










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActualidades Biológicas
Print version ISSN 0304-3584
Actu Biol vol.38 no.105 Medellín July/Dec. 2016
https://doi.org/10.17533/udea.acbi.v37n105a05
ARTÍCULOS DE INVESTIGACIÓN
doi: 10.17533/udea.acbi.v37n105a05
Nuevos registros de macromicetos para Colombia
New records of macrofungi from Colombia
Paola Lombana-Álvarez1,4, Jorge A. Monterroza-Álvarez1,5, Luis F. Chamorro-Quiroz1,6, Ana E. Franco-Molano2, Iris R. Payares-Díaz3
1 Grupo de investigación en ecología y zoología (GIZEUS). Universidad de Sucre. Sincelejo (Sucre), Colombia.
2 Laboratorio de Taxonomía y Ecología de Hongos, Instituto de Biología Universidad de Antioquia. A. A. 1226 Medellín, Colombia. afranco@quimbaya.udea.edu.co
3 Grupo de investigación biotecnología vegetal. Universidad de Sucre. Sincelejo (Sucre), Colombia.iris.payares@unisucre.edu.co
Correos electrónicos: 4 lombanapao@gmail.com; 5 jorge17ma@hotmail.com; 6 chamorroad@hotmail.com.
Recibido: enero 2015; aceptado: marzo 2016 (Received: January 2015; accepted: March 2016).
Resumen
Se presentan los resultados del trabajo realizado en la Estación Meteorológica de CARSUCRE, en bosque seco tropical del municipio de Colosó, departamento de Sucre (Colombia), donde se efectuaron transectos lineales de 100 m al lado del arroyo Sereno y parte alta del bosque (cerro), durante los meses de febreroagosto y noviembre del año 2011, se realizaron un total de 182 colecciones, distribuidas en los filos Ascomycota (4 géneros y 11 especies) y Basidiomycota (31 géneros con 59 especies), se logró recolectar 39 registros nuevos para Sucre y siete para Colombia. Este trabajo constituye un estudio pionero para el departamento de Sucre, debido a que no existe información acerca de los hongos silvestres presentes en el bosque seco tropical (Bs-T).
Palabras clave: bosque seco tropical, Basidiomycota, diversidad, macrohongos, Sucre.
Abstract
We present the results of a study carried out in the tropical dry forest at the Meteorological Station of CARSUCRE, in the municipality of Colosó, Sucre (Colombia). We used line transects of 100 m in length, along the creek Sereno and on the top of the hills; we sampled along such transects from February to August and in November 2011. A total of 182 specimens were collected, belonging to the phyla Ascomycota (4 genera and 11 species) and Basidiomycota (31 genera and 59 species). Moreover, we report 39 new records for Sucre and seven new records for Colombia. Finally, this work is a pioneering study for the Department of Sucre, because there is no information about wild fungi in dry tropical forest (Bs-T).
Key words: Basidiomycota, diversity, macrofungi, Sucre, tropical dry forest.
INTRODUCCIÓN
Colombia se encuentra dentro de una zona tropical privilegiada del mundo y por su variedad de climas, permite el crecimiento y la generación de múltiples formas de vida, que hacen que sea un país megadiverso (Restrepo et al. 2005). A pesar de la abundancia de hongos y del papel que juegan en los ecosistemas, estos han sido poco estudiados. Para el país se han registrado 1.239 especies de macrohongos, la mayoría de los registros (83%) corresponden a colecciones de la región Andina, seguida por la costa Caribe con 6%. El 11% restante distribuido en las demás regiones del país (Franco-Molano y Uribe 2000, Franco-Molano et al. 2010, Vascos-Palacios y Franco-Molano 2013).
Además, entre la gran variedad de hábitats para el crecimiento de macromicetes que ofrece Colombia, se destacan los bosques secos tropicales (Bs-T), que se define como la formación vegetal que presenta una cobertura boscosa continua y que se distribuye entre los 0-1.000 m de altitud con temperatura promedio superior a los 24 °C y precipitación que varía entre los 700 y 2.000 mm anuales. Es probable entonces que la composición taxonómica y la estructura ecológica de los hongos en estos bosques pueda ser única y hasta el momento desconocida y quizás lo sea para siempre, dado el ligero deterioro ocasionado por la rápida fragmentación de los bosques (García-Lemos y Bolaños-R 2010).
En estudios de García-Lemos y Bolaños-R (2010) señalaron que dadas las condiciones de poca humedad de los bosques secos tropicales es de esperar que la abundancia de especies de hongos en el Bs-T, sea más baja que en los bosques húmedos tropicales, sin embargo, el bosque seco posee características físicas, vegetales y microclimáticas que permiten la presencia y gran abundancia de familias, géneros y especies de hongos basidiomecetes.
Para el departamento de Sucre, no existe ningún estudio sobre macromicetes, por tal razón es sumamente importante empezar a realizar trabajos de investigación relacionados con estos organismos, especialmente en un ecosistema tan frágil y en peligro de desaparecer por acción antrópica como lo es el bosque seco tropical. Este trabajo tiene como objetivo contribuir a incrementar el conocimiento de las especies de macromicetes de esta región y el país, y así se pueda generar estrategias para su manejo y conservación de este tipo de ecosistemas. Como resultado de este trabajo se registran siete especies por primera vez para Colombia.
MATERIALES Y METODO
Área de estudio. El trabajo se llevó a cabo en la Estación Meteorológica de CARSUCRE, localizada en las estribaciones de la serranía de Coraza-Montes de María, situada en el municipio de Colosó (9° 29' N; 75° 21' O), en la planicie del litoral del Caribe colombiano y localmente recibe el nombre de Montes de María (figura 1). presentando un clima cálido y seco; dado que no existe un relieve montañoso que incida significativamente sobre la temperatura. La zona está caracterizada por bosques higrotropofíticos, subhigrofíticos y freatofíticos (Cuervo- Díaz et al. 1986), con una extensión de 6.452 ha, de las cuales 3.000 ha son de bosque primario (Galván et al. 2009). Esta zona es de gran importancia biótica, debido a que está constituida por la formación de bosque seco tropical (Bs-T) (Espinal 1985, IavH 1998, Murphy y Lugo 1986).
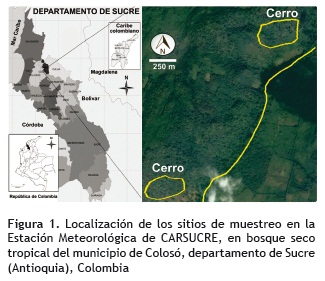
Según Barbosa et al. (1988) se distinguen dos tipos de formaciones vegetales; la primera, conformada por vegetación de ladera que corresponde a bosques de tipo higrotropofíticos calcícolas, desarrollados en fuertes pendientes con afloramientos de rocas calcáreas de origen coralino; entre las especies vegetales se distinguen Bursera simaruba (L.) Sarg, Pseudobombax septenatum (Jacq.) Dugand, Hura crepitans L. y Uribea tamarindoides Dugand y Romero. La segunda formación, conformada por vegetación ribereña, que corresponde a bosques de galería desarrollados en los márgenes de los arroyos, en suelos franco arcillosos, ricos en nutrientes y buena disponibilidad de recursos hídricos presentando elementos florísticos como Anacardium excelsum asociados a Brownea ariza, Enterolobium cyclocarpum y Hura crepitans.
Metodología. Los muestreos se llevaron a cabo en el año 2011 durante 11 visitas al bosque de la Estación Meteorológica de CARSUCRE. De febrero a junio se realizaron recolectas en el área aledaña al arroyo el Sereno y entre los meses de julio, agosto y noviembre se muestrearon los cerros.
Se determinaron en total 27 transectos lineales de 100 m y sobre estos, subparcelas de 7 x 7 m con separación de 10 m entre ellas (Vázquez 2008), en cada visita siempre se hizo la revisión de ramas, arbustos, árboles caídos, hojarasca y suelo (García-Lemos y Bolaños-R 2010).
Los hongos fueron descritos macroscópicamente, siguiendo las recomendaciones de Franco-Molano et al. (2000, 2005) y Ryvarden y Johansen. (1980). Los colores se describieron usando como referencia la tabla de colores de The Online Auction Color Chart (2004). Los especímenes se secaron en un horno a una temperatura de 45-50 °C y posteriormente, se empacaron en bolsas plásticas y se rotularon.
Las descripciones microscópicas se llevaron a cabo en el laboratorio de Biología II de la Universidad de Sucre, los montajes para el microscopio se hicieron en KOH al 5%, reactivo de Melzer y Rojo Congo. Las estructuras fueron observadas y descritas con el objetivo de 100X. Los nuevos registros se encuentran depositados en el herbario de la Universidad de Sucre (HUS).
Para la identificación de los especímenes se emplearon claves taxonómicas especializadas, se comparó con material de referencia del Herbario de la Universidad de Antioquia (HUA) y se contó con la ayuda de especialistas.
RESULTADOS
Durante el estudio se realizaron 182 colecciones de las cuales 119 pertenecen al filo Basidiomycota y 63 al filo Ascomycota; de estas, se identificaron en total 39 especies y 31 morfoespecies (anexo 1). El orden Polyporales con 46%, fue el más abundante por su papel ecológico de descomponer madera. Seguido por el orden Agaricales con 17%, Xylariales 13%, Hymenochaetales 12%, Auriculariales 7% y Pezizales 3%; los menos representativos, Geastrales y Tremellales con 1% cada uno (figura 2).

Este trabajo representa el primer registro de macromicetes (39 especies) para el departamento de Sucre, siete son registros nuevos para el país y se describen a continuación.
Echinochaete brachypora (Mon.) Ryvarden. (1978)
Basidiocarpos: anuales, solitarios. Píleo: 5 cm de diámetro, dimidiado a semicircular; superficie ligeramente tomentosa, de color blanco hueso cuando fresco (oac909) y de color canela cuando está seco (oac668); margen entero y deprimido cuando está seco. Contexto: hasta 3 mm de grosor, color café claro (oac729). Estípite: 1 cm de longitud, 0,8 mm de diámetro. Excéntrico; superficie finamente tomentosa. Himenóforo: poroide, poros angulares y radialmente elongados, de color beige (oac795) cuando fresco y coloración madera cuando seco (oac701), de 1-3 poros por mm. Olor y sabor: no determinado. Sistema hifal: dimítico; hifas generativas de 3-4 μm de diámetro, con septos fíbulados, paredes delgadas, hialinas en KOH; hifas ligativas, hasta 7 μm de diámetro, con paredes gruesas, de amarillo pálido a café en KOH. Elementos setoides: abundantes en el himenio, de 47-72 x 12-15 μm, color amarrillo a marrón, con espinas laterales, paredes gruesas. Basidios: de 26-36 x 9-14 μm, clavados, biesterigmados, amarillentos en KOH, inamiloides en reactivo de Melzer. Queilocistidios: ausentes. Basidiosporas: de 8,5-10,2 x 4-6 μm, cilíndricas, lisas, de pared delgada, hialinas en KOH, inamiloides en reactivo Melzer. Esporada: no obtenida. Figura 3.
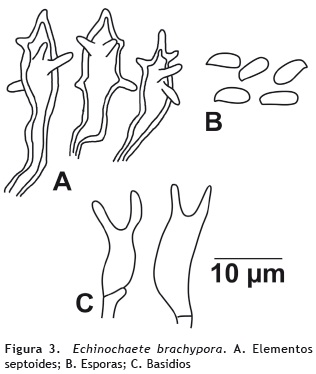
Hábito y hábitat: solitarios, sobre tronco en descomposición.
Distribución. Pantropical: África, Uganda, Tanzania, Malawi, Kenya y Burundi (Ryvarden y Johansen 1980) (HUS584).
Material estudiado: Colombia. Sucre. Colosó. Bosque de la estación meteorológica de CARSUCRE, sobre tronco en descomposición, 20-4-2012, Chamorro, Lombana & Monterrosa (PL090). 08-6-2012, Chamorro, Lombana & Monterrosa (PL0169).
Geastrum rufescens Pers. 1801
Basidiocarpos: en forma de estrella. Exoperidio: proyectándose de 6-8 lóbulos curvados hacia abajo, color pardo (oac721), no higrófano, carnoso. Endoperidio: forma globoso, central, 2,6 cm de diámetro, superfície lisa, glabra, color pardo (oac721). Gleba: color pardo. Opérculo: 6 mm de diámetro, fimbriado, de color pardo. Apófisis: ausente. Micelio basal: color blanco (oac909). Basidiosporas: 3,7 x 4 μm, densamente ornamentadas con espinas cortas y romas, de color pardo. Paredes delgadas, castañas en KOH, inamiloides en Melzer. Basidios: ausentes. Capicilio: formado por hifas de paredes gruesas, de 4-7 μm de diámetro, lumen estrecho, hialinas en KOH y dextrinoides en reactivo de Melzer. Figura 4.
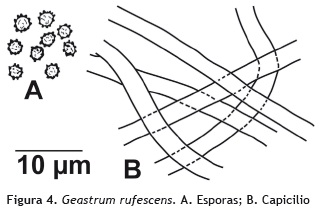
Hábito y hábitat: solitario, sobre suelo.
Distribución. Cosmopolita (Mata 1999).
Material estudiado: Colombia. Sucre. Colosó. Bosque de la estación meteorológica de CARSUCRE, sobre tronco en descomposición, 11-8-2012, Chamorro, Lombana & Monterrosa (JM10), 11-8-2012, Chamorro, Lombana & Monterrosa (LCH4), 24-11-2012, Chamorro, Lombana & Monterrosa (PL25) (HUS586).
Gloeoporus thelephoroides (Hook.) G. Cunn. 1965
Basidiocarpos: sésiles, de consistencia subcarnosa, anuales. Píleo: de 5 cm de diámetro, flabeliforme a dimidiado, plano a ligeramente convexo; superficie velutinosa, ligeramente zonada, color blanco hueso (oac909); margen agudo, ondulado. Himenóforo: poroide, con poros redondos, pequeños, de 6-8 por mm, de color pardo (oac646); tubos de hasta 1 mm de grosor. Olor y sabor: no determinado. Contexto: hasta 1 mm de grosor, color madera, fibroso. Sistema hifal: monomítico hifas generativas de 2-4 μm de diámetro, con septos simples, con paredes delgadas, con o sin ramificaciones, hialinas en KOH, inamiloides en reactivo de Melzer. Basidios: ausentes. Queilocistidios y Pleurocistidios: ausentes. Basidiosporas: de 3,5-5 x 1-1,2 μm, cilíndricas, a alantoides, de paredes delgadas, pequeñas, hialinas en KOH e inamiloides en reactivo de Melzer. Esporada: no obtenida. Figura 5.
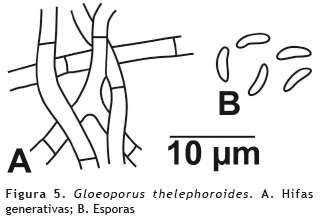
Hábito y hábitat: solitarios, sobre tronco en descomposición.
Distribución. Pantropical; en África registrado desde Sierra Leona hasta Rhodesia y Madagascar (Ryvarden y Johansen 1980).
Material estudiado: Colombia. Sucre. Colosó. Bosque de la estación meteorológica de CARSUCRE, sobre tronco en descomposición, 19-2-2012, Chamorro, Lombana & Monterrosa (LCH066) (HUS587).
Hohenbuehelia fluxilis (Fr.) P.D. Orton 1964
Basidiocarpo: sésil. Píleo: 2 cm de diámetro, convexo a plano; superficie tomentosa, gelatinosa, color blanco (aoc909); margen entero, enrollado. Contexto: hasta 0,2 cm de grosor, de color blanco, gelatinoso. Olor y sabor: dulce. Lamelas: hasta 0,1 cm de ancho, concurrentes, cercanas, de color blanco; margen entero. Lamélulas: de dos longitudes. Esporada: blanca. Basidiosporas: de 8-10 x 4 μm, ovoides a elipsoides, hialinas en KOH, inamiloides en reactivo de Melzer. Basidios: 20-23 x 5-7 μm claviformes, biesterigmados, con paredes delgadas, superficie lisa, hialinos en KOH, inamiloides en Melzer. Queilocistidios: ausentes. Pleurocistidios: 23-32 x 5 μm, con paredes delgadas, hialinos en KOH, inamiloides en Melzer. Trama himenoforal: regular, hialina en KOH, inamiloides en reactivo de Melzer, con hifas de paredes delgadas. Fíbulas presentes. Figura 6.
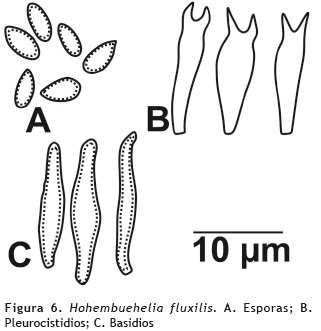
Hábito y hábitat: gregario, sobre tronco en descomposición.
Distribución. En el área del Caribe (Pegler 1983) y países bajos, en general toda Europa (Bas et al. 1995).
Material estudiado: Colombia. Sucre. Colosó. Bosque de la estación meteorológica de CARSUCRE, sobre tronco en descomposición, 24-11-2012, Chamorro, Lombana & Monterrosa (PL22) (HUS619).
Podoscypha nitidula (Berk.) Pat. 1903
Basidiocarpos: estipitados. Píleo: 4,0-4,5 cm de longitud, plano-convexo; superficie lisa, color beige (oac814); margen lobulado, anillos concéntricos color café claro (oac729). Estípite: hasta 1cm de longitud, cilíndrico, central, leñoso; superficie color crema (oac816). Olor y sabor: no determinado. Contexto: hasta 1 mm de grosor, color marrón. Sistema hifal: dimítico; hifas generativas de 4-5 μm de diámetro, con septos simples, de paredes delgadas, hialinas en KOH; hifas esqueléticas de 6-8 μm de diámetro, con paredes gruesas, hialinas en KOH, e inamiloides en reactivo de Melzer. Basidios: de 27- 38 x 9-11 μm, clavados, tetraesterigmados, de paredes delgadas, hialinos en KOH, inamiloides en reactivo de Melzer. Queilocistidios y pleurocistidios: ausentes. Basidiosporas: 4,6-5 x 3,3 μm, elipsoides a subglobosas, hialinas en KOH e inamiloides en Melzer. Esporada: no obtenida. Figura 7.
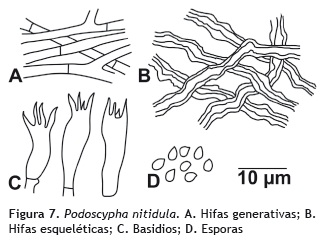
Hábito y hábitat: solitario, sobre tronco en descomposición.
Distribución. Australia, Centro y Sur América (May et al. 2003).
Material estudiado: Colombia. Sucre. Colosó. Bosque de la estación meteorológica de CARSUCRE, sobre tronco en descomposición, 11-8-2012, Chamorro, Lombana & Monterrosa (PL11), 10-8-2012, Chamorro, Lombana & Monterrosa (PL5) (HUS618).
Phellinus lamaoensis (Murrill) Pat. 1923
Basidiocarpos: dimidiados, de consistencia leñosa, perennes. Píleo: 2-7 cm de diámetro; superficie con fuertes zonas concéntricas de color pardo (oac780) escasamente tomentoso; margen entero a lobulado. Estípite: ausente. Himenóforo: poroide, tubos estratificados por capas, 0,1 mm de profundidad; poros redondos, color marrón (oac523), de 7-9 poros por mm. Olor y sabor: no determinado. Contexto: hasta 2.5 mm de grosor, color amarrillo pálido (oac795). Sistema hifal: dimítico; hifas generativas de 4-5 μm de diámetro, con paredes delgadas, hialinas a amarillo pálido en KOH, inamiloides en Melzer; hifas esqueléticas de 3-5 μm de diámetro, abundantes, con paredes gruesas, de amarillas a marrón en KOH, inamiloides en Melzer. Setas himeniales: 29-40 μm de diámetro, abundantes, acuminadas, con paredes gruesas, color amarrillo en KOH Basidios: ausentes. Queilocistidios y pleurocistidios: ausentes. Esporada: no obtenida. Basidiosporas: 4 x 5 μm de diámetro, subglobosas, con paredes delgadas, lisas, hialinas en KOH, inamiloides en reactivo Melzer. Figura 8.
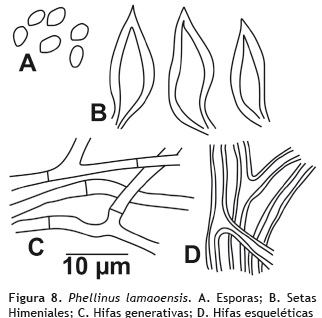
Hábito y hábitat: gregario, sobre tronco en descomposición.
Distribución. Pantropical (Ryvarden y Johansen 1980).
Material estudiado: Colombia. Sucre. Colosó. Bosque de la estación meteorológica de CARSUCRE, sobre tronco en descomposición, 03-2-2012, Lombana, Chamorro & Monterrosa (JM013) (HUS616).
Trametes pavonia (Hook.) Ryvarden 1972
Basidiocarpos: sésiles, coriáceos, anuales. Píleo: 3,5 cm de diámetro, semicircular, de color blanco a crema (oac816); superficie lisa, con zonas concéntricas de color verde hacia la base (oac36); margen simple Himenóforo: color crema a amarillo pálido (oac815), poroide a daedaloides, poros pequeños, de 2-3 por mm, que se oscurecen en KOH. Olor y sabor: no determinado. Contexto: hasta 1 mm de grosor, color blanco. Sistema hifal: trimítico; hifas generativas de 4-5 μm de diámetro, con septos fíbulados, de paredes delgadas, hialinas en KOH; hifas esqueléticas, anchas, poco ramificadas, de 5-6 μm de diámetro, con paredes gruesas, hialinas en KOH; hifas ligadoras de 2-4 μm de diámetro, con paredes gruesas, hialinas en KOH, inamiloides en Melzer. Basidios: 33,5-40,2 x 5-6 μm, clavados, tetraesterigmados, de paredes delgadas, hialinos en KOH, inamiloides en Melzer. Queilocistidios y pleurocistidios: ausentes. Esporada: no obtenida. Basidiosporas: de 6,7-8,2 x 2,4-3 μm, elipsoides, con paredes delgadas, lisas, hialinas en KOH, inamiloides en reactivo Melzer. Figura 9.
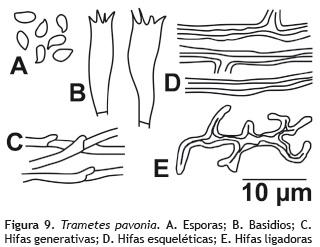
Hábito y hábitat: gregarios, sobre tronco en descomposición.
Distribución. Cosmopolita (Ryvarden y Johansen 1980).
Material estudiado: Colombia. Sucre. Colosó. Bosque de la estación meteorológica de CARSUCRE, sobre tronco en descomposición, 09-6-2012, Chamorro, Lombana & Monterrosa (LCH0178) (HUS617).
DISCUSIÓN
El bosque de la Estación Meteorológica presenta ciertas características biogeográficas que lo catalogan como un bosque seco tropical. En él se presentan altas temperaturas, baja humedad, altitud baja, sequias, suelos constituidos por rocas calcáreas y plantas caducifolias; condiciones que favorecen el desarrollo de las distintas especies registradas en este estudio (Cuervo-Díaz et al. 1986).
El listado de macromicetos de la Estación Meteorológica de CARSUCRE, es el primero que se realiza en bosque seco tropical del departamento de Sucre, en el cual se recolectaron especímenes que representan 39 especies y 31 morfoespecies que constituye una primera aproximación al conocimiento de la diversidad de macromicetos en la Estación Meteorológica de CARSUCRE en el municipio de Colosó.
De las siete especies de macromicetos descritas en este trabajo, cinco son Polyporales (Echinochaete brachyporus, Gloeoporus thelephoroides, Phellinus lamaoensis, Podoscypha nitidula y Trametes pavonia). Estas especies tienen la particularidad de adaptarse a un ambiente seco, lo que las hace depender menos de la humedad y más del sustrato (Pacioni y Lincoff 1981), además poseen características físicas y fisiológicas que les permiten permanecer durante varios años, convirtiéndolos en organismos perennes, gracias a estas condiciones, la probabilidad de frecuentar poliporales es alta durante todo el año.
Los Agaricales representaron el 17% y los Geastrales el 1% de las especies registradas donde Geastrum rufescens (Geastral) y Hohenbuehelia fluxilis (Agarical), se constituyen nuevos registros para el país. Ambas especies presentan factores que influyen sobre su fructificación; como la periodicidad limitada a ciertos periodos del año, particularmente épocas de lluvias y la longevidad moderada, es decir pueden vivir entre una a cuatro semanas (Arnolds 1995), ya que la humedad en este bosque se presenta en ciertos periodos del año donde se presentan con más frecuencias las precipitaciones, adicionalmente a lo anterior, es probable que el arroyo sereno este influyendo sobre la presencia de Geastrum rufescens y Hohenbuehelia fluxilis, debido a que mantiene depósitos con agua en temporada seca, aportando humedad en ciertos puntos dentro de la Estación Meteorológica de CARSUCRE y áreas aledañas.
AGRADECIMIENTOS
Los autores agradecen a la Julieta Carranza por la asesoría brindada en la identificación de las especies en estudio. A los integrantes del grupo de investigación Taxonomía y Ecología de Hongos (TEHOS) de la Universidad de Antioquia. A los miembros del grupo de investigación en Zoología y Ecología de la Universidad de Sucre (GIZEUS) Jorge Díaz y Yeison López y a la Corporación Autónoma Regional de Sucre (CARSUCRE), por permitirnos la recolecta de los especímenes.
REFERENCIAS
Arnolds E. 1995. Problems in measurements of species diversity of macrofungi. En: Allsopp D, Cowell RR, Hawkworth DL, editores. Microbial diversity and ecosystem function. The Netherlands: CAB International. p. 337-352. [ Links ]
Barbosa C, Fajardo-Patiño A, Giraldo H, Rodríguez J. 1988. Proyecto sobre evaluación del hábitat y status del mono tití de cabeza blanca, Saguinus oedipusen Colombia. Unpublished Final Report of Status of Cotton-top Tamarin in Colombia Project. Bogotá (Colombia): INDERENA. p. 123. [ Links ]
Bas C, Kuyper ThW, Noordeloos ME, Vellinga EC. 1995. Flora Agaricina Neerlandica - Critical monographs on the families of agarics and boleti occuring in the Netherlands. Volume 3: Tricholomataceae. Boca Raton (FL, USA): CRC Press. p. 183. [ Links ]
Cuervo-Díaz A, Barbosa-C CE, Ossa V Jaime de la. 186. Aspectos ecológicos y etológicos de primates con énfasis en Alouatta seniculus (Cebidae), de la región de Coloso, Serranía de San Jacinto (Sucre), costa norte de Colombia. Caldasia, 14: 68-70. [ Links ]
Espinal L. 1985. Geografía ecológica del departamento de Antioquia. Revista de la Facultad Nacional de Agronomía, 38 (1): 24-39. [ Links ]
Franco-Molano AE, Corales A, Vasco-Palacios. 2010. Macrohongos de Colombia II. Listado de especies de los órdenes Agaricales, Boletales, Cantharellales y Russulales (Agaricomycetes, Basidiomycota). Actualidades Biológicas, 32 (92): 89-114. [ Links ]
Franco-Molano AE, Uribe-Calle E. 2000. Hongos Agaricales y Boletales de Colombia. Biota Colombiana, 1 (1): 25-43. [ Links ]
Franco-Molano AE, Aldana R, Halling R. 2000. Setas de Colombia (Agaricales, Boletales y otros hongos). Guía de campo. Medellín (Antioquia): Colciencias-Universidad de Antioquia. p. 156. [ Links ]
Franco-Molano AE, Vasco-Palacios, López-Quintero CA, Boekhout T. 2005. Macrohongos de la región del medio Caquetá-Colombia, Guía de campo, Grupo Taxonomia y Ecologia de Hongos. Medellín (Antioquia): Multimpresos, p. 211. [ Links ]
Galván-Guevara S, Sierra I, Gómes H, De la Ossa VJ y Fajardo-Patiño A. 2009. Biodiversidad en el área de influencia de la estación primates de Colosó, Sucre, Colombia. Revista Colombiana Ciencia Animal, 1 (1): 95-118. [ Links ]
García-Lemos A, Bolaños-R AC. 2010. Macrohongos presentes en el bosque seco tropical de la región del Valle del Cauca, Colombia. Revista de Ciencias, Universidad del Valle, 14: 45-54. [ Links ]
IavH (Instituto Alexander Von Humboldt). 1998. Bosque seco tropical (BsT) en Colombia. Programa de inventario de la biodiversidad grupo de exploraciones y monitoreo ambiental GEMA. Bogotá (Colombia): Instituto Alexander Von Humboldt. p. 24. [ Links ]
Mata M. 1999. Macrohongos de Costa Rica. Volumen 1. Santo Domingo de Heredia (Costa Rica): Instituto Nacional de Biodiversidad (INBIO). p. 256. [ Links ]
May TW, Milne J, Shingles S, Jones RH. 2003. Fungi of Australia. Volume 2B. Catalogue and bibliography of Australian Fungi, 2. Basidiomycota p. p. and Myxomycota p. p. Melbourne (Australia): ABRS/CSIRO Publishing. p. 452. [ Links ]
Murphy PG, Lugo AE. 1986. Ecology of tropical dry forest. Annals Review of Ecology and Systematics, 17: 67-68. [ Links ]
Pacioni G, Lincoff G. 1981. Simon and Schuster's Guide to mushrooms. New York: Simon and Schuster. p. 511. [ Links ]
Pegler DN. 1983. Agaric flora of the Lesser Antilles . Kew Bulletin, Additional Series IX. London: Royal Botanic Gardens (Kew ). p. 668. [ Links ]
Restrepo-F GM, Vélez-A PE, Botero-A PA, Pulido-V C. 2005. Reconocimientos de macromicetos asociados al cultivo de Guadua angustifolia en Caldas, Colombia. Manejo Integrado de Plagas y Agroecologia (Costa Rica), 76: 25-31. [ Links ]
Ryvarden L, Johansen I. 1980. A preliminary polypore flora of East Africa. Oslo: Fungiflora. p. 636. [ Links ]
Vascos-Palacios AM, Franco-Molano AE. 2013. Diversity of Colombia macrofungi (Ascomycota-Basidiomycota). Mycotaxon, 121: 100-158. [ Links ]
Vázquez-Mendoza S. 2008. Ecología de macromicetos a lo largo de un gradiente altitudinal en Santa Catarina Ixtepeji, Oaxaca México [Tesis de Maestría]. [Santa Catarina Ixtepeji (México)]: Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional, Unidad Oxacala, Instituto Politécnico Nacional. p. 68. [ Links ]
Anexo
Anexo 1

