










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActualidades Biológicas
Print version ISSN 0304-3584
Actu Biol vol.37 no.102 Medellín Jan./June 2015
ARTÍCULOS DE INVESTIGACIÓN
Sexual maturity and sexual dimorphism in a population of the rocket-frog Colostethus aff. fraterdanieli (Anura: Dendrobatidae) on the northeastern Cordillera Central of Colombia
Madurez y dimorfismo sexual de la ranita cohete Colostethus aff. fraterdanieli (Anura: Dendrobatidae) en una población al este de la Cordillera Central de Colombia
Yeison Tolosa1,2, *, Claudia Molina-Zuluaga1,4,*, Adriana Restrepo1,5, Juan M. Daza1,**
1 Grupo Herpetológico de Antioquia, Instituto de Biología, Universidad de Antioquia, AA 1226, Medellín, Colombia. * These authors contributed equally.
E-mails (**corresponding author): 2 yortegon@ut.edu.co; 3 clamozu@gmail.com; 4 restrepoadriana78@gmail.com; ** juanm. daza@udea.edu.co
Recibido: febrero 2014; aceptado: septiembre 2014.
Abstract
The minimum size of sexual maturity and sexual dimorphism are important life history traits useful to study and understand the population dynamics of any species. In this study, we determined the minimum size at sexual maturity and the existence of sexual dimorphism in a population of the rocket-frog, Colostethus aff. fraterdanieli, by means of morphological and morphometric data and macro and microscopic observation of the gonads. Females attained sexual maturity at 17.90 ± 0.1 mm snout-vent length (SVL), while males attained sexual maturity at 16.13 ± 0.06 mm SVL. Females differed from males in size, shape and throat coloration. Males were smaller than females and had a marked and dark throat coloration that sometimes extended to the chest, while females lacked this characteristic, with a throat either immaculate or weakly pigmented. In this study, we describe some important aspects of the reproductive ecology of a population of C. aff. fraterdanieli useful as a baseline for other more specialized studies.
Key words: Amphibian, Andes, gonads, histology, morphometry, reproduction
Resumen
El tamaño mínimo de madurez sexual y el dimorfismo sexual son importantes características de historia de vida, útiles para estudiar y comprender la dinámica poblacional de cualquier especie. En este estudio, determinamos el tamaño mínimo de madurez sexual y la existencia de dimorfismo sexual en una población de la ranita cohete, Colostethus aff. fraterdanieli, a través de datos morfológicos y morfométricos y de observaciones macro y microscópicas de las gónadas. Las hembras alcanzan la madurez sexual a los 17,90 ± 0,1 mm de longitud rostro cloaca (SVL), mientras que los machos alcanzan la madurez sexual sobre los 16,13 ± 0,06 mm (SVL). Las hembras difirieron de los machos en tamaño, forma y coloración de la garganta. Los machos fueron más pequeños que las hembras y tuvieron una marcada y oscura coloración gular que algunas veces se extiende hasta el pecho, mientras que las hembras carecen de esta característica, con una garganta inmaculada o débilmente manchada. En este estudio, describimos algunos aspectos importantes de la ecología reproductiva de una población de C. aff. fraterdanieli útiles como línea base para otros estudios más especializados.
Palabras clave: Andes, anfibios, gónadas, histología, morfometría, reproducción
INTRODUCTION
Reproductive characteristics are critical components in the life history of any organism, and constitute an important aspect of the population dynamics of any species (Morrison et al. 2004, Watling and Donnelly 2002). The minimum size of sexual maturity (MSSM, the size at which an individual has all the morphological and physiological conditions to begin to breed; Morrison and Hero 2003, Stearns 1992) along with sexual dimorphism (SD, the existence of morphological differences between sexes; Shine 1979), are important life history traits useful to study and understand the population changes through time.
Different theories have been suggested to explain the evolutionary mechanisms behind the MSSM and SD (Lovich et al. 2014, Shine 1979). Life history theory explains the variation in MSSM between sexes, populations and species through natural selection mechanisms mainly related with adult mortality rates (Roff 1992, Stearns 1992). Furthermore, SD is usually explained through two processes: sexual selection (e.g., intrasexual competition for mates, epigametic selection; Lovich and Gibbons 1992) and natural selection (differential interactions of each sex with its environment; Lovich and Gibbons 1992). Both processes can operate independently or in a combination, working together or sequentially to promote a specific degree or type of SD (Shine 1989). However, ecological differences may also be forced as a consequence of sexually selected dimorphism or vice versa (Lovich et al. 2014). Other non-adaptive theories have been suggested, including body-size scaling, genetic correlations between male and female body size, and phylogenetic constraints/ inertia (Lovich et al. 2014).
Amphibians and particularly anurans display an extraordinary diversity of life histories and reproductive strategies, which include since species with unattended eggs deposited in water bodies to develop into aquatic larvae until species with terrestrial eggs and direct development, passing by a wide range of mating and parental behaviors (Duellman and Trueb 1994). This diversity has limited the finding of clear patterns within life history of amphibians, and the ability to generate hypotheses about its occurrence. However, has been found that most amphibian females have larger body sizes than males and thus females usually reach sexual maturity at larger sizes (90% of evaluated species; Shine 1979, Woolbright 1983). Several explanations have been suggested for this pattern: the first one is related to a higher fertility in larger females (Shine 1989, Woolbright 1983). The second one refers to energetic constrains in males because territorial defense, agonistic behavior, and advertising behavior may require energy that would otherwise be used for growth (Woolbright 1983). Another explanation is usually appealed in species where males are larger than females and refers to the existence of physical male combats for mating because large males are more likely to win combats (Katsikaros and Shine 1997, Shine 1979, Wells 1978).
Some studies show the existence of chromatic SD in some amphibian species, where males and females exhibit different colorations patterns in particular body areas like dorsum (Buchanan 1994, Rojas and Endler 2013), upper jaw (Buchanan 1994), vocal sac (Buchanan 1994), hidden surfaces of forelimbs (e.g., Pristimantis bicolor, Rueda- Almonacid and Lynch 1983; Pristimantis erythropleura, Lynch 1996) and fingers (Allobates caeruleodactylus, Lima et al. 2002). However, because these traits are not conservatives across different taxa have received little attention. A more conservative coloration SD between different anuran species is the throat coloration, which is dark, or dark spotted in males whiles is clear or immaculate in females (Grant and Castro 1998, Greenberg 1942, Wells 1978).
In this study, we describe some aspects of the reproductive ecology of Colostethus aff. fraterdanieli, specifically we focus on the following objectives: 1) Determine the minimum size at sexual maturity in males and females and 2) establish if exist some sexual dimorphism in this species. Keeping in mind the general trends of anurans respect these two life history traits, we predicted that females would reach MSSM at larger sizes than males. Additionally, we expected that this species present sexual dimorphism with females larger than males and a darker throat coloration pattern in males.
MATERIALS AND METHODS
Study area. The population is located in the site known as El Edén, a secondary forest in ''vereda San Antonio'', Alejandria municipality, Antioquia, Colombia (6,36664 N, 75,02746 W), in the buffer area of the Jaguas hydroelectric project. The area corresponds to pre-montane wet forest (according to the system of Holdridge 2000) and is located at 1400 m. a. s. l. at the northeastern of the Cordillera Central in Colombia. Temperature varies between 18 and 24 °C, with an average relative humidity of 82.2%, and annual rainfall between 2000 and 4000 mm.
Study species. Colostethus aff. fraterdanieli is an undescribed species sister to C. fraterdanieli with 13% of genetic distance in the mitochondrial gene COI (Daza et al. unpublished). The genus Colostethus is a trans- Andean clade, extending from eastern Central America to northwestern Ecuador, with most species occurring in cloud forests in the western Andes (Grant et al. 2006). Colostethus aff. fraterdanieli is a leaf-litter dwelling frog inhabiting forest fragments with small streams. Like its congeners, this species is extremely fast and agile, responding to danger by hopping a short distance and hiding underneath the leaf litter. Males call constantly from leaf litter. The advertisement call is composed by one note and in some cases; males emit calls with three notes. Adults seem to be territorial and males have been observed carrying 10 to 11 tadpoles in their backs.
Field work. Frogs were collected between March and May 2013 using active search (Crump and Scott 2001). Although it could be useful to collect the animals throughout the year to record the temporal and seasonal variation in reproductive status of the frogs, we had to restrict the fieldwork to March and May due to some logistical difficulties. All collected individuals were euthanized using lidocaine 3%, fixed with formaldehyde 10% and stored in the herpetological collection of the Museo de Herpetología de la Universidad de Antioquia (MHUA; vouchers MHUA-A 7878-7898, 7900-7929, 7931-7933, and 7940-7984).
Minimum size of sexual maturity (MSSM). All individuals were dissected under a stereomicroscope to examine gonadal characteristics and to determine the sex and the reproductive stage. The sex of the individuals were established by the presence of ovaries or testes. The females were classified according to the following reproductive stages: previtellogenic (without vitellogenic follicles), vitellogenic (with brown follicles), and gravid (with oviductal eggs; Méndez-de la Cruz et al. 2013). The females were considered matures when presented vitellogenic follicles, eggs, or when presented at the same time previtelogenic follicles and convoluted oviducts (Prado et al. 2004, Wake and Dickie 1998). We prepared histological slides of testes sections stained with the hematoxylin-eosin (Luna 1968) to classified males like matures or immature. We used the presence of spermatozoa in the seminiferous tubules to identify adult males (Prado et al. 2004, Wake and Dickie 1998). We inferred the MSSM on each sex as the size of the mature individual with the lowest SVL.
Sexual dimorphism (SD). Prior to dissection, frogs were photographed and measured to determine the presence of sexual dimorphism. Only mature individual were included in the analysis of SD. All measurements were taken three times at the stereomicroscope with a digital caliper (0.1 mm precision) and then were averaging to reduce type II errors (Yezerinac et al. 1992). Characters measured were: snout-vent length (SVL), head length (HL), head width (HW) measured at the level of the posterior part of the tympanum, inter-orbital diameter (IOD), inter-nostril distance (IND), eye diameter (ED), disc diameter of the third digit (DD3), disc diameter of the fourth toe (DF4), eye-nostril distance (END), tympanum diameter (TD), length of the humerus (LHU), forearm length (FOL), thigh length (THL), tibia length (TL), and foot length (FL) (figure 1). These characters correspond to the typical measures used to determine morphometric differences in anurans (e.g., Felgueiras-Napoli 2005).
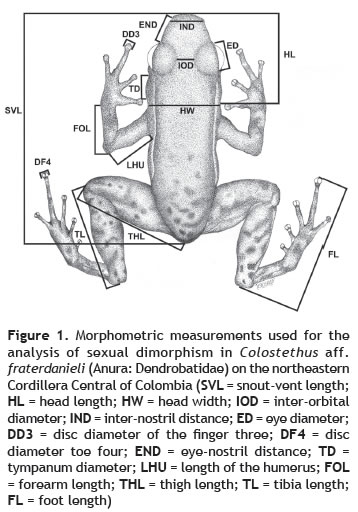
We used a principal components analysis on the covariance matrix of all morphological measurements to reduce the total morphometric variation in few dimensions using the software Past v.3.01 (Hammer et al. 2001). The first principal component (PC1) calculated from a set of morphometric measurements generally expresses body size variation when all traits load largely and in the same direction (Reyment et al. 1984). Remaining variance describes relative shape differences that are not due to body size (and these are expressed in subsequent PCs). The relative contribution of the original variables to each principal components were determined from the matrix of factor-variable correlations (table 1). We evaluated sexual dimorphism in size (PC1) and shape (PC2) using one-way ANOVAs.
In addition to morphometric variation, we classified all mature individuals according to throat coloration. Individuals with spots covering less than 20% of the throat were classified as ''immaculate'' and the remaining individuals were classified as ''spotted''. We used a chi-square test (χ2) to test for differences between sexes in this feature.
RESULTS
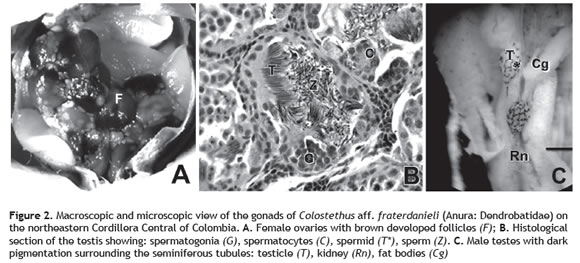
Minimum size of sexual maturity (MSSM). We examined 101 frogs and based on external observation of the gonads and histological evaluation of the testes: We found 22 matures females, 24 immature females, 53 matures males and two immature males. MSSM in females was at 17.90 ± 0.1 mm. All mature females had between 8-10 brown vitellogenic follicles and convoluted oviduct. We did not find any female with eggs in the oviduct (figure 2A). The MSSM in males was 16.13 ± 0.06 mm, and all showed seminiferous tubules with cysts that had most of the stages of spermatogenesis (spermatogonia, spermatocytes, spermatids and sperm). All mature males had free spermatozoa in the seminiferous tubules (figure 2B). The right testis was larger than the left one, and in some individuals, testes had dark pigmentation surrounding the seminiferous tubules (figure 2C). This pigmentation seems not related with the reproductive status of individual.
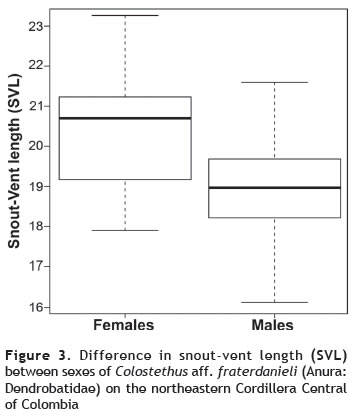
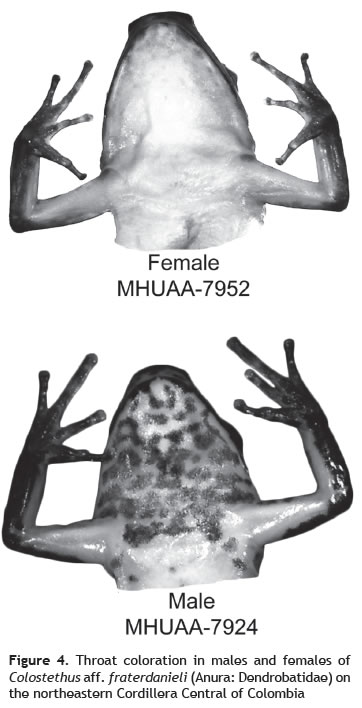
Sexual dimorphism (SD). PCAs showed that the subspaces defined by the first two principal components had the same orientation in both sexes and can be used as size and shape variables. Specifically, the sums of the eigenvalues (S) were 0.023 of a possible range of 0 to 2, where 2 indicates identical subspaces (Krzanowski 1979). The morphometric analysis showed differences in size and shape between males and females (size: df = 1, F = 24.33, p < 0.001; shape: df = 1, F = 109.9, p < 0.001). Females were bigger than males (figure 3). We also found differences between sexes in the throat coloration (df = 1, χ2 = 41.57, p < 0.001). Most of the mature males had the spotted pattern (90%). The two immature males lacked this coloration. All females had an immaculate pattern on the throat (figure 4).
DISCUSSION
In this study we describe some aspects of the reproductive ecology of a population of C. aff. fraterdanieli, we show that females and males differ in MSSM, size, shape, and throat coloration. Females reach sexual maturity around 17.9 mm while males reach it at a smaller size of about 16.1 mm. Due to the great diversity of life histories and reproductive strategies recorded to the dart-poison frogs, this particular group of anurans have attracted the attention of numerous researchers evaluating different reproductive and behavioral characteristics. An important part of these studies has been conducted in species belonging to what was formerly known as Colostethus (before Grant et al. 2006) and that now corresponds to several taxa with Most of these works showed that like C. aff. fraterdanieli, in species as Colostethus beebei (Bourne et al. 2001; now Anomaloglossus beebei; see Grant et al. 2006), Colostethus inguinalis (Wells 1980; now Colostethus panamensis; see Grant 2004), Colostethus subpunctatus (Fandiño et al. 1997; now Hyloxalus subpunctatus, see Grant et al. 2006), Mannophryne oblitterata (Dixon and Rivero-Blanco 1985) and Colostethus trinitatis (Cummins and Swan 1995, now Mannophryne trinitatis, see Grant et al. 2006) females reach sexual maturity at a larger size than males (table 2).
Age and size at sexual maturity is a critical component of life history studies because of their importance in determining the fitness (Roff 1992, Stearns 1992). Observed differences between sexes in MSSM, can relate to patterns of mating and parental effort (Howard 1981). For their reproductive role, females should produce and host the greatest possible number of eggs to maximize their fitness, which favors that females reach a larger size (Howard 1978, 1981, Woolbright 1983). Meanwhile, male fitness is not necessarily related to their body size depending on the mating system display for particular taxa (Howard 1981, Shine 1979).
We found that the ration of females and males in the population was very similar (1 female: 1.3 male), however the proportion of mature individuals differed between sexes, with a huge proportion of mature male (96%) compared with a median percentage of mature females (48%). These differences could be explained by variation in detectability of mature and immature males. Adult males usually display territorial and calling behaviors, which make them more conspicuous to the researchers. Unlike immature males that have more cryptic behavior because they are usually transients (Bailey et al. 2004, Mazerolle et al. 2007, de Solla et al. 2005).
Our results suggest sexual size dimorphism in Colostethus aff. fraterdanieli, where females are larger than males, as has been reported in the most of anuran species (Shine 1979) and particularly in other species of Colostethus (table 2, before Grant et al. 2006). Sexual dimorphism is the result of divergent selective pressures between sexes that promote morphological and behavioral differences (Howard 1981). However, regardless of the selection pressures and evolutionary forces that originated the differences in size between females and males, these seems to be the reflection of divergence in size at sexual maturity because in amphibians both sexes usually have the same body growth rates (Howard 1981). One the most known life history trade-offs is that between size and age at maturity that imply that to reduce time for growth, and decrease age at maturity might result in a decrease of adult body size (Roff 2000).
In this study, we found an evident dimorphism in throat coloration with males having a ''spotted'' pattern and females with ''immaculate'' throats. This kind of chromatic sexual dimorphism is relatively generalized between anurans and is related to a differential distribution of dermal chromatophore units between sexes with a higher number of melanophores per unit area in males (Buchanan 1994, Greenberg 1942). Differences in color saturation seems to be a secondary result of elevated testosterone levels in males (Buchanan 1994, Grant and Castro 1998, Greenberg 1942) therefore, immature males lack this character. Although not quantified here, we also evidenced that the third finger, as in many Colostethus species, is swollen in adult males (see figure 4).
The minimum size of sexual maturity is a critical parameter to understand the population dynamic of any species (Morrison and Hero 2003) and its knowledge is essential to make inferences about evolutionary history, population trends, and conservation status. Despite the importance of this parameter, most of studies in these kind of topics make inferences about it without a direct evaluation in the populations generating inaccuracy in their estimates and conclusions. For this reason, studies as this are valuable and constitute a baseline to other more specialized studies.
ACKNOWLEDGMENTS
This study was funded by ISAGEN and the Universidad de Antioquia under the Convenio Interinstitucional 46/4208. We thank S. Cuartas and the Grupo Herpetológico de Antioquia members for comments to an early draft and help during fieldwork.
REFERENCES
Bailey LL, Simons TR, Pollock KH. 2004. Spatial and temporal variation in detection probability of Plethodon salamanders using the robust capture-recapture design. Journal of Wildlife Management, 68: 14-24. [ Links ]
Bourne GR, Collins AC, Holder AM, McCarthy CL. 2001. Vocal communication and reproductive behavior of the frog Colostethus beebei in Guyana. Journal of Herpetology, 35: 272-281. [ Links ]
Buchanan BW. 1994. Sexual dimorphism in Hyla squirella: Chromatic and pattern variation between the sexes. Copeia, 1994: 797-802. [ Links ]
Crump M, Scott NJ. 2001. Relevamientos por encuentros visuales. In: Heyer R, Donnelly MD, McDiarmid LA, Hayek A, Foster MS. editors. Medición y monitoreo de la diversidad biológica. Métodos estandarizados para anfibios. Comodoro Rivadavia (Argentina): Editorial Universitaria de la Patagonia. p. 80-87. [ Links ]
Cummins CP, Swan MJ. 1995. Variation in reproductive characteristics of the stream frog Colostethus trinitatis on the island of Trinidad. Journal of Tropical Ecology, 11: 603-618. [ Links ]
Dixon JR, Rivero-Blanco C. 1985. A new dendrobatid frog (Colostethus) from Venezuela with notes on its natural history and that of related species. Journal of Herpetology, 19: 177-184. [ Links ]
Duellman W, Trueb L.1994. Biology of amphibians. Baltimore and London: The Johns Hopkins University Press. p. 670. [ Links ]
Fandiño MC, Lüddecke H, Amézquita A. 1997. Vocalisation and larval transportation of male Colostethus subpunctatus (Anura: Dendrobatidae). Amphibia-Reptilia, 18: 39-48. [ Links ]
Felgueiras-Napoli M. 2005. A new species allied to Hyla circumdata (Anura: Hylidae) from Serra da Mantiqueira, Southeastern Brazil. Herpetologica, 61: 63-69. [ Links ]
Grant T. 2004. On the identities of Colostethus inguinalis (Cope, 1868) and C. panamensis (Dunn, 1933), with comments on C. latinasus (Cope, 1863) (Anura: Dendrobatidae). American Museum Novitates, 3444: 1-24. [ Links ]
Grant T, Castro F. 1998. The cloud forest Colostethus (Anura, Dendrobatidae) of region of the Cordillera Occidental of Colombia. Journal of Herpetology, 32: 378-392. [ Links ]
Grant T, Frost DR, Caldwell JP, Gagliardo R, Haddad CF, Kok PJ, Means DB, Noonan BP, Schargel WE, Wheeler WC. 2006. Phylogenetic systematics of dart-poison frogs and their relatives (Amphibia: Athesphatanura: Dendrobatidae). Bulletin of the American Museum of Natural History, 299: 1-262. [ Links ]
Greenberg B. 1942. Some effects of testosterone in the sexual pigmentation and other sex characters of the cricket frog (Acris gryllus). Journal of Experimental Zoology, 91: 435-446. [ Links ]
Hammer O, Harper DAT, Ryan PD. 2001. Past: paleontological statistics software package for education and data analysis. Palaentologia Electronica, 4: 9. [ Links ]
Holdridge LR. 2000. Ecología basada en zonas de vida. Quinta reimpresión. San José (Costa Rica): Instituto Interamericano de Cooperación para la Agricultura (IICA). p. 216. [ Links ]
Howard RD. 1978. The influence of male-defended oviposition sites on early embryo mortality in bullfrogs. Ecology, 59: 789-798. [ Links ]
Howard RD. 1981. Sexual dimorphism in bullfrogs. Ecology, 62: 303-310. [ Links ]
Juncá FA. 1998. Reproductive biology of Colostethus stepheni and Colostethus marchesianus (Dendrobatidae), with the description of a new anuran mating behavior. Herpetologica, 54: 377-387. [ Links ]
Katsikaros K, Shine R. 1997. Sexual dimorphism in the tusked frog, Adelotus brevis (Anura: Myobatrachidae): the role of natural and sexual selection. Biological Journal of the Linnean Society, 60: 39-51. [ Links ]
Krzanowski WJ. 1979. Between groups comparison of principal components. Journal of the American Statistical Association, 74: 703-707. [ Links ]
Lima AP, Keller C. 2003. Reproductive characteristics of Colostethus marchesianus from its type locality in Amazonas, Brazil. Journal of Herpetology, 37: 754-757. [ Links ]
Lima AP, Caldwell JP, Biavati GM. 2002. Territorial and reproductive behavior of an Amazonian dendrobatid frog, Colostethus caeruleodactylus. Copeia, 2002: 44-51. [ Links ]
Lovich JE, Gibbons JW. 1992. A review of techniques for quantifying sexual size dimorphism. Growth, Development, & Aging, 56: 269-281. [ Links ]
Lovich JE, Gibbons JW, Agha M. 2014. Does the timing of attainment of maturity influence sexual size dimorphism and adult sex ratio in turtles? Biological Journal of the Linnean Society, 112: 142-149. [ Links ]
Luna L. 1968. Manual of Histologic Staining Methods of the Armed Forces Institute of Pathology. Third edition. New York (U. S. A.): Blakiston division, McGraw-Hill. p. 258. [ Links ]
Lynch JD. 1996. Eleutherodactylus erythropleura. Catalogue of American Amphibians and Reptiles, 623: 1-3. [ Links ]
Mazerolle MJ, Bailey LL, Kendall WL, Andrew-Royle J, Converse SJ, Nichols JD. 2007. Making great leaps forward: accounting for detectability in herpetological field studies. Journal of Herpetology, 41: 672-689. [ Links ]
Méndez-de la Cruz FR, Villagrán-Santa Cruz M, López-Ortíz ML, Hernández-Gallegos O. 2013. Reproductive cycle of a highelevation, oviparous lizard (Sceloporus spinosus: Reptilia: Phrynosomatidae). The Southwestern Naturalist, 58: 54-63. [ Links ]
Morrison C, Hero J-M. 2003. Geographic variation in life-history characteristics of amphibians: A review. Journal of Animal Ecology, 72: 270-279. [ Links ]
Morrison C, Hero J-M, Browning J. 2004. Altitudinal variation in the age at maturity, longevity and reproductive lifespan of anurans in subtropical Queensland. Herpetologica, 60: 34-44. [ Links ]
Prado A, Abdalla F, Silva P, Zina J. 2004. Late gametogenesis in Leptodactylus labyrinthicus (Amphibia, Anura, Leptodactylidae) and some ecological considerations. Brazilian Journal of Morphological Sciences, 21: 177-184. [ Links ]
Reyment RA, Blackith RE, Campell NA. 1984. Multivariate morphometrics. Second edition. Michigan (U. S. A.): Academic Press. p. 233. [ Links ]
Rojas B, Endler JA. 2013. Sexual dimorphism and intra-populational color pattern variation in the aposematic frog Dendrobates tinctorius. Evolutionary Ecology, 27: 739-753. [ Links ]
Roff DA. 1992. The evolution of life histories: Theory and analysis. New York, (U. S. A.): Chapman & Hall. p. 527. [ Links ]
Roff DA. 2000. Trade-offs between growth and reproduction: an analysis of the quantitative genetic evidence. Journal of Evolutionary Biology, 13: 434-445. [ Links ]
Rueda-Almonacid JV, Lynch JD. 1983. Una nueva especie de Eleutherodactylus para la cordillera oriental de Colombia. Lozania, 42: 1-6. [ Links ]
Shine R. 1979. Sexual selection and sexual dimorphism in the Amphibia. Copeia, 1979: 297-306. [ Links ]
Shine R. 1989. Ecological causes for the evolution of sexual dimorphism: A review of the evidence. The Quarterly Review of Biology, 64: 419-461. [ Links ]
de Solla SR, Shirose LJ, Fernie KJ, Barrett GC, Brousseau CS, Bishop, CA. 2005. Effect of sampling effort and species detectability on volunteer based anuran monitoring programs. Biological Conservation, 121: 585-594. [ Links ]
Stearns SC. 1992. The evolution of life histories. New York (U. S. A.): Oxford University Press. p. 246. [ Links ]
Wake MH, Dickie R. 1998. Oviduct structure, function, and reproductive modes in amphibians. The Journal of Experimental Zoology, 282: 477-506. [ Links ]
Watling J, Donnelly M. 2002. Seasonal patterns of reproduction and abundance of leaf litter frogs in a Central American rainforest. Journal of Zoology, 258: 269-276. [ Links ]
Wells KD. 1978. Territoriality in the green frog (Rana clamitans): vocalizations and agonistic behavior. Animal Behaviour, 26: 1051-1063. [ Links ]
Wells KD. 1980. Behavioral ecology and social organization of a dendrobatid frog (Colostethus inguinalis). Behavioral Ecology and Sociobiology, 6: 199-209. [ Links ]
Woolbright LL. 1983. Sexual selection and size dimorphism in anuran amphibian. The American Naturalist, 121: 110-119. [ Links ]
Yezerinac SM, Lougheed SC, Handford P. 1992. Measurement error and morphometric studies. Systematic Biology, 41: 471-482. [ Links ]
