










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActualidades Biológicas
Print version ISSN 0304-3584
Actu Biol vol.38 no.105 Medellín July/Dec. 2016
https://doi.org/10.17533/udea.acbi.v37n105a02
RESEARCH PAPERS
doi: 10.17533/udea.acbi.v37n105a02
Divergent drivers of tree community composition in lowland and highland forests of the northern tropical Andes, Colombia
Divergencia en los determinantes de la composición de comunidades arbóreas entre bosques de tierras bajas y tierras altas del noroeste de los Andes tropicales, Colombia
Álvaro Idárraga1,4, Álvaro J. Duque-Montoya2,5, Kenneth Feeley3,6
1 Instituto de Biología, Universidad de Antioquia. A. A. 1126. Medellín (Antioquia), Colombia. alvaro.idarraga@udea.edu.co - alvaro.idarraga@gmail.com
2 Departamento de Ciencias Forestales, Universidad Nacional de Colombia Sede Medellín. Medellín (Antioquia), Colombia. ajduque@unal.edu.co - ajduque09@gmail.com
3 International Center for Tropical Botany, Department of Biological Sciences, Florida International University & Fairchild Tropical Botanic Garden. Coral Gables (FL). USA. kfeeley@fiu.edu.
Received: February 2016; accepted: May 2016 (Recibido: febrero 2016; aceptado: mayo 2016).
Abstract
Identifying the determinants of tree species composition among contrasting altitudinal tropical forest types will improve our understanding of the main factors and processes controlling tree species' distributions and patterns of diversity. The study area is located in the northwest region of Colombia. We used Redundancy Analysis (RDA) and a semi-log distance-decay model to analyze the influence of climate, soil fertility, and spatial processes, such as dispersal limitation, on community composition. There were 3 distinct forest types: one in the highlands and two in the lowlands. In the lowlands, both the RDA and the semi-log linear models identified calcium concentrations as the most important explanatory variable for species composition (R2adj = 0.24; p = 0.001 and RMantel = 0.68; p = 0.006, respectively). In the highlands, the RDA only identified longitude as being a significant explanatory variable (R2adj = 0.24; p = 0.001), primarily determined by the biogeographical location of forests on either the West or the Central Cordillera of the Andes. In these neotropical forests, the factors determining tree species composition between lowlands and highlands varied in both extent and nature. Overall, our findings do not support the idea of a higher habitat-plant specialization in highlands than in lowlands due to greater geomorphologic and edaphic variation. Since tropical Andean ecosystems, and in particular highland forests, have received less attention than their lowland counterparts, our findings help to expand the current knowledge of factors and processes determining community composition of tropical montane forests.
Key words: Colombia, elevational gradient, environmental filtering, montane forests, spatial analysis.
Resumen
Identificar los determinantes de la composición de especies arbóreas entre tipos de bosques tropicales altitudinalmente contrastantes incrementarán nuestro conocimiento de los principales factores y procesos que controlan los patrones de diversidad y distribución de las especies. El área de estudio se localiza en la región noroeste de Colombia, principalmente en el departamento de Antioquia. Se empleó el análisis de redundancia (RDA) y un modelo semi-logarítmico linealizado para analizar la influencia del clima, fertilidad del suelo, y procesos espaciales, tales como la limitación en dispersión, sobre la composición de la comunidad arbórea. Se encontraron tres tipos de bosques: uno en tierras altas y dos en tierras bajas. En las tierras bajas, ambos modelos, el RDA y el modelo semi-logarítmico linealizado, identificaron las concentraciones de calcio como la variable que mejor explica la composición de especies (R2adj = 0,24; p = 0,001 y RMantel = 0,68; p = 0,006, respectivamente). En las tierras altas, el RDA solo identificó la longitud como una variable explicativa significativa (R2adj = 0,24; p = 0,001), determinada principalmente por la ubicación biogeográfica de los bosques en la cordillera Occidental o Central de los Andes. En estos bosques neotropicales, los factores que determinan la composición de especies de árboles entre tierras bajas y altas varió en magnitud y naturaleza. En general, nuestros resultados no apoyan la idea que existe una mayor especialización de hábitat en tierras altas que en tierras bajas debido a mayor variación geomorfológica y edáfica. Los ecosistemas tropicales andinos y en particular los bosques de tierras altas, han recibido menos atención que sus contrapartes de tierras bajas; nuestros hallazgos ayudan a ampliar el conocimiento actual de los factores y procesos que determinan la composición de las comunidades de los bosques montanos tropicales.
Palabras claves: análisis ambientales, bosques de montaña, Colombia, filtros ambientales, gradiente de elevación.
INTRODUCTION
Changes in tree species composition along spatial or environmental gradients can be driven by either environmental filtering, which is associated with abiotic factors such as climate, topography and soil characteristics (Aiba and Kitayama 1999, Gentry 1988, Hemp 2005, Vásquez and Givnish 1998), or by spatially-driven stochastic processes, such as dispersal limitation (Condit et al. 2002, Hubbell 2001, Vormisto et al. 2004). Climate is expected to be the most important factor controlling species distributions at the regional scale (Clinebell et al. 1995, Engelbrecht et al. 2007), while topography and soils are expected to play more important roles at the meso and local scales (Clark et al. 1999, Condit et al. 2013, Duque et al. 2002, John et al. 2007). Regarding stochastic factors, in forested areas it is expected that dispersal limitation will not play a significant role in determining community composition at geographic distances larger than approximately 50-100 km (Chain-Guadarrama et al. 2012, Condit et al. 2002). Dispersal limitation has been proposed as the main mechanism determining species neutrality due to its capability to overcome differential competitive advantages associated with species life history (Hubbell 2001). However, the presence of major physical barriers (e.g., the Andean mountain ridges) may cause habitat isolation and promotes dispersal limitation, thereby determining species distributions at large spatial scales as well (Nekola and White 1999).
In the northern part of the Andes, lowland valleys are isolated by of the presence of mountain ranges that reach up to 4000 m.a.s.l. (meters above sea level). Therefore, under the assumption of dispersal limitation as key determinant of community composition, forests located on similar ranks of elevation (e.g., lowlands or highlands) within the same valley or catchment may demonstrate higher floristic similarity when compared to those located on the opposite sides of ridges (Chain-Guadarrama et al. 2012, Condit et al. 2002, Nekola and White 1999). Otherwise, environmental filters imposed by either climatic variability or soil fertility (Condit et al. 2013, Engelbrecht et al. 2007, Garibaldi et al. 2014, Jones et al. 2011), are expected to be the primary determinants of species distributions.
Some authors (e.g., Gentry 1982, Hemp 2005, Jones et al. 2011) have proposed that the turnover in plant species composition between highland ecosystems of the same elevation is greater than turnover between lowland ecosystems because of lack of habitat connectivity as well as higher habitat specialization associated with a greater climatic, edaphic, and topographic variation. Overall, habitat similarity in the high mountain forests is expected to decrease rapidly with geographic distance and similar habitats can be highly fragmented. This isolation poses limits to dispersal (Garibaldi et al. 2014) and could also enhance differences in habitat response or habitat-specialization among taxa (Nekola and White 1999). However, extreme environmental conditions and highly variable climates could promote decreased habitat specialization and broader species ranges due to greater tolerance to thermal variation in the highlands. If this is the case, in the northern Andes we would expect community composition to be relatively homogeneous between highland forests located on the same mountain range, and species composition to be mainly differentiated by dispersal constrains caused by geographical barriers, such as the interandean valleys. The hypothesis about floristic homogenization in highlands of tropical mountains due to species tolerance to high climate variability is analogous to the expected increase of the species' geographic ranges with latitude caused by higher tolerances to environmental variation (Stevens 1992).
The primary goal of the current study was to identify the main factors and processes that determine patterns of community composition for woody plant species inhabiting tropical forests at contrasting elevations. In tropical forests, comparisons of changes in species turnover among forest types with different edaphic and climatic characteristics will improve our understanding of the main processes and factors that control species distributions in environmentally complex ecosystems (Davidar et al. 2007). In particular, we aim to test the hypothesis about the congruence among environmental factors that determine tree species composition at large scales in the lowlands and highlands of tropical forests in this region of Colombia. Thus, based on a survey of 16 1-ha plots conducted along a complex environmental gradient that was largely defined by the Andean mountains, we aimed to answer the following research question: To what extent do environmental factors associated with environmental and/or spatial processes coincide and determine the tree community composition at large scale in lowlands and highlands of montane ecosystems in the northern Andes? We did not find support for a higher habitat-plant specialization in highlands than in lowlands. Improving our understanding of the main factors determining plant community composition in this region will assist in defining conservation strategies for protecting these highly threatened tropical forests (Duque et al. 2014, Myers et al. 2000).
MATERIALS AND METHODS
The study area is located in the northwest region of Colombia between 5° 50' and 8° 61' N; and 74° 61' and 77° 33' W. This region encompasses an altitudinal gradient from sea level to 4000 m.a.s.l. and is highly variable in terms of its topography, climate and soils. The annual precipitation in this region ranges from 1000 to almost 7000 mm. Likewise, the topography and geology are highly variable because of the presence of two mountain ranges influencing patterns of drainage, rainfall and soil fertility at local scales (IGAC 2007).
The study was conducted using data collected from 16 permanent 1-ha forest inventory plots. The permanent plots were distributed across a large geographic area that covers approximately 64 000 km2, mostly in the province of Antioquia. Plot locations span an altitudinal gradient from 50 to 2950 m.a.s.l. (table 1). The average distance between plots was 172.6 km (ranging from 24.5 to 423.1 km). Plots were established in randomly selected areas meeting the following criteria: 1) no signs of recent disturbance; 2) located in either a public or private protected area; 3) located in an accessible area in which work could be performed for long periods; and 4) located in a relatively-secure area without the current presence of guerrillas or any other illegal group. The historical settlement in the region since the 18th century lead to a massive occupation in the central cordillera, which has promoted an intensive deforestation and forest degradation (Cabrera et al. 2011). Overall, the current forest cover only accounts for roughly 30% of the original vegetation (Duque et al. 2014). Four plots (Carepa, El Bagre, Necoclí, and Támesis) are located in small forest fragments (~ 50 ha), and may have faced prior degradation and edge effects such as elevated tree mortality.
All plots were 100 x 100 m with the exception of one plot of 40 x 250 m which was shaped to fit within the only remaining forest fragment in the Támesis geographic area. In each plot, all woody plant individuals (shrubs, trees, palms, and tree ferns) with a diameter at breast height (DBH) ≥ 10 cm were mapped, tagged, and measured. All individuals with a DBH between 1 and 10 cm were counted in a 40 x 40 m subplot located near the center of each plot. Voucher collections were made for each potentially-unique species in each plot. All of the vouchers are kept at the University of Antioquia's Herbarium (HUA). Individuals that could not be identified to the species level were classified as morphospecies based on differences in the morphology of vegetative characters. Approximately 1.2% (333) of individuals were excluded from the analysis due to low-quality vouchers resulting from a lack of clear botanical characters, earlier stages of development, or incorrect enumeration.
For soil sampling, plots were divided into 20 x 20 m quadrats, except for the Támesis plot, which was divided in 10 x 10 m quadrats. We collected samples of the soil A horizon (mineral soil after removing organic layer) from five points in each quadrat. Composite samples (500 g) per quadrat were obtained and air-dried after macroscopic organic matter removal. The chemical properties and texture of the soil samples were analyzed at the Biogeochemical Analysis Laboratory at the Universidad Nacional de Colombia in Medellín. The exchangeable Ca, K, and Mg were extracted with 1 M ammonium acetate and analyzed using atomic-absorption. The available P was extracted with L-ascorbic acid and analyzed using a spectrophotometer UV-VIS. The soil pH was measured in water solution as one part soil to two parts water. The organic matter (O.M.) was determined according to the Walkley and Black volumetric method. Mean values (from quadrats samples) of soil properties per plot, were used for subsequent analyses.
Data analysis. Community composition. Because many species can be artificially classified as unique or rare due to the relatively low density of plots dispersed over our large study area, we included only those species recorded in two or more plots in our analyses. We a priori designated the six plots occurring above 1700 m asl as ''highland'' forests since these plots are located within the cloud-immersion zone and are sometimes exposed to sub-zero temperatures (°C; A. Duque, personal observations). The remaining ten plots (all below 1100 m.a.s.l.) were classified as lowland forests (table 1).
The patterns of community composition in the entire region were defined by means of a Principal Component Analysis (PCA) performed on the Hellinger-transformed species abundances (Legendre and Gallagher 2001, Legendre and Legendre 2012). We then performed an Analysis of Similarities (ANOSIM) with 999 permutations prior to the calculation of the Euclidean distances between the Hellinger-transformed species abundances per plot to identify the different forest types as depicted by the PCA (Borcard et al. 2011, Legendre and Legendre 2012). The ANOSIM test confirms if predefined sampling units are statistically distinct in their species composition under the assumption that compositional intergroup dissimilarities should be greater than intragroup dissimilarities (Oksanen et al. 2016). The species contribution to beta diversity (SCBD), which was derived from the total community composition variance following the methodology proposed by Legendre and De Cáceres (2013), was quantified for each species.
Community composition and environment. Based on the forest types defined by the PCA on the floristic data, we performed a one-way analysis of variance (ANOVA) to evaluate the differences in pH, and the log transformed contents of O.M., Ca, K, Mg, and P. Tukey's tests were performed to compare means. Independent Redundancy Analyses (RDA) based on the Hellinger transformed species abundances was performed to analyze the influence of climate, soil fertility, and spatial processes on compositional patterns (Legendre and Legendre 2012) for both lowland and highland plots. Climatic variables were mean annual rainfall (mm), the number of dry months per year, and the elevation of each plot (m.a.s.l.). Elevation was taken from the NASA shuttle radar topography mission digital elevation model (DEM) at a resolution of 30 arc seconds and was used as a proxy for temperature variation since the Pearson correlation of elevation with both annual mean temperature and minimum temperature was extremely high (r = -0.99). Soil fertility was evaluated by means of the soil pH and the log-transformed content of O.M., Ca, K, Mg, and P. The geographical space was then employed to evaluate the biological spatially driven processes, such as either dispersal limitation (see Jones et al. 2008) or biogeographical similarity. The spatial variables were described by a third-order polynomial of the geographical coordinates, which were centered on their respective means prior to analysis. This method is recommended for modeling spatial structures that operates at large spatial scales (Legendre and Legendre 2012).
In both the lowlands and highlands, the forward selection option was applied to each independent set of explanatory variables (climatic, edaphic, or spatial) to select all significant factors (P < 0.05 after 999 random permutations) explaining the variation of the plant community composition (Borcard et al. 2011). In order to have the most parsimonious RDA model, forward selection was run again on the complete set of selected climatic, edaphic and spatial variables. The distance-decay approach (Nekola and White 1999, Tuomisto et al. 2003) was then applied to analyze the relationship between the floristic similarities and either the log-transformed spatial distances or the environmental distance of the logtransformed variables left in the RDA independent models. The correlation between the floristic distances and spatial or environmental distances was evaluated by a Mantel test (Legendre and Legendre 2012). We transformed the floristic dissimilarities (D) to similarities (S) through a subtraction S = 2-D. A semi-log linear model was then fitted to plot the distance decay of floristic similarity. The approximately linear distance-decay along the logarithm of geographical distance can be used as an explicit prediction (and test) of Hubbell's neutral theory (Hubbell 2001, Tuomisto and Ruokolainen 2006). The RDA and the distance decay methods are complementary methods that aim to explain changes in plant community composition (Tuomisto and Ruokolainen 2006).
RESULTS
In the entire survey, we tallied a total of 26,512 individuals woody plants (DBH ≥ 1 cm) belonging to 1622 species or morphospecies. Of the total species, 959 (59%) could be identified as named species, 577 (36%) were identified only to the genus level, 79 (4.9%) were identified only to the family level, and 7 (0.1%) remained unidentified. In total, 541 species were present in two or more plots and therefore could be included in our analyses.
The number of individuals per plot (including all individuals with DBH ≥ 1 cm found within the 40 x 40 m subplot) ranged between 862 and 2855. The number of species per plot ranged between 55 and 303 (table 1). In the lowlands, there were 358 species (13,303 individuals) that were present in two or more plots, while in highlands there were 169 species (6898 individuals) that occurred in two or more plots.
Definition of forest types. In the entire study area, we identified 3 distinct forest types: one in the highlands and two in the lowlands (figure 1). Along the first PCA axis, which explained 12.7% of the total variation, the highlands were separated from lowlands. The first PCA axis was highly correlated with elevation (r = -0.75), which confirmed the expected strong effect of temperature in determining tree community composition at large scale along the whole elevation gradient from lowlands to highlands. Along the second PCA axis, which explained 11.4% of the total variation, the lowland forests were also divided into two major forest types - henceforth referred to as lowlands 1 and lowlands 2 (figure 1). The ANOSIM confirmed the existence of significant differences in the community composition among the three forest types identified by means of the PCA (R = 0.89, p = 0.001).
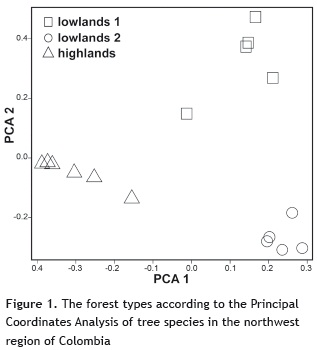
The lowlands 1 group was composed of five plots, including Sapzurro and Puerto Triunfo, which were located at the farthest distance apart of any pair of plots (table 1). The three species with the highest contribution to the total beta diversity belonging to this forest type were Acalypha diversifolia, Clarisia biflora, and Clavija mezii (table 2). The lowlands 2 group was composed of the remaining 5 lowland plots established which were all located within the Cauca basin in the northeast part of the Antioquia province (table 1). The three species with the highest contribution to the total beta diversity belonging to the lowlands 2 forest type were Oenocarpus minor, Pourouma bicolor, and Virola sebifera (table 2). The highlands forest type was dominated by Quercus humboldtii, which was the most abundant species in the entire study area (table 2).
The three highland species that contributed most to beta diversity were Billia rosea, Q. humboldtii, and Wettinia kalbreyeri. Understory species, such as Miconia resima and Palicourea demissa had also an important contribution to total beta diversity. For both the entire dataset and for only the 30 species with the highest contribution to beta diversity, total abundance was highly correlated with SCBD (r = 0.9 and r = 0.83, respectively).
Determinants of community composition and species turnover. Soil fertility in the lowlands 1 forest, as represented by Ca, Mg, and pH, was significantly higher than the soil fertility in the lowlands 2 and the highland forests, which did not differ between each other. However, P concentrations were higher in the highlands than in the lowlands (table 3). In the lowlands, the correlations between Ca, K, and Mg with P were high, but none of them were significant (table 4).
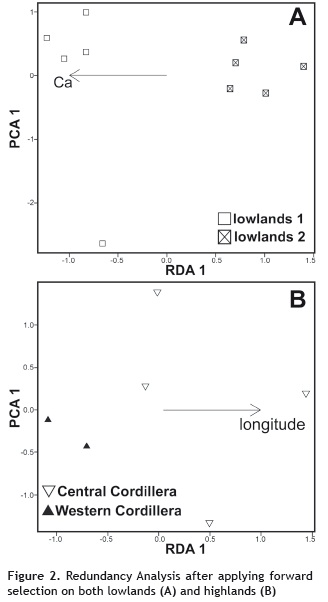
In the lowlands, Ca, longitude, and elevation were retained as significant explanatory variables by the forward selection performed on the separated sets of edaphic, spatial, and climatic explanatory variables, respectively. However, when we ran the forward selection on just Ca, longitude, and elevation only Ca (R2 = 0.24; p = 0.001) remained in the model (figure 2A). Ca was the only variable that remained in the model primarily due to the negative and significant correlation with longitude (r = -0.72) and elevation (r = -0.61). The semi-log linear models confirmed the importance of the increase in Ca contents in soils (RMantel = 0.68; p = 0.006) as well as the increase in the log-transformed geographical distances (RMantel = 0.49; p = 0.004) as significantly associated with compositional turnover in the lowlands (figure 3), but not for elevation (RMantel = 0.25; p = 0.065). In the highlands, the independent forward selection of the RDA on each of the edaphic, spatial, or climatic explanatory set of factors only identified longitude as a significant explanatory variable (R2 = 0.24). The spatial trend in the plant community composition detected by the constrained RDA ordination was determined primarily by the biogeographical location of the plots, which showed a higher floristic similarity between plots located on either the West or the Central Cordillera (figure 2B). However, the semi-log linear model did not support a significant decrease in floristic similarity with the increase in (log-transformed) geographic distances (RMantel = 0.23; p = 0.18; figure 3).
DISCUSSION
We documented the occurrence of three major forest types in the Northern Andean study region that were dominated by different woody plant species. Highland plots (e.g., those located above the cloud-base at 1700 m.a.s.l.) had higher dominance and abundance of species when compared to lowland plots. For example, in the highlands, Q. humboldtii was twice as abundant as any species in the lowlands. Interestingly, the plot located at the lowest elevation within the highland forest type (Anorí) had a mixture of species typical of highlands, such as Billia rosea and Q. humboldtii, and species typical of the lowlands, such as Pouteria torta and Virola sebifera, emphasizing its location within an ecotonal climatic-zone. Gentry (1988) proposed that some plant families, such as Arecaceae and Moraceae, are more abundant and diverse in fertile soils, while others like Burseraceae, Lauraceae, and Sapotaceae, are better-adapted to poorer soils. Although the species that had the highest contribution to the total beta diversity in the richer soils of lowlands belonged to the Moraceae family (Clarisia biflora), our results do not support this generalization. Since some species belonging to the Arecaceae and Moraceae families, such as Pseudolmedia laevis and Oenocarpus minor, respectively, were within the most important species for the total beta diversity in the poorer soils of lowlands, we found it difficult to generalize any likely effect of soils fertility on species distribution in the lowlands at taxonomic levels higher than species.
We assessed the degree to which various climatic, topographic and edaphic variables explain floristic variation in contrasting forest types within this diverse and understudied region of the Neotropics. Our findings did not shed support for the generalized idea about a greater habitat-plant specialization in highlands than in lowlands. Therefore, we reject the main hypothesis about the expected congruence between either environmental or spatial structured factors that determine species composition of tree communities in this region of the northern Andes. Overall, in lowlands, deterministic processes associated with internal ecotones defined by soil properties showed to be the main determinants of tree species composition. In highlands, the biogeographical position along the longitudinal axis created differences in tree community composition between mountain ranges, which were likely driven by isolation due to the barrier imposed by interandean valleys.
In lowland forests, we found that changes in the floristic composition were primarily explained by variation in soil fertility. This suggests that environmental filtering may be the main mechanism determining the structure of the plant community (Gentry 1988, Tuomisto et al. 2003). The fact that the Puerto Triunfo plot appeared as part of the lowlands 1 forest type, even though this plot is located 400 km away from some of the other lowland 1 plots, and is separated from the other lowland 1 plots by the mountain ranges of the Central and West Cordilleras of Colombia and the coastal plains in the north, supports the idea that sites with more similar soil characteristics tend to have greater similarity in species composition regardless of distance or geographic separation. Therefore, plant communities in the lowlands were clearly non-randomly distributed and the associated tree communities could, to a significant degree, be predicted based on environmental factors (Clark et al. 1999, Gentry 1988). Our findings support the importance of niche differences in structuring tree communities across large spatial scales in the lowlands of the Neotropics (Condit et al. 2013, Gentry 1988).
Although our model identified Ca concentrations as the most-important explanatory variable for species composition in the lowlands, the significant correlation of Ca with both Mg concentration and soil pH, suggests a general soil fertility gradient across the sites that distinguishes forest types within the lowlands. Specifically, the lowlands 1 forests have relatively more fertile soils compared to the lowlands 2 plots. These results are similar to those found in other neotropical rainforests, where concentrations of Ca, K, Mg, and pH controlled patterns of tree species composition in both canopy and understory species (Duque et al. 2002, Phillips et al. 2003). As described by Condit et al. (2013), tree species may depend on several nutrients in concert but with Ca and P generally being the two key elements. In our case, there was little variation and low concentrations of P in the lowland plots, which may explain why it was not identified as an explanatory variable of tree community composition. Perhaps, a very high demand of P in forests with high productivity has depleted the soil stock of this important element.
Geographical distances also played important roles in explaining the assemblages of species in the lowlands, as suggested by the negative and significant relationship between compositional similarity and the log-distances between plots. The fact that longitude was not retained as an explanatory variable in the final RDA model was primarily due to the co-linearity with Ca concentrations, which correlation increases with distance. An additional partial Mantel test showed that geographic distances remained significant on controlling the floristic composition after accounting for the effects of Ca (RPartial.Mantel = 0.38; p = 0.019). Disentangling how much of the influence of spatially-structured soil components in structuring tree communities is due to either the soils effect per se or to spatially-structured biological processes such as dispersal limitation, has previously been debated in plant communities (Duivenvoorden et al. 2002, Duque et al. 2009); in this particular case, it appears that plant community composition within the lowlands is largely determined by soil fertility (Garibaldi et al. 2014). This contention is supported by the fact that plots located on similar soils but with different geologies in different catchments showed high similarity in their species composition. In contrast, in mountainous ecosystems of Costa Rica, plots of canopy tree species located below 1500 m.a.s.l. showed significant spatial autocorrelation at distances of up to 40 to 50 km and biologically-structured factors such as dispersal limitation were proposed as one of the major determinants of tree species distributions (Chain-Guadarrama et al. 2012).
Similar to longitude, in the lowlands, elevation was not retained in the final RDA model due to it being significantly correlated with Ca contents (RMantel = 0.33; p = 0.042). In studies of elevational gradients, local thermal conditions have been found to play an important role in shaping species distributions and compositional patterns (Feeley et al. 2013, Hemp 2005). However, in the lowlands of our study area, the results of both analyses were not conclusive due to the RDA and the semi-log linear models producing different outputs at pinpointing elevation as a significant factor. On the one side, according to the RDA model, we found a small but still significant effect of temperature on the plant communities that was masked by the correlation with Ca. Contrasting this, the log-linear model that tests the neutral hypothesis of a constant decrease in species similarity with increasing difference in elevation, did not support a significant effect of elevation on structuring tree communities below 1000 m.a.s.l. (see also Chain- Guadarrama et al. 2012, Gentry 1995). A partial Mantel test showed that elevation did not remain as a significant explanatory variable of floristic changes after removing the effect of Ca (RPartial.Mantel = 0.19; p = 0.11). More detailed analysis about thermal ranges of tree species distributions at large geographical scales (e.g., Feeley et al. 2013) are still needed to improve the effectiveness of conservation strategies aimed at ameliorating global warming effects in lowland tropical forests.
In contrast to expectations, in the highland forest plots, longitude was the only explanatory variable significantly associated with tree community composition in the RDA analysis (figure 3). Similar to elevation in lowlands, in the highlands, the expected decrease in species similarity with the increase in the geographic distance proposed by the neutral theory (Hubbell 2001) was rejected. Thus, the significance of longitude in the RDA model as an explanatory variable of floristic composition is in accord with the observations that tree species composition was more similar between plots established on the same Cordillera (either Central or Western; figure 2B). This may be due to the Cauca river valley constraining the movement of propagules between mountain chains producing an anisotropic effect (sensu Tuomisto and Ruokolainen 2006) from east to west (and vice versa) in the tree community composition. However, we acknowledge that additional studies with increased sample sizes are needed to clarify this floristic-geographic differentiation.
Another explanation for the lack of environmental effects on floristic composition in the highlands is the fact that soil fertility was fairly homogeneous between sites. The highland plots were located on metamorphic and volcanic intrusive rocks that originate in the West and Central Cordilleras, respectively. It is expected that the volcanic intrusive rocks should have high soil fertility because of its geological origin and high concentrations of volcanic ashes. Therefore, some unknown processes may have driven soils fertility or nutrient release to similar levels independent of their geological origin promoting the relative floristic homogeneity found in this study, and thus, the lack of significance of soils in determining tree communities in highlands. In the highlands, for example, neither Ca nor P had any significant influence on structuring highland tree communities. The lack of importance of P in the models could indicate that P is not a limiting resource in these forests. Alternatively, the relatively high P concentrations found in highland soils (in relation to lowlands) may not be available for the plant uptake due to the low pH of the soils that could promote P precipitation. However, a complementary explanation to the lack of importance of environmental factors in structuring the highland tree communities may be the existence of broad distributional ranges for many species. Broad distributional ranges of species that occur within the cloud-immersion zone may be the consequence of the regionally homogeneous soils fertility as well as physiological adaptations to extreme climatic conditions and variation (Stevens 1992).
The results of this study proposes that different factor may play important roles on determining the tree communities inhabiting the quite diverse forests of the northern Andes in Colombia. Although in the whole study area either elevation or temperature was the most important factor determining tree communities (Duque et al. 2015), our findings proposed that the extent and nature of the main factors determining species composition varied according to the altitudinal position. Although additional assessments of this question are needed, they emphasize on the need to develop different strategies to protect the already quite endangered forests of this region. In cases such as those found in lowlands, including within reserves as much as possible of the environmental variation will be a key issue for conservation. In highlands, on contrary, the larger the reserve the more effective the conservation strategy will be.
It is important to note that many Andean forests have been highly degraded and fragmented (Myers et al. 2000), which may limit seed dispersal because of the lack of connectivity and forest continuity in the region. In fact, in the northwest Andean lowlands of Colombia and their surroundings, where forest occupancy by human beings spans several centuries, the local populations of plant species may be a product of physical and genetic isolation. Species isolation by habitat loss and forest fragmentation represents a major threat to this extremely diverse area of the Neotropics and will likely drive local and global extinctions if we do not stop the current deforestation trends (Duque et al. 2014). Since tropical Andean ecosystems, and in particular highland forests, have received less attention than their lowland counterparts, our findings help to expand the current knowledge on these important habitats.
ACKNOWLEDGEMENTS
We thank Fundación Berta Arias, CORANTIOQUIA, Fundación Colibrí, Empresas Pecuarias del Bajo Cauca, Mineros S. A., Rodrigo Celis, and Andrés Upegui for allowing us to install and maintain the plots on their properties. The authors are grateful to James Dalling for his comments and suggestions to a prior version of this manuscript. We also thank Herbarium Universidad de Antioquia for allowing the identification and storage of collections. A. Duque was funded by a Visitant Researcher Fulbright Colombia grant (2014-2015) that was hosted in Miami (FL, U.S.A.) by the International Center for Tropical Botany at Florida International University and the Fairchild Tropical Botanic Garden. K. J. Feeley is supported by the U.S. National Science Foundation (DEB-1350125).
REFERENCES
Aiba S, Kitayama K. 1999. Structure, composition and species diversity in an altitude-substrate matrix of rain forest tree communities on mount Kinabalu, Borneo. Plant Ecology, 140 (2): 139-157. [ Links ]
Borcard D, Gillet F, Legendre P. 2011. Numerical ecology with R. New York: Springer. p. 306. [ Links ]
Cabrera E, Galindo G, Vargas DM. 2011. Protocolo de procesamiento digital de imágenes para la cuantificación de la deforestación en Colombia, nivel nacional escala gruesa y fina. Bogotá D. C. (Colombia): Instituto de Hidrología, Meteorología, y Estudios Ambientales-IDEAM. p. 44. [ Links ]
Chain-Guadarrama A, Finegan B, Vilchez S, Casanoves F. 2012. Determinants of rain-forest floristic variation on an altitudinal gradient in southern Costa Rica. Journal of Tropical Ecology, 28 (5): 463-481. [ Links ]
Clark DB, Palmer MW, Clark DA. 1999. Edaphic factors and the landscape-scale distributions of tropical rain forest trees. Ecology, 80 (8): 2662-2675. [ Links ]
Clinebell II, RR, Phillips OL, Gentry AH, Stark N, Zuuring H. 1995. Prediction of neotropical tree and liana species richness from soil and climatic data. Biodiversity and Conservation, 4 (1): 56-90. [ Links ]
Condit R, Pitman N, Leigh EG, Chave J, Terborgh J, Foster RB, Nuñez P, Aguilar S, Valencia R, Villa G, Muller-Landau HC, Losos E, Hubbell SP. 2002. Beta-diversity in tropical forest trees. Science, 295 (5555): 666-669. [ Links ]
Condit R, Engelbrecht B, Pino D, Pérez R, Turner L. 2013. Species distributions in response to individual soil nutrients and seasonal drought across a community of tropical trees. Proceedings of the National Academy of Science, 110: 5064-5068. [ Links ]
Davidar P. Rajagopal B, Mohandass D, Puyravaud JP, Condit R, Wright SJ, Leigh EG. 2007. The effect of climatic gradients, topographic variation and species traits on the beta diversity of rain forest trees. Global Ecolology and Biogeography, 16 (4): 510-518. [ Links ]
Duivenvoorden, JF, Svenning J-Ch, Wright SJ. 2002. Beta diversity in tropical forests. Science, 295(5555): 636-637. [ Links ]
Duque Á, Sánchez M, Cavelier J, Duivenvoorden JF. 2002. Different floristic patterns of woody understorey and canopy plants in Colombian Amazonia. Journal of Tropical Ecology, 18 (4): 499-525. [ Links ]
Duque Á, Phillips JF, Hildebrand P, Posada CA, Prieto A, Rudas A, Suescún M, Stevenson P. 2009. Distance decay of tree species similarity in protected areas on terra firme forests in Colombian Amazonia. Biotropica, 41 (5): 599-607. [ Links ]
Duque Á, Feeley K, Cabrera E, Callejas R, Idárraga Á. 2014. The dangers of carbon-centric conservation for biodiversity: a case study in the Andes. Tropical Conservation Science, 7 (2): 178-193. [ Links ]
Duque Á, Stevenson P, Feeley K. 2015. Thermophilization of adult and juvenile tree communities in the northern tropical Andes. Proceedings of the National Academy of Science, 7: 10744-10749. [ Links ]
Engelbrecht BM, Comita LS, Condit R, Kursar AT, Tyree MT, Turner BL, Hubbell SP. 2007. Drought sensitivity shapes species distribution patterns in tropical forests. Nature, 447 (7140): 80-82. [ Links ]
Feeley K, Hurtado J, Saatchi S, Silman M, Clark D. 2013. Compositional shifts due to climate-driven species migration. Global Change Biology, 19 (11): 3472-3480. [ Links ]
Garibaldi C, Nieto-Ariza B, Macía MJ, Cayuela L. 2014. Soil and geographic distance as determinants of floristic composition in the Azuero Peninsula (Panama). Biotropica, 46 (6): 687-895. [ Links ]
Gentry AH. 1982. Patterns of neotropical plant species diversity. In: Hecht MK, Wallace B, Prance ET, editors. Evolutionary biology. New York (USA): Plenum Press. Vol. 15: 1-84. [ Links ]
Gentry AH. 1988. Changes in plant community diversity and floristic composition on geographical and environmental gradients. Annals of the Missouri Botanical Garden, 75 (1): 1-34. [ Links ]
Gentry AH. 1995. Patterns of diversity and floristic composition in neotropical montane forests. In: Churchill SP, Balslev H, Forero E, Luteyn J, editors. Biodiversity and conservation of neotropical montane forests. New York: New York Botanical Garden. p. 103-126. [ Links ]
Hemp A. 2005. Continuum or zonation? Altitudinal gradients in the forest vegetation of Mt. Kilimanjaro. Plant Ecology, 184 (1): 27-42. [ Links ]
Hubbell SP. 2001. The unified neutral theory of biodiversity and biogeography. Princeton: Princeton University Press. p. 392. [ Links ]
IGAC. 2007. Antioquia. Características geográficas. Bogotá (Colombia): Instituto Geográfico Agustín Codazzi. p. 320. [ Links ]
John RC, Dalling JW, Harms KE, Yavitt JB, Stallard RF, Mirabello M, Hubbell SP, Valencia R, Navarrete H, Vallejo M, Foster RB. 2007. Soil nutrients influence spatial distributions of tropical tree species. Proceedings of the National Academy of Sciences, 104 (3): 864-869. [ Links ]
Jones MM, Tuomisto H, Borcard D, Legendre P, Clark DB, Olivas PC. 2008. Explaining variation in tropical plant community composition: influence of environmental and spatial data quality. Oecologia, 155 (3): 593-604. [ Links ]
Jones MM, Szyska B, Kessler M. 2011. Microhabitat partitioning promotes plant diversity in a tropical montane. Global Ecology and Biogeography, 20: 558-569. [ Links ]
Legendre P, Gallagher ED. 2001. Ecologically meaningful transformations for ordination of species data. Oecologia, 129 (2): 271-280. [ Links ]
Legendre P, Legendre L. 2012. Numerical ecology. 3th edition. Amsterdam: Elsevier. p. 1006. [ Links ]
Legendre P, De Cáceres M. 2013. Beta diversity as the variance of community data: dissimilarity coefficients and partitioning. Ecology Letters, 16 (8): 951-963. [ Links ]
Myers N, Mittermier RA, Mittermier CG, da Fonseca GAB, Kent J. 2000. Biodiversity hotspots for conservation priorities. Nature, 403 (6772): 853-858. [ Links ]
Nekola JC, White PS. 1999. The distance decay of similarity in biogeography and ecology. Journal of Biogeography, 26 (4): 867-878. [ Links ]
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O'Hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner H. 2016. Vegan: Community Ecology Package. R package ver. 2.3-5 [Internet]. Accessed: 03 May 2016. Available from: https://cran.r-project.org/web/packages/vegan/index.html. [ Links ]
Phillips OL, Núñez-Vargas P, Monteagudo AL, Peña-Cruz A, Chuspe- Zans ME, Galiano-Sánchez W, Yli-Halla M, Rose S. 2003. Habitat association among Amazonian tree species: a landscape-scale approach. Journal of Ecology, 91 (5): 757-775. [ Links ]
Stevens GC. 1992. The elevational gradient in altitudinal range: an extension of Rapoport's latitudinal rule to altitude. American Naturalist, 140 (6): 893-911. [ Links ]
Tuomisto H, Ruokolainen K. 2006. Analyzing or explaining beta diversity? Understanding the targets of different methods of analysis. Ecology, 87 (11): 2697-2708. [ Links ]
Tuomisto H, Ruokolainen K, Yli-Halla M. 2003. Dispersal, environment, and floristic variation of western Amazonian forests. Science, 299 (5604): 241-244. [ Links ]
Vásquez JA, Givnish TJ. 1998. Altitudinal gradients in tropical forest composition, structure, and diversity in the sierra de Manantlan. Journal of Ecology, 86 (6): 999-1020. [ Links ]
Vormisto J, Svenning J-Ch, Hall P, Balslev H. 2004. Diversity and dominance in palm (Arecaceae) communities in terra firme forests in the western Amazon basin. Journal of Ecology, 92 (4): 577-588. [ Links ]

