











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkIntroducción
La familia Astroblepidae es un grupo monotípico, representado por el género Astroblepus,Humboldt, 1805, con 81 especies válidas (Eschmeyer y Fong 2017), registrando para Colombia 32 especies, por lo que es considerado el país con el mayor número de especies de esta familia (Schaefer 2003). Este grupo, endémico del Neotrópico, se encuentra distribuido desde Panamá hasta Bolivia a 28° de latitud sur aproximadamente (Schaefer et al. 2011), ocurriendo principalmente en ambientes correntosos.
Morfológicamente, presentan varias modificaciones que les permiten distribuirse en dichos ambientes, como la boca en forma de ventosa y aletas pélvicas móviles (Johnson 1912) que les permiten desplazarse corriente arriba, razón por la cual se les ha llamado comúnmente como “escaladores”. Generalmente, las especies del género habitan sistemas altamente conservados, con aguas limpias y oxigenadas, soportando altas velocidades en el flujo del agua (>130 cm/s) (Miranda-Chumacero 2006). Poseen una distribución altitudinal que se extiende entre los 100 y 4600 m s.n.m., con mayor frecuencia entre los 500 y 2000 m s.n.m. (Schaefer y Arroyave 2010). Las especies de esta familia toleran bajas temperaturas del agua, lo que les ha permitido la colonización y el establecimiento en ambientes de alta montaña (Jacobsen 2008, Miles 1971) y actualmente son considerados un importante bioindicador del estado de conservación de los sistemas en que habitan (Schaefer et al. 2011).
Tal como fue planteado por Schaefer et al. (2011), la identificación morfológica de las especies de Astroblepus no es una tarea fácil, debido a que la mayoría son definidas por un único carácter contrastante o por combinaciones de caracteres externos que presentan altos niveles de variación inter e intraespecífica, llevando en muchas ocasiones a la sobreposición entre especies. Después del trabajo de Regan (1904), no existe, hasta el momento, una revisión taxonómica integral de las especies del género que permita establecer la identidad taxonómica de las especies presentes en Colombia. Muchas de las especies identificadas en listados regionales y nacionales, presentan epítetos de especies descritas originalmente para regiones distantes de otros países; como ejemplo, Astroblepus longifilis (Steindachner, 1882) del Amazonas ecuatoriano, ha sido registrada para las cuencas trasandinas de los ríos Magdalena y Cauca y ríos de la vertiente Pacífica de Colombia. Así mismo, especies como A. homodon (Regan, 1904) y A. longifilis, aparentemente presentan una amplia distribución en el territorio nacional, situación que es necesario evaluar considerando el alto grado de endemismo del grupo.
Un claro ejemplo de esta situación fue identificado en estudios de composición y estructura de la comunidad íctica realizados entre los años 2005 y 2012 para la cuenca del río Porce, donde fueron registradas nueve especies: A. trifasciatus (Eigenmann, 1912), A. unifasciatus (Eigenmann, 1912), A. chapmani (Eigenmann, 1912), A. guentheri (Boulenger, 1887), A. grixalvii (Humboldt 1815), A. supramollis (Pearson, 1937) , A. cyclopus (Humboldt, 1805), A. latidens (Eigenmann, 1918) y A. micrescens (Eigenmann, 1918), en cuanto varios individuos solo fueron identificadas a nivel de género “sp”, y otros fueron reconocidos en las categorías “cf” (comparar con) y “aff” (afin a). De este conjunto, algunas especies tienen localidades tipo en otros países como A. supramollis descrita para la localidad de las Balsas en Perú y A. cyclopus descrita para el río Esmeraldas en Ecuador; así mismo, dentro de las especies registradas para Colombia, algunas de las especies identificadas en el río Porce tienen localidades en regiones distantes como A. trifasciatus y A. unifasciatus descritas para el río Dagua, A. chapmani descrita para la quebrada Boquia en Quindío, A. latidens descrita para la localidad Piperel y distribuida en la cuenca alta del río Meta y A. micrescens, descrita de la quebrada de Agua Larga al norte de Bogotá.
Para intentar identificar las especies distribuidas en la cuenca del río Porce, Hernández et al. (2016) realizaron un análisis de morfometría geométrica usando 12 puntos homólogos de tipo I, II y III; sin embargo, los resultados no fueron estadísticamente significativos (p=0,5194) y no fue posible observar una tendencia de agrupamiento de los datos que correspondiera con la identificación previa de las especies.
Considerando las dificultades en la identificación morfológica de este grupo de especies y la necesidad de esclarecer su identidad taxonómica para monitorear el ensamblaje de sus especies en la región, empleamos la secuencia del gen citocromo oxidasa I (COI) para la identificación genética de los individuos de acuerdo con la metodología propuesta por Hebert et al. 2003; adicionalmente, identificamos caracteres fenotípicos para cada una de las unidades genéticas observadas, principalmente basados en proporciones morfológicas. De acuerdo con los resultados obtenidos proponemos una clave para la identificación de especies del género presentes en la cuenca del río Porce.
Materiales y Métodos
Área de estudio
El río Porce atraviesa el Valle de Aburrá como extensión del río Medellín, iniciando su curso en el alto de San Miguel a 2660 m s.n.m. atravesando la parte central del departamento de Antioquia en dirección noreste (EPM 2004, Jiménez-Segura et al. 2014). Su valle es un cañón profundo en forma de “V” con pendientes pronunciadas y asimétricas normalmente mayores del 50%. La temperatura de esta región es de aproximadamente 21 °C con una precipitación anual de 2248 mm. Los sistemas de primer orden o quebradas afluentes se caracterizan por presentar alta transparencia del agua, elevadas velocidades en el flujo del agua, buena oxigenación, sustrato rocoso, vegetación lateral arbustiva y arbórea, los cuales, en conjunto, constituyen el hábitat de especies del género Astroblepus.
Recolecta de especímenes
Los especímenes fueron capturados con el uso de atarrayas y electropesca en 12 quebradas tributarias al río Porce, cuenca Magdalena-Cauca (Figura 1). Los ejemplares voucher fueron ingresados a la Colección de Ictiología de la Universidad de Antioquia (CIUA) e identificados previamente (Tabla 1). Se obtuvo una muestra de tejido muscular de la base de la aleta dorsal de los individuos, la cual fue preservada en alcohol al 96% y conservada a -20 °C.
Análisis filogenético de la secuencia del gen COI
Para la extracción de ADN se empleó el kit comercial GeneJet Genomic DNA Purification (Thermo Fisher Scientific). La amplificación fue realizada por PCR empleando los cebadores FishF1 (5´-TCAACCAACCACAAAGACATTGGCAC-3´) y FishR1 (5´-TAGACTTCTGGGTGGCCAAAGAATCA-3´) (Ward et al 2005). El volumen final de las reacciones de PCR fue 30 µl, conteniendo 19,88 µl de agua ultrapura, 3,0 µl de Taq Buffer (10x), 4,80 µL de MgCl2 (25 mM), 0,48 µl del mix de dNTPS (2 mM), 0,30 µl de cada cebador (10 mM), 0,24 µl de Taq Polimerasa y 1,0 µl de ADN. El perfil térmico empleado para las amplificaciones fue: desnaturalización inicial por 2 minutos a 95 °C, seguido por 35 ciclos de desnaturalización a 94 °C por 30 segundos, hibridación a 54 °C por 30 segundos y extensión a 72 °C por 1 minuto. Posteriormente se efectuó la extensión final a 72 °C por 10 minutos. Todos los productos de PCR fueron corroborados en gel de agarosa al 1% y posteriormente fueron secuenciados por el método estándar de Sanger en un secuenciador automatizado AB1 en Macrogen (Corea del Sur).
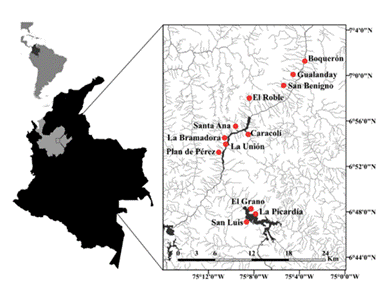
Figura 1 Ubicación geográfica de las 12 quebradas afluentes del río Porce en las cuales fueron capturados los ejemplares de Astroblepus.
Las secuencias obtenidas se editaron y alinearon individualmente usando el programa Clustal W (Thomson et al. 1994) en el programa BIOEDIT 7. 0.5 (Hall 1999). Las secuencias consenso fueron alineadas y se construyó una matriz final, a partir de la cual se estimaron las distancias genéticas entre y dentro de los agrupamientos mediante el modelo de distancia de Kimura de dos parámetros (K2P), en el programa MEGA 5.01 (Tamura et al. 2011). Para los análisis de delimitación de especies, inicialmente estimamos un árbol ultramétrico usando el programa BEAST versión 1.7 (Drummond 2012), bajo el modelo de evolución GTR+G+I. Los análisis se reañizaron con una longitud de 10.000.000 de cadenas MCMC (Cadena de Markov Monte Carlo), seguido por un período de burn-in del 10%, alcanzando un nivel óptimo de convergencia entre el número de iteraciones y el logaritmo de la asignación probabilística. El árbol ultramétrico obtenido se empleó para realizar la delimitación de especies usando el Modelo “Generalized Mixed Yule Coalescent Model” (GMYC) en el programa R versión 3.0.0, usando el paquete “SpLiTS” (Species Limits by Threshold Statistics) y parámetros estándar (intervalo= c(0,10)) que especifican el tiempo de transición entre y dentro de las especies. El uso del modelo GMYC se ha convertido en una aproximación efectiva para la delimitación de especies basadas en datos filogenéticos (Fujisawa y Barraclough 2013, Gerar et al. 2013, Lo et al. 2017). Este método no requiere información previa de las tasas de diversificación, y fue desarrollado específicamente para análisis usando un sólo locus y basado en la predicción que la evolución independiente conduce a la aparición de grupos genéticos distintos (Acinas et al. 2004, Barraclough y Herniou 2003).
Análisis corporal basado en proporciones y conteos
Con la finalidad de observar cuales caracteres morfológicos soportan las agrupaciones establecidas mediante GMYC, se tomaron 13 medidas del cuerpo de los 129 individuos (Tabla 2). Se realizó el conteo de los radios de todas las aletas y la identificación del tipo de dientes tanto de la mandíbula inferior como de la premaxila. Las 13 medidas corporales fueron empleadas para realizar un análisis descriptivo de componentes principales, con la finalidad de encontrar “espacios de dimensiones más pequeñas” detectando qué o cuáles componentes explican la variabilidad entre los ejemplares muestreados; y un análisis discriminante para evaluar la relevancia de dichas variables en la conformación de grupos. Para realizar esto, las medidas se expresaron en razón de la longitud estándar (LE) como sugiere Wilson (1998), se transformaron a log10 y fueron probadas estadísticamente con Kruskall-Wallis aplicando el programa PAST (Hammer et al. 2001).
Resultados
Caracterización genética
Se obtuvieron las secuencias del gen COI con una longitud de 580 pb para los 129 especímenes de Astroblepus. Las secuencias no presentaron inserciones, deleciones o codones de parada. La matriz presentó una composición nucleotídica de 25,7% adenina, 28,8% citosina, 16,6% guanina y 28,9% timina. Se observó un total de 36 haplotipos registrando una diversidad haplotípica h=0,664. Considerando la matriz total, se identificaron 284 sitios conservados, 298 sitios polimórficos y 186 sitios informativos para parsimonia. Los análisis con el modelo GMYC indicaron que el umbral de tiempo antes del cual todos los nodos en el árbol reflejan eventos de diversificación y después del cual todos los nodos en el árbol reflejan eventos coalescentes es de -0,006512577. De acuerdo con lo anterior, el modelo identificó 12 entidades taxonómicas, soportado por un valor de verosimilitud para un modelo nulo de 1212,183 en comparación con la máxima probabilidad para el modelo GMYC de LM=1239,63 (Figura 2). Las distancias genéticas K2P estimadas para los 12 agrupamientos (denotados con letras A-L) corroboran los resultados obtenidos del análisis de GMYC, con valores de 0,08 a 0,41 de distancia genética entre los agrupamientos, con un valor medio de 0,0219 (Tabla 3). En comparación, las distancias intragrupos fueron bajas, con valores entre 0,001 y 0,004.
Tabla 1 Identificación original de los individuos del género Astroblepus empleados en el análisis, su respectivo voucher, lote de colección y la clasificación mediante GYMC (OUT’s).
| Identificación Previa | Voucher | Lote | OUT’s |
|---|---|---|---|
| Astroblepus grixalvii | 7442 | 1944 | E |
| Astroblepus trifasciatus | 7474 | 2020 | H |
| Astroblepus supramollis | 7471 | 2205 | C |
| Astroblepus unifasciatus | 7464 | 2206 | C |
| Astroblepus trifasciatus | 7460 | 2207 | C |
| Astroblepus unifasciatus | 7457 | 2208 | C |
| Astroblepus unifasciatus | 7458 | C | |
| Astroblepus micrescens | 7462 | 2210 | C |
| Astroblepus chapmani | 7475 | 2211 | A |
| Astroblepus supramollis | 7466 | 2212 | C |
| Astroblepus sp.1 | 7463 | 2214 | A |
| Astroblepus trifasciatus | 7465 | 2215 | C |
| Astroblepus cf grixalvii | 7480 | 2218 | H |
| Astroblepus grixalvii | 7477 | 2226 | H |
| Astroblepus chotae | 7476 | 2228 | E |
| Astroblepus supramollis | 7469 | 2229 | C |
| Astroblepus chotae | 7468 | 2230 | E |
| Astroblepus aff latidens | 7461 | 2233 | C |
| Astroblepus trifasciatus | 7472 | 2236 | C |
| Astroblepus sp.1 | 7481 | 2237 | A |
| Astroblepus supramollis | 7528 | 2413 | C |
| Astroblepus grixalvii | 7534 | 2415 | E |
| Astroblepus unifasciatus | 7530 | 2416 | C |
| Astroblepus cyclopus | 7520 | 2417 | A |
| Astroblepus trifasciatus | 7506 | 2419 | C |
| Astroblepus chapmani | 7508 | 2421 | A |
| Astroblepus chapmani | 7510 | A | |
| Astroblepus chapmani | 7509 | A | |
| Astroblepus guentheri | 7517 | 2422 | F |
| Astroblepus supramollis | 7515 | 2425 | C |
| Astroblepus latidens | 7531 | 2426 | A |
| Astroblepus unifasciatus | 7523 | 2429 | C |
| Astroblepus trifasciatus | 7513 | 2432 | C |
| Astroblepus sp.3 | 7524 | 2434 | H |
| Astroblepus supramollis | 7516 | 2435 | C |
| Astroblepus cf latidens | 7522 | 2438 | A |
| Astroblepus cf latidens | 7521 | A | |
| Astroblepus trifasciatus | 7525 | 2441 | C |
| Astroblepus trifasciatus | 7526 | C | |
| Astroblepus trifasciatus | 7497 | 2450 | C |
| Astroblepus grixalvii | 7565 | 2542 | E |
| Astroblepus grixalvii | 7564 | C | |
| Astroblepus sp. | 7573 | 2548 | A |
| Astroblepus sp. | 7572 | A | |
| Astroblepus sp.3 | 7575 | 2549 | H |
| Astroblepus cf latidens | 7576 | 2550 | H |
| Astroblepus unifasciatus | 7577 | 2551 | C |
| Astroblepus unifasciatus | 7578 | C | |
| Astroblepus trifasciatus | 7579 | 2552 | C |
| Astroblepus trifasciatus | 7581 | 2553 | C |
| Astroblepus trifasciatus | 7582 | C | |
| Astroblepus unifasciatus | 7597 | 2610 | A |
| Astroblepus unifasciatus | 7600 | 2613 | C |
| Astroblepus sp.5 | 7967 | 3445 | C |
| Astroblepus trifasciatus | 7944 | 3448 | C |
| Astroblepus trifasciatus | 7945 | C | |
| Astroblepus trifasciatus | 8000 | 3449 | C |
| Astroblepus trifasciatus | 7999 | C | |
| Astroblepus trifasciatus | 7965 | 3450 | C |
| Astroblepus trifasciatus | 7968 | C | |
| Astroblepus trifasciatus | 7975 | 3451 | C |
| Astroblepus trifasciatus | 7976 | L | |
| Astroblepus trifasciatus | 7974 | J | |
| Astroblepus trifasciatus | 7969 | C | |
| Astroblepus trifasciatus | 7978 | C | |
| Astroblepus trifasciatus | 7973 | C | |
| Astroblepus trifasciatus | 7977 | C | |
| Astroblepus grixalvii | 7998 | 3453 | E |
| Astroblepus grixalvii | 7997 | E | |
| Astroblepus grixalvii | 8029 | E | |
| Astroblepus trifasciatus | 7961 | 3456 | C |
| Astroblepus trifasciatus | 7964 | C | |
| Astroblepus trifasciatus | 7960 | C | |
| Astroblepus trifasciatus | 7959 | C | |
| Astroblepus sp.5 | 7947 | 3457 | C |
| Astroblepus sp.5 | 7948 | C | |
| Astroblepus trifasciatus | 8007 | 3458 | C |
| Astroblepus trifasciatus | 8019 | C | |
| Astroblepus trifasciatus | 8004 | D | |
| Astroblepus trifasciatus | 8022 | C | |
| Astroblepus trifasciatus | 8013 | C | |
| Astroblepus trifasciatus | 8005 | C | |
| Astroblepus trifasciatus | 8021 | C | |
| Astroblepus trifasciatus | 8017 | C | |
| Astroblepus trifasciatus | 8024 | C | |
| Astroblepus trifasciatus | 8020 | C | |
| Astroblepus trifasciatus | 8016 | C | |
| Astroblepus trifasciatus | 8018 | C | |
| Astroblepus trifasciatus | 8008 | C | |
| Astroblepus trifasciatus | 8014 | C | |
| Astroblepus trifasciatus | 8023 | C | |
| Astroblepus trifasciatus | 8003 | C | |
| Astroblepus chapmani | 7989 | 3460 | A |
| Astroblepus sp.5 | 7972 | 3466 | I |
| Astroblepus sp.5 | 7979 | C | |
| Astroblepus guentheri | 7956 | 3467 | C |
| Astroblepus guentheri | 7957 | A | |
| Astroblepus guentheri | 7958 | A | |
| Astroblepus guentheri | 7955 | K | |
| Astroblepus sp.1 | 8030 | 3469 | H |
| Astroblepus trifasciatus | 7949 | 3470 | C |
| Astroblepus chapmani | 7971 | 3471 | I |
| Astroblepus sp.1 | 7980 | 3475 | C |
| Astroblepus trifasciatus | 7985 | 3476 | C |
| Astroblepus trifasciatus | 7992 | C | |
| Astroblepus trifasciatus | 7991 | C | |
| Astroblepus trifasciatus | 7993 | C | |
| Astroblepus trifasciatus | 7990 | C | |
| Astroblepus trifasciatus | 7986 | C | |
| Astroblepus guentheri | 8119 | 3491 | E |
| Astroblepus guentheri | 8120 | E | |
| Astroblepus guentheri | 8121 | E | |
| Astroblepus guentheri | 8123 | G | |
| Astroblepus guentheri | 8114 | G | |
| Astroblepus guentheri | 8113 | G | |
| Astroblepus cf unifasciatus | 7922 | 4225 | C |
| Astroblepus sp. | 7929 | 4228 | C |
| Astroblepus cf unifasciatus | 7931 | 4229 | C |
| Astroblepus cf unifasciatus | 7934 | C | |
| Astroblepus cf unifasciatus | 7933 | C | |
| Astroblepus cf unifasciatus | 7930 | C | |
| Astroblepus cf unifasciatus | 7935 | C | |
| Astroblepus cf unifasciatus | 7932 | C | |
| Astroblepus cf unifasciatus | 7926 | 4230 | C |
| Astroblepus cf unifasciatus | 7921 | B | |
| Astroblepus cf unifasciatus | 7925 | C | |
| Astroblepus cf unifasciatus | 7924 | C | |
| Astroblepus cf unifasciatus | 7923 | C |
Tabla 2 Medidas lineales realizadas sobre el cuerpo de ejemplares del género Astroblepus.
| Medidas | Abreviaturas en el texto |
|---|---|
| Longitud estándar | LE |
| Longitud Total | LT |
| Longitud de la cabeza | LC |
| Distancia del extremo posterior de la aleta dorsal a la base anterior del pedúnculo caudal | DDBC |
| Distancia de la base de la aleta dorsal a la espina adiposa | DDPE |
| Distancia de la espina adiposa al pedúnculo caudal | DEPC |
| Longitud predorsal | LPD |
| Longitud prepélvica | LPPEL |
| Longitud preanal | LPA |
| Distancia de la aleta pectoral a la aletas pélvica | DPP |
| Distancia de la aleta pélvica a la aleta anal | DPA |
| Distancia Interorbital | DIO |
| Diámetro del ojo | DO |
Tabla 3 Distancias genéticas interespecíficas obtenidas usando el modelo de Kimura 2 parámetros (K2P) para los 12 grupos de Astroblepus identificados con GYMC.
| A | B | C | D | E | F | G | H | I | J | K | |
| B | 0,13 | ||||||||||
| C | 0,09 | 0,08 | |||||||||
| D | 0,22 | 0,18 | 0,14 | ||||||||
| E | 0,15 | 0,17 | 0,13 | 0,27 | |||||||
| F | 0,19 | 0,22 | 0,18 | 0,31 | 0,15 | ||||||
| G | 0,16 | 0,19 | 0,15 | 0,28 | 0,11 | 0,083 | |||||
| H | 0,17 | 0,19 | 0,16 | 0,28 | 0,10 | 0,114 | 0,081 | ||||
| I | 0,23 | 0,25 | 0,22 | 0,36 | 0,193 | 0,274 | 0,243 | 0,222 | |||
| J | 0,23 | 0,28 | 0,23 | 0,391 | 0,226 | 0,273 | 0,235 | 0,233 | 0,233 | ||
| K | 0,26 | 0,287 | 0,25 | 0,398 | 0,249 | 0,298 | 0,252 | 0,265 | 0,238 | 0,248 | |
| L | 0,25 | 0,39 | 0,26 | 0,41 | 0,25 | 0,30 | 0,257 | 0,256 | 0,265 | 0,29 | 0,274 |
Descripción morfológica de los grupos identificados genéticamente
Cada uno de los agrupamientos identificados en el análisis genético fue caracterizado morfológicamente usando los caracteres mencionados en la Tabla 2 y Tabla 4. Los resultados del análisis estadístico de Kruskall-Wallis no mostraron diferencias significativas entre las variables morfológicas analizadas.
Tabla 4 Estados de carácter para los caracteres y medidas significativas, identificados por morfometría tradicional, para los 12 grupos obtenidos mediante GMYC (los valores de las distancias fueron estandarizados con la longitud estándar de cada organismo).
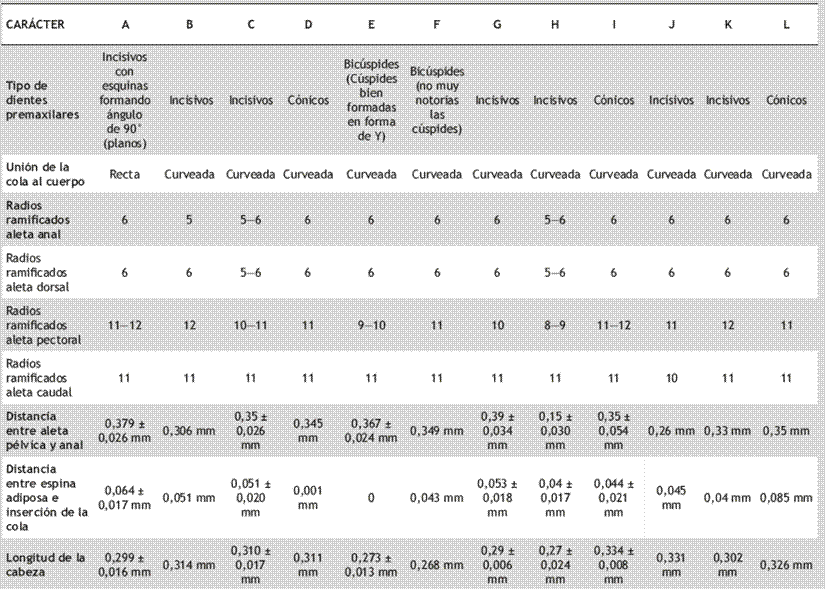
El grupo A presentó los dientes premaxilares incisivos y el perfil dorsal, a la altura del pedúnculo caudal, describió una línea recta. Los individuos incluidos en este grupo presentaron una longitud estándar entre 37,48−57,02 mm, longitud de la cabeza 0,276−0,324 mm, distancia de la base posterior de la aleta dorsal a la base del radio no ramificado de la aleta caudal 0,42−0,46 mm, distancia de la base de la aleta dorsal a la espina adiposa 0,363−0,402 mm, distancia de la espina adiposa a la base del radio no ramificado de la aleta caudal 0,044−0,099 mm, longitud predorsal 0,405−0,458 mm, longitud prepélvica 0, 364−0,434mm, longitud preanal 0,702−0,809 mm, distancia de las aletas pectorales a las pélvicas 0,134−0,246 mm, distancia de la aleta pélvica a la aleta anal 0,317−0,429 mm, distancia interorbital de 0,053−0,068 mm y diámetro del ojo de 0,02 mm. Radios de la aleta dorsal: i, 6, pectoral: i, 11−12, anal: i, 6.
El grupo B estuvo conformado por solo un individuo el cual presentó los dientes premaxilares incisivos y el perfil dorsal, a la altura del pedúnculo caudal, describió una línea curveada. Presentó una longitud estándar de 52,4 mm, longitud de la cabeza 0,314 mm, distancia de la base posterior de la aleta dorsal a la base del radio no ramificado de la aleta caudal 0,42 mm, distancia de la base de la aleta dorsal a la espina adiposa 0,418 mm, distancia de la espina adiposa a la base del radio no ramificado de la aleta caudal 0,051 mm, longitud predorsal 0,405 mm, longitud prepélvica 0,306 mm, longitud preanal 0,72 mm, distancia de las aletas pectorales a las pélvicas 0,162 mm, distancia de la aleta pélvica a la aleta anal 0,408 mm, distancia interorbital de 0,057 mm y diámetro del ojo de 0,02 mm. Radios de la aleta dorsal: i, 6, pectoral: i, 12, anal: i, 5.
El grupo C, con el mayor número de individuos, se caracterizó por un radio blando no ramificado y 10−11 radios blandos ramificados en las aletas pectorales y radio blando no ramificado y 10−11 radios blandos ramificados en la aleta caudal. Los individuos presentaron una longitud estándar entre 21,57−63,37 mm, longitud de la cabeza 0,27−0,35 mm, distancia de la base posterior de la aleta dorsal a la base del radio no ramificado de la aleta caudal 0,37−0,52 mm, distancia de la base de la aleta dorsal a la espina adiposa 0,28−0,45 mm, distancia de la espina adiposa a la base del radio no ramificado de la aleta caudal 0,023−0,131 mm, longitud predorsal 0,42−0,50 mm, longitud prepélvica 0,19−0,79 mm, longitud preanal 0,35−0,85 mm, distancia de las aletas pectorales a las pélvicas 0,11−0,24 mm, distancia de la aleta pélvica a la aleta anal 0,23−0,35 mm, distancia interorbital de 0,050−0,083 mm y diámetro del ojo de 0,019−0,04 mm. Radios de la aleta dorsal: i, 5−6, pectoral: i, 10−11, anal: i, 5−6 y caudal: ii, 10−11, y dientes de la premaxila incisivos.
El grupo D presentó la menor distancia entre la espina de la aleta adiposa y la base del radio no ramificado de la aleta caudal. Presentó una longitud estándar de 42,92 mm, longitud de la cabeza 0,31 mm, distancia de la base posterior de la aleta dorsal a la base del radio no ramificado de la aleta caudal 0,458 mm, distancia de la base de la aleta dorsal a la espina adiposa 0,42 mm, distancia de la espina adiposa a la base del radio no ramificado de la aleta caudal 0,001 mm, longitud predorsal 0,45 mm, longitud prepélvica 0,406 mm, longitud preanal 0,77 mm, distancia de las aletas pectorales a las pélvicas 0,20 mm, distancia de la aleta pélvica a la aleta anal 0,35 mm, distancia interorbital de 0,08 mm y diámetro del ojo de 0,027 mm. Radios de la aleta dorsal: i, 6, pectoral: i, 11, anal: i, 6 y caudal: ii, 11 y los dientes de la premaxila cónicos.
El grupo E no presentó espina en la aleta adiposa y presentó un mayor diámetro del ojo. Los individuos presentaron longitud estándar entre 35,31−103,22 mm, longitud de la cabeza 0,26−0,30 mm, distancia de la base posterior de la aleta dorsal a la base del radio no ramificado de la aleta caudal 0,44−0,51 mm, distancia de la base de la aleta dorsal a la espina adiposa 0,48−0,56 mm, distancia de la espina adiposa a la base del radio no ramificado de la aleta caudal 0,02−0,05 mm, longitud predorsal 0,37−0,43 mm, longitud prepélvica 0,36-0,42 mm, longitud preanal 0,73−0,79 mm, distancia de las aletas pectorales a las pélvicas 0,18−0,22 mm, distancia de la aleta pélvica a la aleta anal 0,32-0,40 mm, distancia interorbital de 0,05-0,07 mm y diámetro del ojo de 0,021-0,039 mm. Radios de la aleta dorsal: i, 6, pectoral: i, 9−10, anal: i, 6 y caudal: ii, 11 y los dientes de la premaxila bicúspides.
El grupo F presentó los dientes de la premaxila bífidos y puntiagudos. Los individuos pertenecientes a este grupo presentaron una longitud estándar entre 35,31-128,4 mm, longitud de la cabeza 0,269-0,294 mm, distancia de la base posterior de la aleta dorsal a la base del radio no ramificado de la aleta caudal 0,44-0,55 mm, distancia de la base de la aleta dorsal a la espina adiposa 0,48-0,58 mm, longitud predorsal 0,36−0,42 mm, longitud prepélvica 0,36-0,42 mm, longitud preanal 0,73-0,79 mm, distancia de las aletas pectorales a las pélvicas 0,19-0,22 mm, distancia de la aleta pélvica a la aleta anal 0,32-0,42 mm, distancia interorbital de 0,056-0,078 mm y diámetro del ojo de 0,02-0,05 mm (lucen “ojones” con respecto a los otros individuos examinados). Radios de la aleta dorsal: i, 6, pectoral: i, 9−10, anal: i, 6 y caudal: ii, 11 y dientes de la premaxila bicúspides.
El grupo H presentó una cabeza pequeña y las aletas dorsal y caudal presentaron un radio blando no ramificado y 5−6 radios blandos ramificados. Los individuos pertenecientes a este grupo presentaron una longitud estándar entre 45,38-53,58 mm, longitud de la cabeza 0,256-0,268 mm, distancia de la base posterior de la aleta dorsal a la base del radio no ramificado de la aleta caudal 0,49-0,51 mm, distancia de la base de la aleta dorsal a la espina adiposa 0,42-0,48 mm, distancia de la espina adiposa a la base del radio no ramificado de la aleta caudal 0,022-0,071 mm, longitud predorsal 0,35−0,38 mm, longitud prepélvica 0,27-0,42 mm, longitud preanal 0,371-0,798 mm, distancia de las aletas pectorales a las pélvicas 0,187-0,212 mm, distancia de la aleta pélvica a la aleta anal 0,316-0,362 mm, distancia interorbital de 0,053-0,064 mm y diámetro del ojo de 0,02−0,03 mm. Radios de la aleta dorsal: i, 5−6, pectoral: i, 8−11, anal: i, 5−6, y dientes de la premaxila incisivos.

Figura 2: Árbol ultramétrico usando secuencias del gen Citocromo Oxidasa subunidad I (COI) para 129 especímenes del género Astroblepus. En la columna A se observan los 12 grupos obtenidos mediante MGYC.
El grupo I presentó la cabeza más grande del grupo de los especímenes analizados. Los individuos pertenecientes a este grupo presentaron una longitud estándar entre 26,49-28,3 mm, longitud de la cabeza 0,33-0,34 mm, distancia de la base posterior de la aleta dorsal a la base del radio no ramificado de la aleta caudal 0,37-0,39 mm, distancia de la base de la aleta dorsal a la espina adiposa 0,31-0,36 mm, distancia de la espina adiposa a la base del radio no ramificado de la aleta caudal 0,05-0,03 mm, longitud predorsal 0,45 mm, longitud prepélvica 0,44-0,45 mm, longitud preanal 0,74-0,80 mm, distancia de las aletas pectorales a las pélvicas 0,16-0,19 mm, distancia de la aleta pélvica a la aleta anal 0,31-0,39 mm, distancia interorbital de 0,06-0,08 mm y diámetro del ojo de 0,02−0,04 mm. Radios de la aleta dorsal: i, 6, pectoral: i, 11−12, anal: i, 6, y dientes de la premaxila cónicos.
El grupo J presentó la menor separación entre las aletas pélvicas y anales. La longitud estándar fue 37,96 mm, longitud de la cabeza 0,33 mm, distancia de la base posterior de la aleta dorsal a la base del radio no ramificado de la aleta caudal 0,44 mm, distancia de la base de la aleta dorsal a la espina adiposa 0,39 mm, distancia de la espina adiposa a la base del radio no ramificado de la aleta caudal 0,045 mm, longitud predorsal 0,47 mm, longitud prepélvica 0,44 mm, longitud preanal 0,70 mm, distancia de las aletas pectorales a las pélvicas 0,21 mm, distancia de la aleta pélvica a la aleta anal 0,26 mm, distancia interorbital de 0,07 mm y diámetro del ojo de 0,03 mm. Radios de la aleta dorsal: i, 6, pectoral: i, 11, anal: i, 6, y dientes de la premaxila incisivos.
El grupo K presentó los dientes de la premaxila incisivos y la aleta pectoral presentó 12 radios blandos ramificados; una longitud estándar de 46,24 mm, longitud de la cabeza 0,30 mm, distancia de la base posterior de la aleta dorsal a la base del radio no ramificado de la aleta caudal 0,41 mm, distancia de la base de la aleta dorsal a la espina adiposa 0,41 mm, distancia de la espina adiposa a la base del radio no ramificado de la aleta caudal 0,039 mm, longitud predorsal 0,42 mm, longitud prepélvica 0,43 mm, longitud preanal 0,75 mm, distancia de las aletas pectorales a las pélvicas 0,19 mm, distancia de la aleta pélvica a la aleta anal 0,33 mm, distancia interorbital de 0,06 mm y diámetro del ojo de 0,02 mm. Radios de la aleta dorsal: i, 6, anal: i, 6.
Finalmente, el grupo L presentó una mayor distancia de la espina adiposa a la base del radio no ramificado dorsal de la aleta caudal, una longitud estándar 36,44 mm, longitud de la cabeza 0,33 mm, distancia de la base posterior de la aleta dorsal a la base del radio no ramificado de la aleta caudal 0,45 mm, distancia de la base de la aleta dorsal a la espina adiposa 0,36 mm, distancia de la espina adiposa a la base del radio no ramificado de la aleta caudal 0,09 mm, longitud predorsal 0,47 mm, longitud prepélvica 0,39 mm, longitud preanal 0,70 mm, distancia de las aletas pectorales a las pélvicas 0,15 mm, distancia de la aleta pélvica a la aleta anal 0,35 mm, distancia interorbital de 0,06 mm y diámetro del ojo de 0,03 mm. Radios de la aleta dorsal: i, 6, anal: i, 6 y los dientes de la premaxila son incisivos.
De acuerdo con los grupos identificados y la agrupación de dos o más individuos, se realizó una clave dicotómica para facilitar la identificación en campo. Todas las medidas fueron estandarizadas respecto la longitud estándar.
1a. Longitud de la cabeza cabe 3,72 a 3,9 veces en LE………………………………...……………………………...…………….…GRUPO H.
1b. Longitud de la cabeza cabe 4 veces o más en LE ………………………………………………………………………………………….……2
2a. Línea dorsal a la altura del pedúnculo caudal describe una línea recta (Figura 3A-B)…………………………………………………...…………...……..……….GRUPO A.
2b. Línea dorsal a la altura del pedúnculo caudal describe un ángulo…………………………………………………….…………………..….…………..3
3a. Aleta adiposa presente, extendiéndose desde la base posterior de la aleta dorsal hasta la base del radio no ramificado de la aleta caudal, sin espina adiposa presente o evidente. (Figura 3C-D)…………………………...……………………………………………………..GRUPO E.
3b. Aleta adiposa presente, reducida o moderada. Espina adiposa presente y evidente………………………………………………………………………..…………….4
4a. Distancia entre la aleta pélvica y la aleta anal cabe de 2,3 a 2,74 veces en LE…………………………………………………………………………...……GRUPO G.
4b. Distancia entre la aleta pélvica y la aleta anal cabe de 2,79 a 3,4 veces en LE…………………………………………………………………………………GRUPO C.

Figura 3: Caracteres diagnósticos para los especímenes de Astroblepus. A. Inserción recta de la cola a la base del radio simple de la aleta caudal. B. Inserción ondulada u oblicua de la cola a la base del radio simple de la aleta caudal. C. Aleta adiposa bien desarrollada sin espina adiposa evidente. D. Aleta adiposa moderada con espina adiposa evidente
Para cada uno de los grupos determinados anteriormente, y debido a que no se contaba con revisión de ejemplares tipo, ni ejemplares de diversas localidades, ni un número representativo de ejemplares por grupo, no se asignó un epíteto específico existente, pero se realizó una identificación a nivel de grupo, considerando las características de los individuos pertenecientes a él, según las claves taxonómicas hasta hoy disponibles, denominándolos así: Grupo A (grupo latidens), Grupo C (grupo trifasciatus), Grupo E (grupo grixalvii), Grupo G (grupo guentheri), Grupo H (grupo micrescens).
Discusión
Los análisis moleculares empleando la región del gen COI han sido utilizados, durante la última década, para la identificación genética de especies y su eficacia se ha validado en varios grupos de animales (Waugh 2007), incluidos los peces (Rosso et al. 2012, Valdez et al. 2009, Ward et al. 2005). Ward et al. (2005) proporcionaron pruebas de la eficacia del ADN “barcoding” en la identificación de peces marinos, en un estudio que examinó más de 200 especies australianas. En general, la identificación de especies a través del código de barras de ADN constituye una poderosa herramienta para la identificación de la diversidad de especies, principalmente en grupos donde es limitada la identificación de caracteres morfológicos únicos, relacionada principalmente con su alta variación y sobreposición (Camacho et al. 2011, Talavera et al. 2013, Tapan et al. 2017). A nivel molecular, la información genética de los individuos revela la variación que ha sido fijada en el genoma y que ha permitido su diversificación.
Una de las principales dificultades en la identificación de especies del género Astroblepus se debe a que la mayoría de sus especies son definidas por un único carácter o por combinaciones de caracteres externos que muestran altos niveles de variación inter e intraespecífica (Schaefer 2011). Claramente, esto puede ser observado al comparar la identificación inicial de los individuos analizados en estudio y los agrupamientos definidos por el análisis de delimitación de especies, siendo que, muchos de los especímenes que conformaban un mismo lote fueron asignados en diferentes grupos. Igualmente, análisis morfométricos previos para el mismo conjunto de individuos identificaron únicamente tres grupos (Hernández et al. 2016). Como se evidencia en revisiones taxonómicas realizadas para el género, muchos de los caracteres usados para definir los límites entre especies de Astroblepus, como la forma del cuerpo, el tamaño del cuerpo y la configuración, así como los patrones de conformación, han sido confundidos con variación a diferentes niveles, que pueden resultar de complejas contribuciones de múltiples fuentes tanto intrínsecas como extrínsecas, tales como la ontogenia, el dimorfismo sexual y la variación geográfica (Miranda-Chumacero 2006), lo que dificulta la delimitación de especies por métodos morfológicos, tal como fue observado durante este estudio. No obstante, considerando la importancia de la identificación morfológica, es indispensable la realización de una revisión taxonómica completa para las especies del género en Colombia, empleando tanto herramientas moleculares, así como explorando otro tipo de caracteres morfológicos (osteológicos, miológicos y merísticos) que permitan una adecuada delimitación de las especies.
De acuerdo con los resultados obtenidos, la diversidad de especies ha sido sobre-estimada y hasta el momento, para el área de estudio, pueden identificarse claramente 12 unidades taxonómicas operacionales (OTU’s), las cuales son soportadas por los valores de distancia genética. A pesar que los límites de diferenciación basados en la distancia genética dependen del conocimiento de las relaciones filogenéticas de un grupo taxonómico, considerando la ausencia de este tipo de información para el grupo de estudio, nuestros análisis de diferenciación genética siguen a Ward (2009), el cual sugiere que valores interespecíficos mayores a 3% y valores intraespecíficos menores al 1% pueden ser límites usados para la identificación de especies; si un espécimen desconocido tiene más de 2% de divergencia, con un nivel de confianza mayor al 95%, puede ser considerado como una especie diferente. No obstante, considerando la escasa información disponible para el grupo, estos resultados deben ser sujetos a posterior corroboración. Actualmente no existe un banco de datos para especies del género distribuidas en Colombia incluido en el Fishbold y solo se han registrado secuencias en el GenBank para especies distribuidas en Perú (Schaefer et al. 2011) lo que limita un análisis comparativo de los resultados obtenidos.
Aunque no se determinaron características únicas que permitieran diferenciar los grupos establecidos, la descripción morfológica de las OTU’s identificadas en este estudio representa una contribución importante en la identificación de los especímenes de las quebradas afluentes al río Porce.
En conclusión, nuestros resultados resaltan la importancia del uso de herramientas moleculares y morfológicas en la identificación de especies que presentan altos niveles de variación intraespecífica.

