










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActualidades Biológicas
Print version ISSN 0304-3584
Actu Biol vol.37 no.103 Medellín July/Dec. 2015
https://doi.org/10.17533/udea.acbi.v37n103a03
ARTÍCULOS DE INVESTIGACIÓN
doi: 10.17533/udea.acbi.v37n103a03
Diversidad de visitantes y aproximación al uso de nidos trampa para Xylocopa (Hymenoptera: Apidae) en una zona productora de pasifloras en Colombia
Diversity of visitors and approach to the use of trap nest for Xylocopa (Hymenoptera: Apidae) in a passion fruit production area in Colombia
Mario S. Pinilla-Gallego1 , Guiomar Nates-Parra2
1 Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, sede Bogotá. Bogotá, Colombia. simonpinilla@yahoo.com.
2 Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, sede Bogotá. Bogotá, Colombia. mgnatesp@unal.edu.co.
Recibido: agosto 2014; aceptado: marzo 2015.
RESUMEN
Las abejas del género Xylocopa son los polinizadores más eficientes de muchas plantas del género Passiflora, por esta razón, se ha propuesto el uso de "nidos trampa" para aumentar el número de visitas de estos polinizadores. El objetivo de este trabajo fue determinar la diversidad de visitantes florales y el nivel de polinización en cultivos de granadilla (Passiflora ligularis), gulupa (Passiflora edulis f. edulis) y maracuyá (Passiflora edulis f. flavicarpa), así como explorar la viabilidad de la cría de Xylocopa a través de nidos trampa. En cada cultivo se evaluó la frecuencia de visitas de Xylocopa y otros insectos durante dos periodos de floración, así como el porcentaje de formación de frutos y características de los frutos producidos. Se usaron nidos trampa de tubos de bambú y troncos de eucalipto. Los resultados muestran que en el cultivo de gulupa el porcentaje de formación de frutos es alto a pesar de la baja frecuencia de visitas de Xylocopa, mientras que en los cultivos de granadilla y maracuyá se podría incrementar el porcentaje de formación de frutos. Solo se logró que Xylocopa nidificara en los tubos de bambú del cultivo de maracuyá, pero con escaso éxito. Se discuten las posibles razones y se proponen lineamientos para los estudios de nidos trampa para Xylocopa en Colombia. Se realiza la primera descripción de los nidos de Xylocopa nasica en tubos de bambú.
Palabras clave: abejas carpinteras, abejas silvestres, nidos trampa, Passiflora, polinización, Xylocopa
ABSTRACT
The carpenter bees of the genus Xylocopa are some of the most efficient pollinators of many plants of the genus Passiflora and for this reason the use of "trap nest" has been proposed to increase the number of visits of these pollinators. The aim of this study was to establish the floral visitors diversity and pollination level in granadilla (Passiflora ligularis), purple passion fruit (Passiflora edulis f. edulis) and yellow passion fruit (Passiflora edulis f. flavicarpa) crops, as well as explore the viability of breeding Xylocopa using trap nest. In each crop the frequency of visits of Xylocopa and other insects in two blooms were assessed, as well as fruit set and fruit characteristics. Trap nests of bamboo canes and eucalyptus logs were used. Results showed that in the purple passion fruit crop the fruit set is high despite the low frequency of Xylocopa visits, while in the granadilla and yellow passion fruit crops the fruit set could be increased. We found that Xylocopa only nested in bamboo canes in the yellow passion fruit crop, but with low success. We discuss the possible reasons for this and propose guidelines for future Xylocopa trap nest studies in Colombia. The first description of Xylocopa nasica nests in bamboo canes is made.
Key words: carpenter bees, Passiflora, pollination, tramp nest, wild bees, Xylocopa
INTRODUCCIÓN
El género Passiflora (Passifloraceae) es uno de los más importantes en Colombia debido al gran número de especies cultivadas que posee. Algunas de las especies más importantes son la granadilla (Passiflora ligularis, Juss 1805), la gulupa (Passiflora edulis f. edulis, Sims 1818) y el maracuyá (Passiflora edulis f. flavicarpa, Degener 1932), siendo muy apetecidas en el mercado nacional e internacional (Miranda et al. 2009). Para el 2010, Colombia contaba con cerca de 4.300 ha destinadas al cultivo de granadilla, y una producción anual cercana a 50.000 t (Agronet 2012); para el 2012 la gulupa contaba con cerca de 600 ha cultivadas y una producción anual de 6.000 t, constituyendo el 40% de las exportaciones colombianas de frutas al mercado europeo y siendo la segunda fruta con mayor exportación en Colombia (Agronet 2012). La granadilla y la gulupa se cultivan principalmente en Boyacá, Antioquia, Risaralda y Tolima (Melgarejo 2012), mientras que el maracuyá, con cerca de 6.000 ha cultivadas principalmente en Santander y Valle del Cauca, cuenta con una producción anual de 80.000 t (Agronet 2012, Salinas 2010).
Las flores de estas plantas tiene antesis de un día, son perfectas y suelen ser auto-compatibles (p. ej., gulupa y granadilla), aunque algunas de ellas pueden presentar auto-incompatibilidad (p. ej., maracuyá), además de que la separación espacial de los órganos reproductivos de la flor hace que estas plantas requieran de vectores para su polinización, ya sea manual o llevada a cabo por polinizadores silvestres (Ángel et al. 2011, Bernal y Cabrera 2006, Da Silva et al. 1999, Valarezo et al. 2009). Debido a la morfología de la flor, los polinizadores más eficientes son las abejas grandes, especialmente las del género Xylocopa, ya que por su tamaño y comportamiento entran en contacto con las anteras y transfieren el polen a los estigmas de forma eficiente (Kishore et al. 2010, Koschnitzke y Sazima, 1997, Medina et al. 2011).
Las abejas del género Xylocopa son grandes, solitarias, comúnmente negras o de colores oscuros y construyen sus nidos en madera muerta, por lo que son conocidas como "abejas carpinteras" (Freitas y Oliveira 2003, González et al. 2009). Debido a su importancia en la polinización de los cultivos de Passiflora, se han propuesto varios modelos para la cría de Xylocopa. El primer modelo fue propuesto por Oliveira y Freitas (2003), quienes propusieron una "caja racional" que consiste en una colmena tipo Langstroth para Apis mellifera (Linneo 1758) modificada, en la que se introducen varias tablas en las cuales se espera que las hembras de Xylocopa nidifiquen. Dichos autores encontraron que en cada caja pueden nidificar de tres a cinco hembras. Pereira y Garófalo (2010) evaluaron un segundo modelo de tubos de bambú. Este tipo de nidos consiste en tubos de bambú cerrados en un extremo por el nodo de la planta, de 20-30 cm de largo y un diámetro de cerca de 2 cm, dentro de los cuales las hembras de Xylocopa construyen sus nidos. Un tercer modelo evaluado por Marchi y Melo (2010), consiste en troncos en los cuales se hacen pequeños agujeros de 2 cm de diámetro para inducir la nidificación de las hembras de Xylocopa. De estos modelos, los tubos de bambú parecen ser el modelo más exitoso. Adicionalmente, Freitas y Oliveira (2003) evaluaron el efecto de introducir cajas racionales para Xylocopa en cultivos de maracuyá en Brasil, con lo cual lograron aumentar el número de visitas a las flores y en consecuencia el nivel de polinización, y la producción del cultivo.
En Colombia se ha identificado a Xylocopa frontalis (Oliver 1789) y Xylocopa lachnea (Moure 1951) como principales polinizadores de la gulupa (Medina et al. 2011) y granadilla (Nates-Parra et al. 2010) en el municipio de Buenavista (Boyacá); Xylocopa sp. ha sido comunicada como polinizador del maracuyá en Valle del Cauca (Caicedo et al. 1993a). A pesar de esto, en Colombia solo existe un reporte sobre nidos trampa para Xylocopa en troncos de eucalipto (Caicedo et al. 1993b).
Debido a lo anterior, los objetivos de este estudio fueron: 1) determinar la diversidad de visitantes florales y la frecuencia de visitas de individuos de Xylocopa en una zona productora de pasifloras; 2) evaluar el nivel de polinización natural que se presenta en los cultivos y; 3) explorar la viabilidad de la cría de individuos de Xylocopa a través de nidos trampa de tubos de bambú y troncos de eucalipto.
MATERIALES Y MÉTODOS
Área de estudio. El estudio se llevó a cabo en el municipio de Buenavista (Boyacá), Colombia. Se seleccionó un cultivo de cada frutal: granadilla, gulupa y maracuyá. El cultivo de granadilla se encontraba en la vereda Sabaneta (5° 31' 03" N, 73° 56' 48" O) a 2.220 m. s. n. m., con un área de 1.500 m2. El cultivo de gulupa se encontraba en la vereda Santa Rosa (5° 30' 33" N, 73° 58' 26" O), a 1.800 m. s. n. m, con un área de 5.990 m2. El cultivo de maracuyá se ubicaba en la vereda La Herradura (5° 28' 32" N, 74° 00' 36" O) a 894 m. s. n. m, con un área de 1.930 m2.
Frecuencia de visitantes florales. Para medir la frecuencia de visitas de Xylocopa y otros visitantes en los cultivos se ajustó la metodología propuesta por Vaissiere et al. (2011). En los cultivos de granadilla y gulupa los muestreos se realizaron entre las 9 y 14 h, ya que estas son las horas de mayor actividad reportada para estas abejas en la zona de estudio (Nates-Parra et al. 2010, 2012). En el cultivo de maracuyá los muestreos se realizaron entre las 13 y 17 h, debido a que las flores abren a las 13 h (Amaya 2010). Se realizaron recorridos de 15 minutos cada hora, durante los cuales se registró el número de individuos de Xylocopa y otros visitantes que se encontraban en las flores. Se contó el número de flores abiertas presentes en las zonas donde se realizaron los recorridos. Se midió la temperatura (T) y la humedad relativa (HR) del aire durante los muestreos mediante un higrotermómetro. Se realizaron muestreos en dos periodos de floración, con cuatro días de muestreo en cada uno. En el cultivo de granadilla los muestreos se realizaron entre el 3-6 de julio del 2013 y 3-6 de diciembre del 2013. En el cultivo de gulupa los muestreos se realizaron entre el 11-14 de julio del 2013 y 23-26 de febrero del 2014. En el cultivo de maracuyá se realizó muestreos entre el 28-31 de julio del 2013; sin embargo, no fue posible realizar muestreos en una segunda floración debido a que en diciembre del 2013 el cultivo sufrió un ataque de arañuelas (= trips, Thysanoptera), y el propietario realizó una poda severa al cultivo, impidiendo que se presentara una segunda floración dentro del periodo de estudio.
Evaluación del nivel de polinización natural y características de los frutos. Para evaluar el nivel de polinización natural en los cultivos se marcaron 30 flores abiertas durante cada día de muestreo, para un total de 120 flores por cultivo en cada periodo de floración. Siete días después de marcadas las flores se determinó el porcentaje de formación de frutos (Ángel et al. 2011, Freitas y Oliveira 2003). Posteriormente, se recolectaron los frutos maduros formados a partir de las flores marcadas y se tomaron datos de tamaño (longitud y diámetro, en cm), peso fresco total (g), peso fresco de la pulpa (g) y número de semillas. Esta metodología se realizó en los dos periodos de floración mencionados anteriormente.
Nidos trampa. Se realizaron recorridos en zonas cercanas a los cultivo para buscar nidos naturales de Xylocopa, con el fin de instalar nidos trampa cerca de los nidos naturales, ya que esto puede aumentar la probabilidad de que Xylocopa use los nidos trampa (Caicedo et al. 1993b). Una vez determinados los lugares adecuados, se instalaron los nidos trampa en cada cultivo y zonas cercanas donde se encontraron nidos naturales. Los nidos trampa consistían en tubos de bambú de entre 15 a 25 cm de largo, con un diámetro de apertura de 1,3 a 2 cm, cerrados en un extremo por el nodo natural de la planta. También fueron usados troncos de Eucalyptus debido a que se ha registrado que estas abejas nidifican en dicho sustrato (Caicedo et al. 1993b, Camillo 2003); los troncos tenían aproximadamente 1 m de altura y 25 cm de diámetro, y su madera era blanda. En los troncos se perforaron agujeros de 1,5 cm de diámetro y 2 cm de profundo para intentar inducir la nidificación de Xylocopa (Da Silva et al. 2012, Oliveira 2008). Se instalaron 7 troncos y 200 tubos de bambú en cada zona de cultivo, para un total de 21 troncos y 600 tubos de bambú. Todos los tubos de bambú fueron instalados de tal manera que estuvieran protegidos de la lluvia (figura 1A), mientras que los troncos de eucalipto estuvieron expuestos a la lluvia. Los nidos trampa se instalaron entre el 25-30 de julio del 2013 (después de la primera floración evaluada de cada cultivo), y se les realizó seguimiento durante ocho meses.

De los tubos de bambú en los que nidificaron hembras de Xylocopa se tomaron medidas de largo, diámetro de apertura, grosor, número de celdas construidas, longitud de las celdas y grosor del opérculo en la base y en la parte central. En los casos en lo que se encontró polen dentro de los nidos, se tomó una muestra para realizar un análisis palinológico e identificar el origen botánico del polen hasta el nivel taxonómico más bajo posible.
Análisis de datos. Se usó la prueba de Shapiro-Wilk para determinar la distribución normal, y la prueba de Bartlett para evaluar la homogeneidad de varianzas. En caso de que los datos no tuvieran una distribución normal se transformaron por raíz cuadrada. Se aplicó la prueba paramétrica de análisis de varianza (ANAVA) de una vía para identificar diferencias significativas (p ≤ 0,05) en los datos de porcentaje de formación de frutos, longitud y diámetro de los frutos, peso total, peso de la pulpa y número de semillas. Para todos los análisis se utilizó el software de libre distribución R 2.12 (R Core Team 2014).
RESULTADOS
Frecuencia de visitantes florales. Granadilla. Las especies de abejas que visitaron las flores de granadilla fueron A. mellifera, Eulaema cingulata (Fabricius 1804), Thygater aethiops (Smith 1854) y X. lachnea. De estas, la más abundante fue A. mellifera, que durante la primera floración presentó una alta frecuencia de visitas ente las 9 y 12 h. Su frecuencia de visitas disminuye después de las 13 h. Durante la segunda floración las visitas de A. mellifera disminuyeron, aunque continuó siendo el visitante más frecuente y mantuvo el mismo patrón de visitas (figura 2).
La especie T. aethiops fue el segundo visitante más frecuente, y el número de visitas aumentó considerablemente en los muestreos de la segunda floración. Su actividad se mantiene constante entre las 9-12 h, después disminuye. La frecuencia de visitas por parte de X. lachnea fue baja, con valores mayores entre las 9 y 12 h, y un pico de actividad a las 9 h. Durante los muestreos de la segunda floración las visitas de X. lachnea aumentaron ligeramente a las 9 h. Las visitas de E. cingulata fueron muy esporádicas, tanto en la primera como en la segunda floración, y podría considerarse como un visitante ocasional en este cultivo (figura 2). Todas las abejas visitan las flores para obtener néctar.
Durante los muestreos de la primera floración la temperatura osciló entre 18 y 20 °C y la HR entre 58 y 68%. El número de flores abiertas en cada día de muestreo fue de 105 ± 22. Durante los muestreos de la segunda floración la temperatura osciló entre 16 y 22 °C y la HR entre 58 y 86%. El número de flores abiertas en cada día de muestreo fue de 83 ± 10.
Gulupa. Los visitantes registrados en este cultivo fueron A. mellifera, E. cingulata, Paratrigona sp., T. aethiops, Trigona truculenta (Almeida 1985) y X. lachnea. Durante la primera floración las visitas de abejas en general fueron escasas, siendo T. truculenta el visitante más frecuente, la cual presentó una mayor frecuencia de visitas entre las 9 y 11 h, después de lo cual el número de visitas disminuye rápidamente. Durante la segunda floración el número de visitas de T. truculenta aumentó considerablemente, siendo el segundo visitante más frecuente, y mantuvo el mismo patrón de visitas que en la primera floración (figura 3). Esta especie puede visitar las flores para recolectar polen o néctar, y en algunas ocasiones puede perforar la base de la corola de la flor para robar néctar.
La frecuencia de visitas de A. mellifera durante la primera floración fue baja, con su máximo número de visitas a las 9 h, disminuyendo cada hora a medida que avanzaba el día, pero durante la segunda floración el número de visitas aumentó considerablemente, llegando a ser el visitante más frecuente durante ese periodo, y mantuvo el mismo patrón de visitas que durante la primera floración (figura 3). Las visitas pueden ser para recolección de polen o néctar. La frecuencia de visitas por parte de X. lachnea fue baja y constante entre las 9 y 12 h, y disminuye a partir de las 13 h. Durante los muestreos de la segunda floración las visitas de X. lachnea fueron menos frecuentes y se registraron desde las 10 h, con un pico de actividad entre las 11-12 h, después de lo cual el número de visitas disminuye rápidamente (figura 3). Las visitas son para recolectar néctar.
Con respecto a la frecuencia de visitas de T. aethiops, esta fue baja a lo largo de todo el día durante la primera y segunda floración, y podría considerarse como un visitante ocasional en este cultivo. Las visitas de E. cingulata también fueron esporádicas, y durante la primera floración sólo se presentaron entre las 12 y 13 h. Durante la segunda floración las vistas de esta especie se observaron durante todo el día, pero continuaron siendo esporádicas. Por su parte, Paratrigona sp. solo se observó durante los muestreos de la segunda floración, y tuvo una frecuencia de visitas baja pero constante a lo largo de todo el día (figura 3). Las visitas de estas tres especies fueron para recolectar néctar
Durante los muestreos de la primera floración la temperatura osciló entre 18 y 24 °C y la HR entre 47 y 77%. El número de flores abiertas en cada día de muestreo fue de 116 ± 10. Durante los muestreos de la segunda floración la temperatura osciló entre 16,8 y 28,8 °C y la HR entre 61 y 89%. El número de flores abiertas en cada día de muestreo fue de 220 ± 7.
Maracuyá. Los visitantes registrados en este cultivos fueron Plebeia sp., Xylocopa nasica (Pérez 1901) y T. truculenta; esta última fue el visitante más frecuente en este cultivo, con un pico de actividad a las 14 h, llegando hasta 90 visitas/15 min, después de lo cual la actividad se reduce notoriamente (figura 4). La mayoría de las visitas de esta especie son para recolección de polen, y es común observarlas perforando la base de las flores para acceder a los nectarios (robo de néctar), incluso antes de que las flores abran.
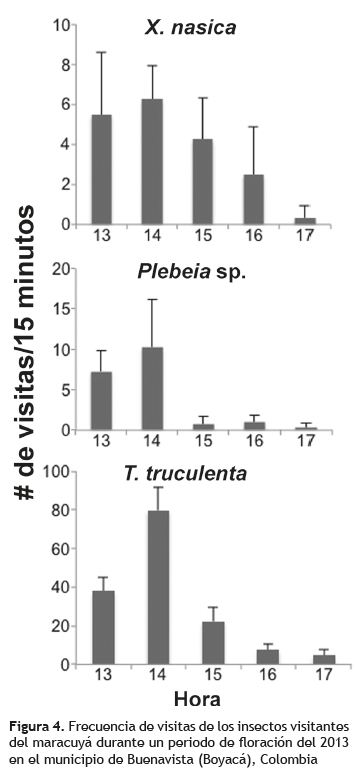
Las especies Plebeia sp. y T. truculenta presentaron el mismo patrón de visitas, con un alto número de visitas entre las 13 y 14 h, después de lo cual la frecuencia de visitas disminuye rápidamente; las visitas de Plebeia sp. son para recolección de polen, y no se observó robo de néctar. La frecuencia de visitas de X. nasica fue la más alta de Xylocopa en los tres cultivos de Passiflora, llegando hasta 9 visitas/15 min entre las 13 y 14 h; posteriormente la frecuencia de visitas empieza a disminuir (figura 4). Estas abejas visitan las flores para recolectar néctar.
Durante los muestreos de la floración evaluada la temperatura osciló entre 25 y 35 °C. No se registró la HR debido a una falla en el equipo de medición. El número de flores abiertas en cada día de muestreo fue de 195 ± 51.
Nivel de polinización natural y características de los frutos. En el cultivo de granadilla no hubo diferencias significativas (F = 0,709; df = 1; p > 0,05) entre el porcentaje de formación de frutos, la longitud y el diámetro de los frutos, peso de la pulpa y número de semillas entre la primera y segunda floración. La única variable que mostró diferencias significativas entre la primera y segunda floración fue el peso total de los frutos (F = 9,8; df = 1; p < 0,05) (tabla 1). En el cultivo de gulupa se encontraron diferencias significativas entre el peso total (F = 16,3; df = 1; p < 0,05) y el número de semillas (F = 1,35; df = 1; p < 0,05) de los frutos de la primera y segunda floración. No se encontraron diferencias significativas entre las demás variables evaluadas (F = 0,016; df = 1; p > 0,05) (tabla 1). En el caso del cultivo de maracuyá solo se obtuvieron datos de una floración, por lo que no fue posible comparar los datos de porcentaje de formación de frutos y características de los frutos (tabla 1), ni medir el efecto que habrían tenido los nidos trampa en este cultivo.
Nidos trampa. Dentro de la finca del cultivo de granadilla se encontraron tres nidos naturales activos de Xylocopa en troncos de eucalipto utilizados en cercas. En zonas cercanas se encontraron cuatro nidos activos en troncos de aliso (Alnus sp.) muerto, para un total de siete nidos naturales, cerca de los cuales se ubicaron los nidos trampa. Ninguno de los nidos trampa de bambú o troncos de eucalipto fue usado por estas abejas para nidificar (porcentaje de ocupación 0%).
En el cultivo de gulupa se encontró un nido activo de Xylocopa en una tabla que hacia parte del techo de una caseta donde se guardaba herramienta. En zonas cercanas al cultivo se encontraron cuatro nidos activos en las vigas de soporte del techo de una casa, para un total de cinco nidos activos; en este caso la madera no pudo ser identificada. Ninguno de los nidos trampa de bambú o troncos de eucalipto fue usado por estas abejas para nidificar (porcentaje de ocupación 0%).
Dentro de la finca del cultivo de maracuyá se encontraron dos nidos naturales de Xylocopa en un tronco caído de aguacate (Persea sp.), y este fue el único cultivo donde Xylocopa nidificó en los tubos de bambú. Se logró obtener seis nidos de Xylocopa nasica (porcentaje de ocupación de 3%) (figura 1B). Los tubos en los que nidificaron las abejas tenían una longitud de 19,4 ± 3,3 cm, diámetro de apertura de 1,6 ± 0,1 cm y grosor de las paredes del tubo de 4 ± 1 mm (n = 6). Las hembras habían raspado la parte interna de los tubos en los lugares donde construyeron las celdas, de tal manera que el grosor de los tubos se vio reducido en esas partes. En promedio los nidos tenían 3 ± 2 celdas, la longitud de las celdas fue de 2,1 ± 0,2 cm, el grosor en la base del opérculo fue de 5 ± 2 mm y el grosor en el centro del opérculo fue de 3 ± 2 mm (n=15) (figura 1C). De los nidos recolectados se obtuvieron cinco muestras de polen, dentro de las cuales se encontró polen de plantas de las familias Fabaceae, Sapindaceae y Solanaceae, siendo el polen de Fabaceae el más abundante. No se obtuvieron nidos en los troncos de eucalipto.
DISCUSIÓN
Frecuencia de visitantes florales y características de los frutos. Franco et al. (2007) comunicaron siete especies de abejas visitantes de cultivos de granadilla en altitud similar al evaluado en este trabajo (2.100 m), y al igual que en este caso, la frecuencia de visitas de Xylocopa fue baja, pero el porcentaje de formación de frutos fue alto (84%). En ambos estudios, el nivel de polinización elevado puede deberse al efecto combinado de las visitas de varias especies de abejas o al alto número de visitas de A. mellifera, ya que son abejas de tamaño medio y pueden estar transfiriendo polen al estigma de las flores, y dado que la granadilla es una planta auto-compatible la probabilidad de que A. mellifera contribuya de manera significativa a su polinización es mayor (Nates-Parra et al. 2010). Aun así se debe tener en cuenta que el nivel de polinización en el cultivo evaluado se encuentra entre el 61 y 77%, por lo que se podría esperar aumentar dicho porcentaje, y en este caso lo más viable seria incrementar las visitas de A. mellifera aumentando el número de colmenas cerca al cultivo durante los periodos de floración.
El peso total de los frutos de granadilla en la segunda floración fue significativamente mayor que los de la primera floración; sin embargo, es poco probable que este aumento en el peso total se deba a una mejora en la polinización, ya que el número de semillas y el peso de la pulpa no aumentaron, y estas son las variables que están directamente relacionadas con la polinización (Dafni et al. 2005). Por esta razón, es poco probable que el aumento en las visitas de T. aethiops durante la segunda floración sean la causa del aumento en el peso total de los frutos, ya que las visitas únicas de Thygater generan solamente un 5,3% de formación de frutos (Nates-Parra et al. 2010). Es más probable que estas diferencias se deban a cambios en variables ambientales (precipitación, temperatura) o de manejo de la fertilización del cultivo.
Medina et al. (2011) registraron 12 especies de abejas visitantes en un cultivo de gulupa a 1.657 m de altitud en la misma zona en la que se realizaron los muestreos de este estudio. En contraste, en nuestros muestreos solo se observaron cinco especies de abejas visitando las flores. Estas diferencias podrían deberse a la pequeña pero importante diferencia en altura de los cultivos (cerca de 200 m), que puede restringir la distribución de muchas especies de abejas. Dichos autores además reportan que el 33% de las visitas a las flores son por parte de Xylocopa (Medina et al. 2011), lo que muestra que la frecuencia de visitas de Xylocopa observada en este estudio fue baja en las dos floraciones evaluadas. Esto puede deberse a diferencias en el manejo de agroquímicos en los cultivos y a que cerca al cultivo evaluado se encontraba poca vegetación natural, lo que afecta negativamente las poblaciones de abejas y como consecuencia la frecuencia de visitas de Xylocopa en los cultivos (Freitas y Oliveira 2003).
A pesar de esto, el porcentaje de formación de frutos en ambas floraciones fue superior al 90%, lo cual es un valor alto, que probablemente se deba al efecto sinérgico de las visitas de A. mellifera y T. truculenta, que aunque en ocasiones pueden ser ladrones de polen y néctar, también pueden hacer contacto con las estructuras reproductivas de la flor (Nates-Parra et al. 2012), y dado que la flor de la gulupa es auto-compatible (Ángel et al. 2011), permitiría un nivel de polinización alto. El aumento en las visitas de A. mellifera y T. truculenta durante la segunda floración probablemente se deba al aumento en el número de flores abiertas durante los muestreos de la segunda floración, lo que podría atraer más abejas. El aumento en la frecuencia de estos visitantes explica el aumento en el número de semillas de los frutos y en consecuencia el peso total de los frutos, ya que estas son variables directamente relacionadas con la polinización (Dafni et al. 2005). Estos resultados muestran que en la gulupa es posible obtener un porcentaje de formación de frutos alto gracias a las visitas de otras abejas cuando las visitas de Xylocopa son escasas. Por esta razón, en este cultivo no sería necesario introducir nidos trampa para Xylocopa, además de que es difícil que estas abejas los usen en esta zona.
En el cultivo de maracuyá se observó la frecuencia de visitas más alta por parte de Xylocopa de los tres cultivos de Passiflora evaluados. Esto puede deberse a que esta zona es la que cuenta con mayor área de vegetación natural de los tres cultivos. Adicionalmente este cultivo se encontraba a menor altitud, donde la abundancia de Xylocopa es mayor que en zonas más altas (como los cultivos de granadilla y gulupa), lo que explicaría que la población de Xylocopa sea mayor que en las demás zonas.
A pesar de la alta frecuencia de visitas de X. nasica, el nivel de polinización natural fue el más bajo de los tres cultivos evaluados. Dado que las flores abren cerca del mediodía, muchas abejas ya se encuentran activas, y en este caso las visitas de Plebeia sp. y T. truculenta son exclusivamente para recolectar polen. Muchos autores han informado que cuando el robo de polen por parte de A. mellifera o Meliponini es alto en cultivos de maracuyá, el polen se remueve rápidamente y no llega a estar disponible para que abejas grandes como Xylocopa lo transporten entre flores para una efectiva polinización (Camillo 2003, Junqueira et al. 2013, Oliveira 2008). Esta podría ser la razón por la qué a pesar de la alta frecuencia de visitas de Xylocopa el porcentaje de formación de frutos fue bajo. Adicionalmente, la flor del maracuyá es auto-incompatible (Valarezo et al. 2009), por lo que es menos probable que abejas pequeñas como T. truculenta lleguen a aportar a la polinización de esta planta. Como solución al problema de robo de polen se ha propuesto el uso de trampas florales, que atraen las abejas pequeñas, y pueden reducir hasta en 83% las visitas de A. mellfera en cultivos de maracuyá (Pelaez, 2004), y debido a que en este caso los tubos de bambú si fueron usador por Xylocopa, podrían ser una opción para aumentar la población de polinizadores en los cultivos. Silveira et al. (2012) además proponen realizar polinización manual en solo un estigma, lo que sumado a una buena población de polinizadores puede mejorar la producción sin aumentar demasiado los costos de producción.
Nidos trampa. En países como Brasil los estudios sobre nidos trampa para Xylocopa han avanzado mucho y cuentan con información detallada sobre hábitos de nidificación y biología de especies como X. frontalis y Xylocopa grisescens (Lepeletier 1841). En Colombia este es el primer registro de uso de nidos trampa por X. nasica, así como la primera descripción de los tubos de bambú usados por esta especie para nidificar. El diámetro de los tubos de bambú usados por X. nasica (1,6 ± 0,1 cm) es menor que el diámetro de lo tubos usados por X. frontalis y X. grisescens (1,9 ± 0,3 cm) (Pereira y Garófalo 2010), y esto puede deberse a que X. nasica es de menor tamaño, por lo que puede preferir tubos de menor diámetro. Por otro lado, la longitud de los tubos usados por X. nasica fue similar al registrado para X. frontalis y X. grisescens.
Diferentes autores señalan diferentes porcentajes de uso de tubos de bambú por parte de Xylocopa, que pueden estar desde el 8,5% (Pereira y Garófalo 2010), 12% (Marchi y Melo 2010), 25% (Camillo 2003), hasta un 45% (Junqueira et al. 2012), y se ha registrado nidificación en nidos trampa de troncos de Eucalyptus sp., Tibouchina sp. (Melastomataceae) y Ficus sp. (Moraceae) (Camillo 2003, Marchi y Melo 2010). En este estudio, de los nidos trampa instalados (tubos de bambú y troncos de eucalipto), sólo se logró que Xylocopa nidificaran en los tubos de bambú del cultivo de maracuyá, y aun así el porcentaje de uso de los tubos fue de tan solo 3%. El poco éxito obtenido con los nidos trampa probablemente se deba al bajo nivel de conocimiento que tenemos sobre la biología de estas abejas en Colombia. Algunas de los factores más importantes a tener en cuenta son los siguientes:
Algunas especies de Xylocopa presentan ciclos de reproducción semestrales o anuales, como es el caso de X. frontalis en Morretes (Brasil), en donde las hembras fundan sus nidos entre los meses de octubre y diciembre (Marchi y Melo 2010). Debido a que en este estudio el seguimiento de los nidos trampa fue solo por ocho meses, es posible que el bajo porcentaje de uso se deba a que las hembras no se encontraran en sus fase reproductiva. Es muy poco lo que se conoce sobre la biología reproductiva de las especies nativas, por lo que se requieren estudios por periodos superiores a un año, que permitan establecer si las especies de una zona presentan ciclos estacionales de reproducción, y la época en que esto sucede, para así poder determinar los momentos adecuados para instalar nidos trampa que favorezcan la reproducción de estas especies.
En los cultivos de granadilla y gulupa las visitas de Xylocopa fueron escasas, esto indica que su población en los periodos evaluados era baja, lo que hace menos probable que alguna hembra llegue a utilizar los nidos trampa. Junqueira et al. (2013) obtuvieron resultados similares al introducir tubos de bambú en cultivos de maracuyá donde la población inicial de Xylocopa era baja, mientras que al introducir nidos trampa en cultivos con mayor población inicial de Xylocopa lograron que los tubos de bambú fueran usados por las abejas. Este parece ser el caso del cultivo de maracuyá de este trabajo, donde el número de visitas de Xylocopa en las flores fue el más alto de los tres cultivos y fue el único sitio donde Xylocopa usó los nidos trampa. Estos resultados sugieren que en zonas donde Xylocopa es escasa, el uso de nidos trampa requeriría mayor tiempo para poder aumentar notoriamente su población, por lo que sería más viable instalar los nidos trampa en una zona con una alta densidad de Xylocopa, y una vez los nidos estén ocupados, trasladarlos a la zona deseada, ya sea para polinización de cultivos o para recuperar poblaciones en peligro.
También se debe tener en cuenta que hay especies que construyen sus nidos naturales de forma recta (p. ej., Xylocopa pubescens), por lo que pueden verse más atraídas hacia los tubos de bambú, mientras que las especies que construyen galerías ramificadas (p. ej., Xylocopa sulcatipes) pueden verse menos atraídas, ya que los tubos de bambú no les permiten dicha arquitectura en los nidos (Velthuis et al. 1984), siendo necesario en este caso usar troncos como nidos trampa. Para las especies nativas es muy poco lo que se sabe sobre su biología y arquitectura de los nidos: Fernández y Nates-Parra (1985) estudiaron la localización y contenido de algunos nidos en el parque de La Macarena (Meta); Caicedo et al. (1993b) encontraron que Xylocopa sp. presenta un 22% de uso de troncos de eucalipto en Zarzal (Valle del Cauca), y González et al. (2009) describieron nidos naturales de X. lachnea. Por lo anterior, es importante llevar a cabo estudios que establezcan que especies son más aptas para nidificar con nidos trampa, así como determinar las características de los tubos de bambú o las especies vegetales preferidos para nidificar por las especies de cada zona, para de esta manera aumentar la probabilidad de que usen los nidos trampa, como lo informaron Junqueira et al. (2012), quienes en un primer intento obtuvieron 21% de uso de tubos de bambú, pero luego de determinar las características adecuadas de los tubos obtuvieron un porcentaje de uso del 45%.
CONCLUSIONES
Las especies de visitantes florales que se pueden encontrar en cultivos de Passiflora varían incluso en zonas cercas, y en este caso, dependiendo de la biología reproductiva de la planta y de las especies de visitantes, puede presentarse o no un bajo nivel de polinización. En el cultivo de gulupa se presenta alto nivel de polinización gracias a las frecuentes visitas de abejas de tamaño mediano, y no se requeriría aumentarlo. En el cultivo de la granadilla el porcentaje de polinización es intermedio y podría aumentarse con el manejo de colmenas de A. mellifera, ya que en esta zona Xylocopa no usó los nidos trampa. Por otro lado, el bajo nivel de polinización en el cultivo de maracuyá probablemente se debe al frecuente robo de polen por parte de abejas pequeñas, y en este caso el uso de tubos de bambú para la cría de Xylocopa si sería una alternativa para incrementar el nivel de polinización en el cultivo. Con este trabajo se aporta información sobre los hábitos de nidificación de una especie nativa y además este es el primer reporte de la nidificación de Xylocopa nasica en nidos trampa, con lo que se contribuye con datos para futuros estudios sobre esta especie. De igual manera, también es importante resaltar que antes de pensar en usar nidos trampa de Xylocopa para aumentar su densidad en un cultivo, es importante determinar la época y duración de los ciclos reproductivos de las especies presentes en la región y las características de los sustratos usados para nidificar naturalmente, para de esta forma diseñar los nidos trampa apropiados y así incrementar la probabilidad de que sean usados por estas abejas.
AGRADECIMIENTOS
A los productores Rodrigo Ballesteros, José García y Arnoldo Forero. Al Departamento Administrativo de Ciencia, Tecnología e Innovación (COLCIENCIAS) y a la Universidad Nacional de Colombia por la financiación del proyecto dentro del programa "Jóvenes investigadores e innovadores 2012". A los evaluadores anónimos por sus aportes.
REFERENCIAS
Agronet [internet]. 2012. Ministerio de Agricultura. Fecha de acceso: el 27 de abril del 2014. Disponible en: http://www.agronet.gov.co/www/htm3b/ReportesAjax/parametros/reporte16_2011. aspx?cod=16. [ Links ]
Amaya JE. 2010. Cultivo de maracuyá (Passiflora edulis Sim f. flavicarpa Deg). Trujillo (Perú): Gerencia Regional Agraria La Libertad. p. 24. [ Links ]
Ángel C, Nates-Parra G, Ospina R, Melo CD. 2011. Biología floral y reproductiva de la gulupa (Passiflora edulis Sim edulis). Caldasia, 33 (2): 433-451. [ Links ]
Bernal JA, Cabrera CA. 2006. Manual técnico del cultivo de granadilla (Passiflora ligularis Juss) en el departamento del Huila. Neiva (Colombia): Cadena Productiva Frutícola. p. 34. [ Links ]
Caicedo G, Vargas H, Gaviria J. 1993a. Evaluación de Xylocopa spp. (Hymenoptera: Anthophoridae) como polinizadores en el cultivo del maracuyá (Passiflora edulis var. flavicarpa Degener). Revista Colombiana de Entomología, 19 (3): 107-110. [ Links ]
Caicedo G, Vargas H, Gaviria J. 1993b. Estudio del modelo natural de asentamiento de Xylocopa (Hymenoptera: Anthophoridae) para la adaptación de refugios en el cultivo de maracuyá (Passiflora edulis var. flavicarpa Degener). Revista Colombiana de Entomología, 19 (2): 72-78. [ Links ]
Camillo E. 2003. Polinização do maracujá. Ribeirão Preto (Brasil): Editorial Holos. p. 44. [ Links ]
Dafni A, Kevan GP, Husband CB. 2005. Practical pollination biology. Ontario (U. S. A): Enviroquest Ltd. p. 315. [ Links ]
Da Silva CI, Oliveira PE, Garófalo CA. 2012. Manejo e conservação de polinizadores do maracujazeiro amarelo (Passiflora edulis f. flavicarpa). En: Fátima M, editor. 2012. III semana dos polinizadores, Palestras e resumos. Petrolina (Brasil): Editorial Márcia de Fátima Ribeiro. p. 162- 178. [ Links ]
Da Silva M, Bruckner CH, Picaço M, Molina-Rugama AJ. 1999. Número floral, clima, densidad poblacional de Xylocopa spp. (Hymenoptera: Anthophoridae) y polinización de maracuyá (Passiflora edulis f. flavicarpa). Revista de Biología Tropical, 47 (4): 711-718. [ Links ]
Fernández F, Nates-Parra G. 1985. Hábitos de nidificación en abejas carpinteras del género Xylocopa (Hymenoptera: Anthophoridae). Revista Colombiana de Entomología, 11(2): 36-41. [ Links ]
Franco Y, Alzate F, Peláez JM. 2007. Factores ambientales incidentes en la población de Xylocopa y su efecto en el cultivo de granadilla en tres veredas del municipio Guarne (Colombia). Revista Universidad Católica de Oriente, 24: 73-86. [ Links ]
Freitas BM. Oliveira JH. 2003. Ninhos racionais para mamangava (Xylocopa frontalis) na polinização do maracujá-amarelo (Passiflora edulis). Ciência Rural, 33 (6): 1135-1119. [ Links ]
González VH, González MM, Cuellas Y. 2009. Notas biológicas y taxonómicas sobre los abejorros del maracuyá del género Xylocopa (Hymenoptera: Apidae, Xylocopini) en Colombia. Acta Biológica Colombiana, 14 (2): 31-40. [ Links ]
Junqueira CN, Hogendoorn K, Augusto C. 2012. The use of trap-nests to manage carpenter bees (Hymenoptera: Apidae: Xylocopini), pollinators of passion fruit (Passifloraceae: Passiflora edulis f. flavicarpa). Annals of the Entomological Society of America, (105 (6): 883-889. [ Links ]
Junqueira CN, Yamamoto M, Oliveira PE, Hogendoorn K, Augusto SC. 2013. Nest management increases pollinators density in passion fruit orchard. Apidologie, 44 (6): 729-737. [ Links ]
Kishore K, Pathak KA, Shukla K, Bharali A. 2010. Studies on floral biology of passion fruit (Passiflora spp.). Pakistan Journal of Botany, 42 (1): 21-29. [ Links ]
Koschnitzke C, Sazima M. 1997. Biologia floral de cinco espécies de Passiflora L. (Passifloraceae) em mata semidecídua. Revista Brasileira de Botânica, 20 (2): 119-126. [ Links ]
Marchi P, Melo GR. 2010. Biologia De Nidificação De Xylocopa (Neoxylocopa) Frontalis (Olivier) (Hymenoptera, Apidae, Xylocopini) em Morretes, Paraná. Oecologia Australis, 14 (1): 210-231. [ Links ]
Medina J, Ospina R, Nates-Parra G. 2011. Efectos de la variación altitudinal sobre la polinización en cultivos de gulupa (Passiflora edulis f. edulis). Acta Biológica Colombiana, 17 (2): 381-396. [ Links ]
Melgarejo LM, editores. 2012. Ecofisiología del cultivo de gulupa (Passiflora edulis Sim). Bogotá (Colombia): Universidad Nacional de Colombia. p. 144. [ Links ]
Miranda D, Fischer G, Carranza C, Magnitskiy S, Casierra F, Piedrahíta W, Flórez LE. 2009. Cultivo, poscosecha y comercialización de las pasifloras en Colombia: maracuyá, granadilla, gulupa y curuba. Bogotá (Colombia): Sociedad colombiana de ciencias hortícolas. p. 360. [ Links ]
Nates-Parra G, Amaya M, Ospina R. 2010. Manejo de la polinización con abejas en cultivos de gulupa y granadilla. Bogotá (Colombia): Universidad Nacional de Colombia, Facultad de Ciencias, Departamento de Biología. Informe técnico final N.° DIB- 8003251. [ Links ]
Nates-Parra G, Amaya M, Ospina R, Ángel C, Medina J. 2012. Biología floral, reproductiva, polinización y polarizadores en gulupa (Passiflora edulis var. edulis). 115-121. En: Melgarejo L, editor. 2012. Ecofisiología del cultivo de gulupa (Passiflora edulis Sim). Bogotá (Colombia): Universidad Nacional de Colombia. p. 135-144. [ Links ]
Oliveira PE. 2008. Sustainable Management of pollinators for passionfruit (Passiflora edulis f. flavicarpa Deg., Passifloraceae) production in central Brazil. En: Benfica CA, editor. 2008. Pollinators management in Brazil. Brasilia (Brasil): Ministry of the environment. p. 30-33. [ Links ]
Oliveira JH, Freitas BM. 2003. Colonização e biologia reprodutiva de mamangavas (Xylocopa frontalis) em um modelo de ninho racional. Ciencia Rural, 33 (4): 693-697. [ Links ]
Pelaez JM. 2004. Trampas florales para el control de Apis mellifera en cultivos de maracuyá (Passiflora edulis var. flavicarpa Degener). II Encuentro Colombiano sobre abejas silvestres, Bogotá, Colombia. [ Links ]
Pereira M, Garófalo C. 2010. Biologia da nidificação de Xylocopa frontalis e Xylocopa grisescens (Hymenoptera, Apidae, Xylocopini) em ninhos-armadilha. Oecologia Australis, 14 (1), 193-209. [ Links ]
R Core Team, A Language and Environment for Statistical [Internet]. 2014. R Foundation for Statistical Computing. Fecha de acceso: 10 de noviembre del 2014. Disponible en http://www.r-project.org [ Links ]
Salinas H. 2010. Guía técnica para el cultivo de "maracuyá amarillo". Roldanillo (Colombia): Instituto de Educación Técnica Profesional de Roldanillo Valle. p. 44. [ Links ]
Silveira M, Abot A, Nascimento J, Rodrigues E, Rodriguez S, Puker A. 2012. Is manual pollination of yellow passion fruit completely dispensable? Scientia Horticulturae, 146 (1): 99-103. [ Links ]
Vaissiere B, Freitas BM, Gemmill-Herren,B. 2011. Protocol to detect and assess pollination deficits in crops: a handbook for its use. Roma (Italia): Food and Agriculture Organization of the United Nations (FAO). p. 72. [ Links ]
Valarezo A, Cañarte E, Valarezo O, Zambrano O. 2009. Manejo del cultivo de maracuyá (Passiflora edulis f. flavicarpa Deg) en el litoral Ecuatoriano. Portoviejo (Ecuador): INIAP. p. 12. [ Links ]
Velthuis HH, Wolf Y, Gerling D.1984. Provisioning and preparation of the brood cells in two carpenter bees, Xylocopa sulcatipes MAA and Xylocopa pubescens spinal (Hymenoptera: Anthophoridae). Israel Journal of Entomology, 18 (1): 39-51. [ Links ]
