eslabones de la cadena alimentaria como aves y anfibios, o para otros invertebrados depredadores.
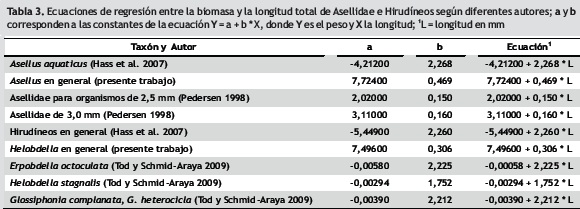
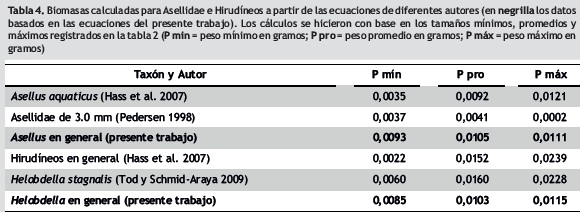
Al aplicar y comparar algunas de las ecuaciones desarrolladas en otras latitudes (tabla 3) a los datos de los invertebrados del humedal Jaboque (tabla 4), se observa que en el caso de los hirudíneos de las tallas medias y máximas la biomasa se ve sobreestimada cuando se emplean las ecuaciones de Hass et al. (2007) y de Tod y Schmid–Araya (2009), la primera desarrollada para un lago alemán y la segunda para un río de calizas en Inglaterra. Por su parte, la biomasa de los isópodos del género Asellus, calculada con las ecuaciones de Pedersen et al. (1998, la cual fue obtenida con animales cultivados en laboratorio en Dinamarca), y de Hass et al. (2007), tiende a estar subestimada para los individuos de Jaboque, en especial para las mismas clases de tallas mencionadas. Como se ve, las condiciones del humedal Jaboque (y posiblemente de otros humedales de la ciudad de Bogotá de similares características) determinan un comportamiento específico de la relación entre talla y peso de los organismos que no se pueden predecir acertadamente a través de las ecuaciones propuestas para otras regiones del mundo. Esto probablemente sea también cierto para otros grupos de invertebrados, lo que sugiere continuar con este tipo de investigaciones que permitan hallar regresiones ajustadas a las circunstancias de los ambientes acuáticos neotropicales.
Por supuesto, sería mucho más preciso desarrollar ecuaciones específicas para las especies, ya que su capacidad de predicción sería mayor (Cressa 1999). Por ello, las regresiones halladas en este trabajo se deben tomar como una primera aproximación para estos grupos de hirudíneos e isópodos de humedales andinos tropicales, de los cuales no se disponía de este tipo de ecuaciones. Las ecuaciones propuestas permitirán estimar la biomasa y la producción secundaria de estos taxones en ambientes tropicales similares al humedal Jaboque. Además, constituyen un punto de partida para posteriores investigaciones, tanto a nivel experimental de laboratorio como para trabajos de campo, ya que los cambios en la relación longitud–peso podrían dar una idea de las alteraciones humanas que causan modificaciones en estas comunidades. Tales variaciones permitirían valorar mejor los impactos sobre los ecosistemas, en especial si se trata de ambientes intervenidos como los humedales de la región andina.
AGRADECIMIENTOS
Los autores agradecen al Posgrado en Ciencias Biología, al Laboratorio de Genética y a los Laboratorios de Ecología Acuática y de Palinología (Universidad Nacional de Colombia, sede Bogotá) por las facilidades logísticas y por el préstamo de sus instalaciones para realizar las actividades de determinación y mediciones de longitud y peso de los organismos. Un reconocimiento especial a John Charles Donato por su orientación y a Silvia Morales de la Universidad Nacional de Colombia, por su colaboración en la medición de biomasa.
REFERENCIAS
Anderson NH, Sedell JR. 1979. Detritus processing by macroinvertebrates in stream ecosystems. Annual Review of Entomology, 24: 351–77. [ Links ]
Basset A. 1993. Resource–mediated effects of stream pollution on food absorption of Asellus aquaticus (L.) populations. Oecologia, 93 (3): 315–321. [ Links ]
Becker B, Moretti MS, Callisto M. 2009. Length–dry mass relationships for a typical Shredder in Brazilian stream (Trichóptera: Calamoceratidae). Aquatic Insects, 31 (3): 227–234. [ Links ]
Benke AC, Huryn AD, Smock LA, Wallace JB. 1999. Length–mass relationships for freshwater macroinvertebrates in North America with particular reference to the southeastern United States. Journal of the North American Benthological Society, 18 (3): 308–343. [ Links ]
Benke AC. 1996. Secondary production of macroinvertebrates. In: Hauer FR, Lamberti GA, editors. Methods in stream ecology. San Diego (U. S. A.): Academic Press. p. 557–578. [ Links ]
Burgherr P, Meyer E. 1997. Regression analysis of linear body dimensions vs. dry mass in stream macroinvertebrates. Archiv fuer Hydrobiologie, 139 (1): 101–112. [ Links ]
Chará AMS, Chará JD, Zúñiga MC, Pedraza GX, Giraldo LP. 2010. Clasificación trófica de insectos acuáticos en ocho quebradas protegidas de la ecorregión cafetera colombiana. Universitas Scientiarum, 15 (1): 27–36. [ Links ]
Cressa C.1999. Dry mass estimates of some tropical aquatic insects. Revista de Biología Tropical, 47 (1–2): 133–141. [ Links ]
Cyr H, Downing JA, Peters RH. 1997. Density–body size relationships in local aquatic communities. Oikos, 79 (2): 333–346. [ Links ]
De Marco P, Araujo MAR, Barcelos MK, Dos Santos MBL. 2001. Aquatic invertebrates associated with the water hyacinth (Eichhornia crassipes) in an eutrophic reservoir in tropical Brazil. Studies on Neotropical Fauna and Environment, 36 (81): 73–80. [ Links ]
Edwards FK, Lauridsen RB, Armand L, Vincent HM, Jones IJ. 2009. The relationship between length, mass and preservation time for three species of freshwater leeches (Hirudinea). Fundamental and Applied Limnology, 173 (4): 321–327. [ Links ]
Gonçalves Jr JF, Santos AM, Esteves FA. 2004. The influence of the chemical composition of Typha domingensis and Nymphaea ampla detritus on invertebrate colonization during decomposition in a Brazilian coastal lagoon. Hydrobiologia, 527: 125–137. [ Links ]
Haas K, Köhler U, Diehl S, Köhler P, Dietrich S, Holler S, Jaensch A, Niedermaier M, Vilsmeier J. 2007. Influence of fish on habitat choice of water birds: a whole system experiment. Ecology, 88 (11): 2915–2925. [ Links ]
Huryn AD, Wallace JB. 2000. Life history and production of stream insects. Annual Review of Entomology, 45: 83–110. [ Links ]
Learner A, Potter D. 1974. Life–history and production of the leech Helobdella stagnalis (L.) (Hirudinea) in a shallow eutrophic reservoir in South Wales. Journal of Animal Ecology, 43 (1): 199–208. [ Links ]
Martien RF, Benke AC. 1977. Distribution and production of two crustaceans in a wetland pond. American Midland Naturalist, 98 (1): 162–175. [ Links ]
Merritt RW, Cummins KW. 1996. An introduction to the aquatic insects of North America. 3rd ed. Dubuque (U. S. A.): Kendall/Hunt. p. 862. [ Links ]
Meyer E. 1989. The relationship between body length parameters and dry mass in running water invertebrates. Archiv fuer Hydrobiologie, 117 (2): 191–203. [ Links ]
Miserendino ML. 2001. Relaciones longitud–peso para macroinvertebrados de ambientes dulceacuícolas de Patagonia (Argentina). Ecología Austral, 11 (1): 3–8. [ Links ]
Monzón A, Casado C, Montes C, García De Jalón D. 1991. Organización funcional de las comunidades de macroinvertebrados acuáticos de un sistema fluvial de montaña (Sistema Central, río Manzanares, España). Limnetica, 7: 97–112. [ Links ]
Nolte U. 1990. Chironomid biomass determination from larval shape. Freshwater Biology, 24 (3): 443–451. [ Links ]
Pedersen CL. 1998. A simple device for sorting live benthic invertebrates into size groups. Hydrobiologia, 368: 61–63. [ Links ]
Pennak RW. 1978. Fresh–water invertebrates of the United States. 2nd ed. Nueva York (U. S. A.): Jhon Wiley. p. 803. [ Links ]
Rangel JO, González C. Parra LN. 2005. Biodiversidad del humedal Jaboque. Hacia su restauración ecológica. Cartilla de Divulgación. Bogotá (Colombia): Convenio Empresa de Acueducto y Alcantarillado de Bogotá, Universidad Nacional de Colombia, Universidad Colegio Mayor de Cundinamarca. p. 22. [ Links ]
Rangel JO, Orjuela MA, editores. 2003. El Humedal de Jaboque. Bogotá (Colombia): Instituto de Ciencias Naturales. p. 15. [ Links ]
Rangel JO. 2005. Investigación aplicada en restauración ecológica en el humedal de Jaboque. Convenio de cooperación científica y técnica entre la Empresa de Acueducto y Alcantarillado de Bogotá y la Universidad Nacional de Colombia. Informe Final. Bogotá (Colombia): Empresa de Acueducto y Alcantarillado de Bogotá (EAAB), Universidad Nacional de Colombia (UN), Bogotá. p. 935. [ Links ]
Rivera JJ, Pinilla GA, Camacho DL. 2013. Grupos tróficos de macroinvertebrados acuáticos en un humedal urbano andino de Colombia. Acta Biológica Colombiana, 18 (2): 43–56. [ Links ]
Rivera JJ. 2011. Relación entre la composición y biomasa de la comunidad de macroinvertebrados acuáticos y las variables físicas y químicas en el humedal Jaboque Bogotá–Colombia (,Tesis de Maestría). (,Bogotá (Colombia)): Universidad Nacional de Colombia. p. 174. [ Links ]
Rosemberg D, Resh V, editors. 1993. Freshwater biomonitoring and benthic macroinvertebrates. New York: Chapmann and Hall. p. 488. [ Links ]
Sabo JL, Bastow JL, Power ME. 2002. Length–mass relationships for adult aquatic and terrestrial invertebrates in a California watershed. Journal of the North American Benthological Society, 21 (2): 336–343. [ Links ]
Salas HJ, Martino P. 2001. Metodologías simplificadas para la evaluación de eutroficación en lagos cálidos tropicales. Lima (Perú): Centro Panamericano de Ingeniería Sanitaria y Ciencias del Ambiente, Programa Regional CEPIS/HPE/OPS OPS/CEPIS/PUB. p. 60. [ Links ]
Smock LA. 1980. Relationship between body size and biomass of aquatic insects. Freshwater Biology, 10 (4): 375–383. [ Links ]
Statpoint Technologies Inc. 2006. Statgraphics Centurion XV (programa de ordenador). Virginia (U. S. A.): Stat Point Inc. [ Links ]
Tillman DL, Barnes JR. 1973. The reproductive biology of the leech Helobdella stagnalis (L.) in Utah Lake, Utah. Freshwater Biology, 3 (2): 137–145. [ Links ]
Tod SP, Schmid–Araya JM. 2009. Meiofauna versus macrofauna: Secondary production of invertebrates in a lowland chalk stream. Limnology and Oceanography, 54 (2): 450–456. [ Links ]
Tomanova S, Goitia E, Helesic J. 2006. Trophic levels and functional feeding groups of macroinvertebrates in neotropical streams. Hydrobiologia, 556: 251–264. [ Links ]
Towers DJ, Henderson IM, Veltman CJ. 1994. Predicting dry weight of New Zealand aquatic macroinvertebrates from linear dimensions. New Zealand Journal of Marine and Freshwater Research, 28 (2): 159–166. [ Links ]
