











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Human degradation of stream ecosystems has led to the creation of several methods to assess the severity of anthropogenic impacts. An accurate assessment of the state of stream ecosystems would involve the integration of biotic and abiotic components, such as water quality, and surrounding land use, along with the organisms inhabiting the channel and its riparian zone (Gonçalves and de Menezes 2011). Some techniques used to assess stream ecosystems involve the evaluation of physical and chemical parameters, analyses utilizing microbiota, and some of the most common methods utilize biomonitoring indices. Biotic indices, in comparison with other methods, are of great advantage because they are practical, low cost techniques that require relatively little training to implement (Alba-Tercedor and Sánchez-Ortega 1988). Although other water quality methods detect physical-chemical parameters directly, they can only reflect the water quality at the moment of the sampling (Gonçalves and de Menezes 2011). On the other hand, biological indices provide us with an idea of the long term state of the aquatic ecosystem due to the fact that biological communities are constantly being exposed to the possible anthropogenic or natural disturbances (Alba-Tercedor and Sánchez-Ortega 1988).
Biotic indices are often based on tolerance metrics attributed to organisms according to their life history, in which the presence or absence of certain taxa indicates the state of the ecosystem (Washington 1984). While there are a large variety of biomonitoring indices, two commonly used for aquatic ecosystems are the Family Biotic Index (FBI) created by Hilsenhoff (1988) in the United States and the Biological Monitoring Working Party (BMWP) created by Armitage et al. (1983) in England. The FBI has been used in the continental U.S.A (Hilsenhoff 1988), and also adapted for El Salvador (Sermeño Chicas et al. 2010) and Puerto Rico (Gutiérrez-Fonseca and Ramírez 2016). The BMWP has been adapted for a number of regions throughout Latin America, including Costa Rica (MINAE-S 2007), Cuba (Naranjo-López et al. 2005), Colombia (Roldán 2003) and Puerto Rico (Gutiérrez-Fonseca and Ramírez 2016). Both indices use tolerance values for aquatic macroinvertebrates (v. g., shrimp, insects, gastropods, acari) with the goal of obtaining an index score that represents the water quality due to the level of disturbance for a given area (Armitage et al. 1983, Hilsenhoff 1988). Another method for assessing stream impairment is the Stream Visual Assessment Protocol (SVAP) (Bjorkland et al. 2001). The SVAP ignores biotic components and consists of a visual evaluation of physical elements for a stream which results in a score representing the level of impact. In spite of these indices having been used regularly in stream monitoring programs, each individual index does not provide a very holistic view of the stream ecosystem. The FBI and BMWP just evaluate the stream channel using aquatic taxa, but they do not evaluate the riparian zone. The SVAP does include the riparian area and the physical characteristics of the channel, but does not take into account the biota.
When compared with traditional biomonitoring methods, the use of spider communities could result in a useful and more holistic addition to traditional monitoring methods. Spiders are commonly found in a wide range of habitats and because of their sensitivity to changes in prey, habitat, and land use, they have been widely discussed as possible bioindicator taxa (Chan et al. 2009, Kato et al. 2003, Rodrigues and Mendonça 2012). Spiders commonly reside in the riparian zone of streams, preying on emergent aquatic insects and other terrestrial organisms that inhabit these ecotones (Akamatsu et al. 2004, Gillespie 1987, Kato et al. 2003). Riparian spider communities can consist of many different taxa with a variety of different hunting strategies (v. g., web spinners, cursorial hunters, ambush hunters) which should respond differently to spatial structure and prey availability.
In this study, our objective is to assess the utility of changes in spider community metrics (abundance and richness) as possible bioindicators of stream ecosystems. We sought to determine whether these changes in the spider communities within an urban watershed are related to changes in surrounding land use and how they compare to well-known stream monitoring protocols that have recently been adapted for the island of Puerto Rico. We expect that riparian spider communities can be an important component for the creation of a more holistic approach to evaluating stream ecosystems and we propose the use of spiders as indicators of riparian habitats that can be utilized when designing future integrated biotic indices for aquatic ecosystems.
Materials and methods
The selected study area was within the Río Piedras watershed located in northeastern Puerto Rico within the greater metropolitan area of San Juan, which has a population density of around 3,190 people/km2 (Ramírez et al. 2014). The Río Piedras watershed flows through the center of the San Juan metropolitan area and represents a drainage area of 67 km2 with a predominantly urban land use (Lugo et al. 2011). The watershed forms an urban gradient, with the least amount of urbanization around its headwaters and then increasing levels of urbanization in the lowlands to where it empties into the San Juan Bay (Ramírez et al. 2014). We selected six sites that are a part of a long term sampling program for the Río Piedras watershed, with each site representing an individual subwatershed. Sites were numbered 1 to 6 according to the percent vegetation (%VEG) within each subwatershed, with lower numbers representing higher levels of vegetation (Figure 1). Land use data for each subwatershed was adapted from Ramírez et al. (2014). At a site within each subwatershed we identified a 100 m reach transect that contained habitat heterogeneity (v. g., pools, riffles).
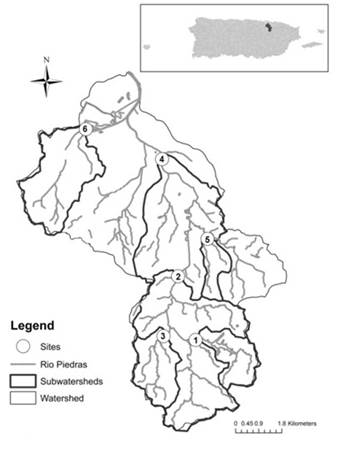
Figure 1 Study sites within the Río Piedras watershed in the greater metropolitan area of San Juan, Puerto Rico. Sites are numerically ordered according to total percent vegetation (%VEG) for each subwatershed. Sites are numbered 1-6 from high to low %VEG.
We utilized a semi-quantitative sampling method with a “D-Net” by manually disturbing all representative microhabitats over a period of 3 minutes. This process was then repeated three times for each site. Sampling was performed following the methods described in the Biological Monitoring Working Party for Puerto Rico (BMWP’PR) and the Family Biotic Index for Puerto Rico (FBI’PR) protocols (Gutiérrez-Fonseca and Ramírez 2016).
BMWP’PR and FBI’PR scores were calculated using the macroinvertebrate data and the respective formulas presented by Gutiérrez-Fonseca and Ramírez (2016). A description of water quality was assigned to each site according to the index score and its corresponding designation. The Stream Visual Assessment Protocol for Puerto Rico (SVAP’PR) as described by Rodríguez and Ramírez (2014) was performed as well. As part of this protocol, we collected data for canopy coverage utilizing a densitometer and we visually determined the percentage of different vegetation substrates present along the riparian area (v. g., trees, grasses). Total percent vegetation (%VEG) for each subwatershed was taken from Ramírez et al. (2014).
Along each 100 m transect spiders were collected by sweep-netting the adjacent riparian vegetation along both sides of the stream using a butterfly net for a period of three minutes. For each site this process was repeated three times. Spiders were then separated manually, stored in 70% ethanol and later identified to the lowest possible taxonomic level. This sampling method was adapted from aquatic macroinvertebrate sampling protocols. With the combined replicates (n=3) we calculated abundance and richness (genera and family) for spider communities at each site. All spiders and aquatic macroinvertebrates were preserved in collections that are a part of long term biomonitoring programs for possible future research of the lab but specimens were all fairly common taxa and were not of any particular taxonomic value to be introduced as an official museum collection.
Linear regression analyses were used to test for correlations between surrounding land use (%VEG) and the three monitoring index scores: BMWP’PR, FBI’PR and SVAP’PR for each of the six sites. Linear regressions were then used to test for correlations between spider community metrics: family richness, genera richness and abundance with %VEG and the three monitoring index scores. All linear regressions were conducted with R statistical software (R Core Team 2012) and p values less than or equal to 0.05 were considered significant.
Results
There was a large degree of variation in vegetation coverage across the watershed associated with the level of surrounding urbanization. In general, a greater amount of %VEG was found at the top of the Río Piedras watershed with decreasing levels of vegetation in the lower areas of the watershed (Figure 1). Maximum and minimum %VEG in the watershed was 83.0% and 22.7%, respectively (Table 1). The differences in %VEG for each subwatershed represented a clear disturbance gradient along the Río Piedras watershed. This gradient was also reflected by the scores of the monitoring indices calculated for each site.
Table 1 Percent vegetation (%VEG), monitoring protocols with scores and stream condition descriptions and the spider community metrics for each site. Monitoring protocols include: BMWP’PR = Biological Monitoring Working Party for Puerto Rico, FBI’PR = Family Biotic Index for Puerto Rico, SVAP’PR = Stream Visual Assessment Protocol for Puerto Rico

Monitoring protocols separated study sites across the watershed and two of the three indices had significant correlations with %VEG (Figure 2). The BMWP’PR resulted in index scores ranging from “excellent” (Site 1) to “bad” condition (Site 5), but showed no significant correlations with %VEG. The FBI’PR classified sites from “good” to “very poor” condition and the SVAP’PR from “high” to “low” physical condition (Table 1). Both, the FBI’PR and SVAP’PR, identified Site 3 as the best in terms of overall stream condition, and Site 6 as the worst (Table 1). Both the FBI’PR (R2 =0.82, p=0.01) and the SVAP’PR (R2 =0.78, p=0.02) were significantly correlated with %VEG (Figure 2).
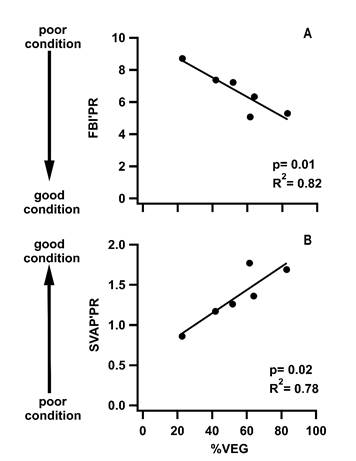
Figure 2 Linear regressions between total percent vegetation for each subwatershed (%VEG) and two of the three monitoring index scores that were found to be significantly correlated. A. Family Biotic Index for Puerto Rico (FBI’PR) and B. Stream Visual Assessment Protocol for Puerto Rico (SVAP’PR)
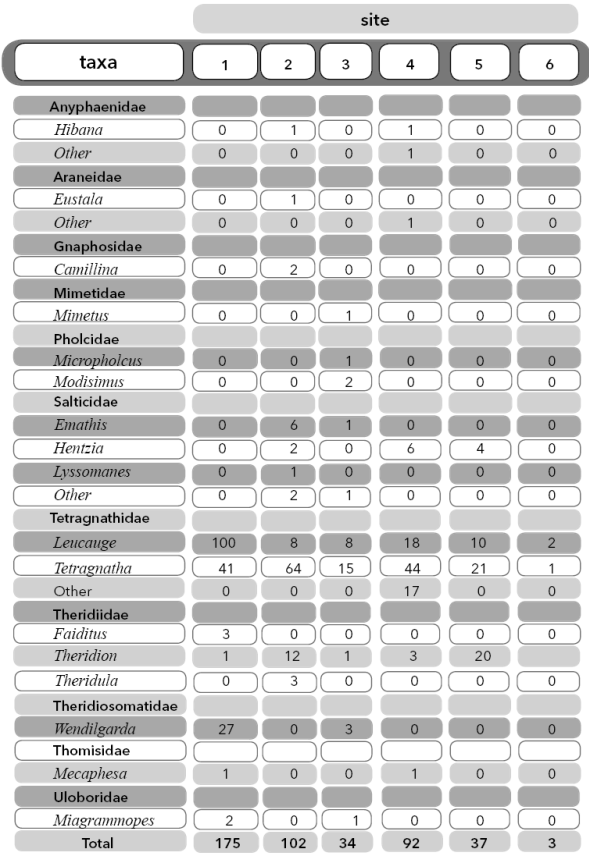
A total of 443 spiders were identified, representing 11 families and 17 genera excluding four unidentified individuals. This included a wide variety of spider taxa with different hunting strategies and web spinning behaviors (v. g., orb webs, sheet webs, dome webs and cursorial hunters). Maximum abundance was observed at Site 1 and 2 (175 and 102 individuals, respectively) and the minimum at Site 6 (3 individuals) (Table 1). The most common family taxon found at each site was Tetragnathidae (horizontal orb-weavers), with its two genera Leucauge and Tetragnatha comprising around 78% of total individuals for all sites (table 2). The rarest family taxa were Araneidae (vertical orb weavers), genus Eustala and Mimetidae (pirate spiders), genus Mimetus, both comprising only 0.4% of total collected individuals (Table 2). Of the 17 genera found, two are considered specialists of aquatic ecosystems: Wendilgarda and Tetragnatha. Site 2 contained the highest genera richness (10 genera) and Site 6 the lowest (2 genera) (table 1). We found Site 3 to have the highest family richness (7 families) and Site 6 the lowest (1 family) (Table 1). The three most common species identified were Tetragnatha boydi, O. Pickard-Cambridge, 1898, Leucauge argyra (Walckenaer, 1841) and Wendilgarda clara Keyserling, 1886. However, due to the fact that immature spiders cannot be accurately identified to species all spiders were therefor identified to the lowest possible level of genera or family.
Table 2 Genera abundance for each family found at the six sites. Taxa catalogued as “Other” were unknown juvenile individuals that could only be identified to family with confidence.
Among the spider community metrics of abundance, family richness, and genera richness, only abundance showed a significant correlation with %VEG (R2 =0.74, p=0.03) (Figure 3). When spider community metrics were analyzed with the three monitoring indices only family richness was found to have significant correlations with FBI’PR (R2 =0.65, p=0.05) and the SVAP’PR (R2 =0.65, p=0.05) (Figure 3).
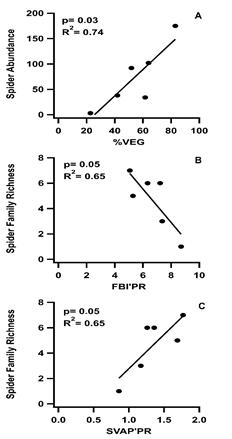
Figure 3 Linear regression analyses between total percent vegetation for each subwatershed (%VEG) and two of the three monitoring index scores that were found to be significantly correlated with at least one of the spider community metrics (spider abundance and spider family richness). A. Total percent vegetation for each subwatershed (%VEG), B. Family Biotic Index for Puerto Rico (FBI’PR) and C. Stream Visual Assessment Protocol for Puerto Rico (SVAP’PR).
Discussion
Our objective was to test whether riparian spider communities respond to changes in the environment along an urban gradient and how changes in spider communities relate to current monitoring protocols as indicators of impacts to stream ecosystems. The six sites ordered by %VEG were found to form a clear gradient within the watershed, which was also reinforced by the results of the monitoring protocols. We found clear changes in spider communities within the urban watershed with a significant correlation between spider abundance and %VEG, as well as family richness and two stream monitoring protocols. The clear relationships between spider abundance and family richness to changes in land use and aquatic monitoring protocols shows the utility for riparian spiders to be considered for future biomonitoring protocols.
The observed changes in the community composition of spiders along the urban gradient can be a reflection of two principal factors: differences in prey availability and/or changes in vegetation. Studies have shown that prey availability (v. g., emerging aquatic insects) can significantly affect the density of riparian spiders along a stream (Kato et al. 2003, Marczak and Richardson 2007) studies such as Chan et al. (2009) have found that communities of riparian spiders increased by around 41% in places where additional substrate was available. Urban streams are often characterized by having altered riparian zones and changes in aquatic insect communities due to anthropogenic activities (Jesús-Crespo and Ramírez 2011, Ramírez et al. 2012) and therefore riparian spider communities should respond strongly to urbanization.
Less urbanized areas, such as Site 1 and Site 2, contained a more heterogeneous mixture of vegetation in comparison to the most urbanized areas, such as Site 6, with a more homogeneous vegetation coverage dominated by grasses. Vegetation structure in the riparian zone influences spider richness, making heterogeneous habitats more diverse than homogeneous ones (Chan et al. 2009, Greenstone 1984, Laeser et al. 2005). Leaser et al. (2005) found that web-weaving spider diversity and abundance decreased in disturbed streams (channelized or with vegetation loss). According to the study, tetragnathid spiders were the only taxa that were the least affected by this kind of disturbance due to their ability to use other substrates such as grasses to build their webs. This would suggest that habitat heterogeneity allows for a greater diversity of taxa and hunting guilds to inhabit the riparian zone.
The significant correlation between spider family richness and two of the three common monitoring methods shows the utility of riparian spiders as possible bioindicators. Both indices, the FBI and SVAP, have been shown to be useful in determining stream condition despite the clear differences in the methods and the focus of the two monitoring protocols (Hilsenhoff 1988, Rodríguez and Ramírez 2014). The FBI index relies strictly on a combination of abundance and family richness of aquatic macroinvertebrates, while the SVAP is a visual assessment of riparian vegetation and other physical characteristics of the stream channel. The SVAP has been described as a good tool in detecting changes in streams through time (Rodríguez and Ramírez 2014), while the FBI has been praised as a protocol that can reflect very well the changes in the aquatic system (Kalyoncu and Gulboy 2009). The correlation found with these methods along the urban gradient shows that spider richness is useful in detecting those impacts that were reflected in the scores provided by the FBI’PR and SVAP’PR.
The primary task in biomonitoring is the search for the ideal indicator (Bonada et al. 2006). Our study shows that riparian spiders could be useful indicators, but what remains is to create a tolerance index. One of the limitations when creating a monitoring method is the need to adjust it to local taxonomic richness values for the specific geographic location. Gutiérrez-Fonseca and Ramírez (2016) confronted this problem during the implementation of the BMWP and FBI to Puerto Rico. Due to the low macroinvertebrate richness in Puerto Rico they had to decrease the traditional quality ranges of the protocols in order to obtain representative values of water quality. Therefore, similar adjustments should be considered for a biomonitoring method using riparian spider communities. Similar to aquatic insects some taxa of spiders have global distributions, while other taxa are more restricted to certain geographical regions (v. g., tropical and temperate species). However, by focusing at the family level it should be possible to eliminate some issues of localized genera or endemic species.
The relatively low family richness in our study is most likely due to Puerto Rico being a small oceanic island and naturally tends to have less diversity than large continental regions. In Brazil a maximum of 25 families were found along six 30 m transects (Rodrigues et al. 2014); and in Denmark 16 families were found in forest fragments (Horvath et al. 2014). These studies contrast greatly with the 11 families found in our study, however due to differences in geographic locations and sampling methods it is difficult to accurately compare these findings. A previous study conducted along a forested headwater stream in northeastern Puerto Rico reported a maximum of five spider families (Kelly et al. 2015). However, their study only included web-spinning spiders in their sampling and therefore the additional taxa we found were most likely related to the inclusion of cursorial spiders in our sampling. The use of a simple standardized sampling method will eliminate some of the confounding variables when comparing studies and will be essential in creating an accurate biomonitoring protocol for different geographic regions.
Riparian spider communities respond to impacts in both the riparian and aquatic habitats and represent a more holistic view at the moment of evaluating the condition of stream ecosystems. To the extent of our knowledge this is the first study that has investigated spider community responses to disturbances compared with other well-established stream monitoring protocols. As a result of our findings we strongly recommend the inclusion of riparian spider communities as part of future biomonitoring protocols in order to provide further insight into the impacts of disturbances to stream ecosystems.

