











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
La reproducción es el mecanismo mediante el cual una especie genera nueva descendencia con el fin de perpetuarse a sí misma (Vazzoler 1996). Los peces han desarrollado gran diversidad de estrategias y tácticas reproductivas sincronizadas con las condiciones que permitan la fertilización, la incubación y el reclutamiento de la nueva cohorte. En síntesis, la estrategia reproductiva es el patrón general de la reproducción de los individuos de una especie y las tácticas son las variaciones a ese patrón típico y responden a las fluctuaciones ambientales (Wooton 1984). En los sistemas dulceacuícolas tropicales la reproducción de los peces es estacional y se asocia con las lluvias y las crecientes de los ríos. Estas condiciones marcan la maduración gonadal de los individuos maduros y son favorables a la incubación y al desarrollo de las larvas (Araujo-Lima 1994, Jiménez-Segura y López-Casas 2007, Nakatani et al. 2001, Rodríguez y Kossowski 2004, Welcomme 1979a).
El éxito reproductivo puede verse reducido por diversas causas, entre estas se incluyen la disminución del número de adultos por sobrepesca, la pérdida de lugares de desove, la alteración de hábitats para el desarrollo de las larvas, los cambios en el cauce de los ríos y represamientos (Renaud 1997). Estos últimos modifican la continuidad en el flujo unidireccional (Dauble y Geist 2000, Welcomme 1979b), eliminan la conectividad entre sectores del río, generan cambios fisicoquímicos en la masa de agua y alteran los regímenes de flujo aguas abajo del embalse (Ward y Stanford 1995). Estos cambios en conjunto modifican las condiciones óptimas para la reproducción, puesto que los peces sincronizan su estrategia reproductiva con la estacionalidad de las lluvias y en las crecientes (Gomes y Agostinho 1997).
El concepto de ensamblaje usado en este artículo está fundamentado en Magurran (2004), quien afirma que el ensamblaje se define como una comunidad en la cual las especies que la componen están relacionadas filogenéticamente; por otro lado, la dinámica reproductiva se utilizará para hacer referencia a los cambios a lo largo del tiempo de la intensidad y la actividad reproductiva del ensamblaje de peces. El ensamblaje del área de influencia de los embalses Porce II y Porce III está compuesto por 48 especies (EPM 2016), sin embargo, para los análisis de este estudio solo se consideraron aquellas especies que tuvieron mayor frecuencia de ocurrencia.
El objetivo de este trabajo fue estudiar la dinámica reproductiva del ensamblaje de peces del área de influencia de los embalses Porce II y Porce III, dentro de la cuenca media del río Porce.
Materiales y Métodos
Área de estudio
La cuenca media del río Porce pertenece a la cuenca Magdalena-Cauca (Colombia) y presenta un régimen de lluvias esencialmente bimodal, con dos períodos con altas precipitaciones, el primero entre abril-mayo y el segundo entre septiembre-octubre (Silva 2008) (Figura 1).
La temperatura atmosférica media anual es de 21 °C, la humedad relativa es aproximadamente del 80% (EPM y UdeA 2012). La geomorfología del valle del río Porce está desarrollada en un cañón profundo en “V” con laderas amplias y pendientes asimétricas. Esta característica del río Porce permitió la construcción de los embalses que se describen a continuación.
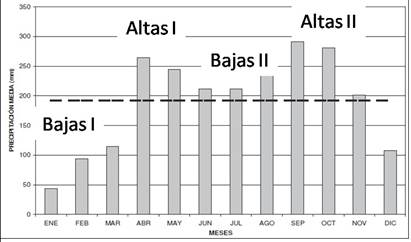
Figura 1 Histograma de precipitación media mensual de una estación pluviométrica representativa de la zona de estudio.
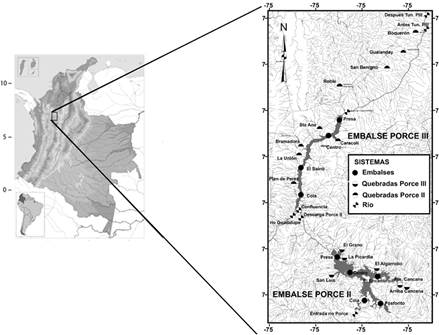
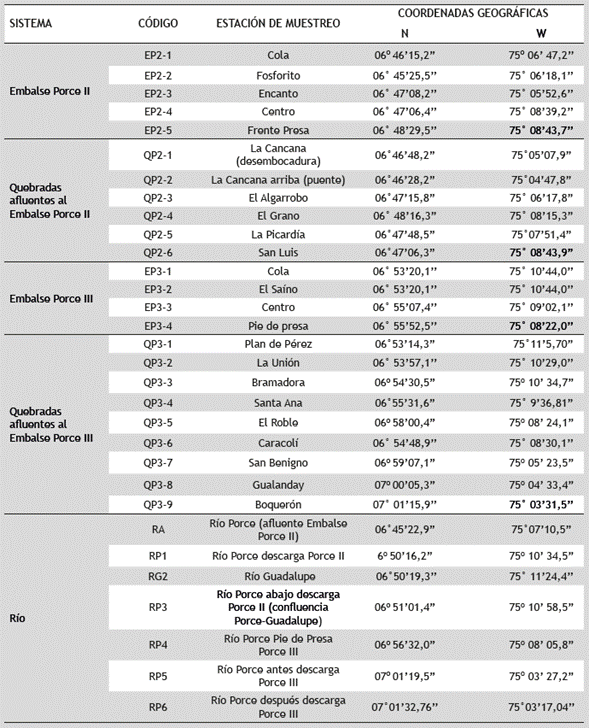
El embalse Porce II entró en su fase de funcionamiento en el año 2002; está localizado entre 06° 48' 29.5'' N y 75°08'43.7'' W, tiene un área de 13’780.000 m2, un rango de altitud de 936-939 m y un tiempo de retención de 30 días. El embalse Porce III inició operaciones a finales del año 2010; está ubicado entre 06° 55' 52.5'' N y 75° 08' 22.0''W, con un área de 4’610.000 m2, un rango de altitud de 512-694 m y el tiempo de retención es de 8 días. Estos embalses fueron construidos en cascada y están ubicados en la cuenca media del río Porce, al nordeste del departamento de Antioquia, en jurisdicción de los municipios de Gómez Plata, Guadalupe, Amalfi y Anorí (Tabla 1, Figura 2). La función de estos dos embalses es la generación de energía eléctrica (EPM y UdeA 2012).
Diseño muestral
Entre los años 2011 y 2016 se realizaron en total 13 jornadas de campo, correspondiendo siete de estas a los periodos de altas lluvias y el resto a los momentos de bajas lluvias. A pesar de la temporalidad climática definida por la distribución de lluvias, el nivel de los embalses dependió de la generación de energía y no necesariamente de los aportes del río Porce y de las quebradas.
En cada campaña de campo se obtuvieron muestras de la ictiofauna presente en 31 estaciones. Estos sitios se distribuyeron en tres ambientes principales: embalses en el cauce del río Porce, en quebradas tributarias a los embalses y en ríos afluentes o efluentes a estos embalses; así mismo, estos ambientes dan lugar a cinco sistemas, embalse Porce II, embalse Porce III, Quebradas Porce II, Quebradas Porce III y río Porce (Tabla 1, Figura 2).
Captura de peces
Para capturar los peces en el embalse se usaron redes de espera con diferentes ojos de malla (1, 2, 3, 4, 5, 6, 7, 8, 9, 10), mientras que en los sistemas de río y quebradas se emplearon atarrayas y electropesca. Las capturas fueron realizadas generalmente en horas del día (7:00-14:00 horas). Cada ejemplar capturado fue fotografiado, medido en su longitud estándar y pesado, y posteriormente anestesiado con aceite de clavo, el cual es un sedante comúnmente usado en peces (Neiffer et al. 2009); cuando el anestésico hizo su efecto, el pez fue fijado en solución de formalina al 10% (Murphy y Willis 1996) y se le asignó un número de campo. Posteriormente, el espécimen fue almacenado para su procesamiento en el laboratorio de la Universidad de Antioquia. Las consideraciones éticas que se tuvieron en cuenta para el desarrollo de este trabajo de investigación son las descritas por Murphy y Willis (1996).
Métodos en Laboratorio
Cada ejemplar traído del campo al laboratorio fue pesado (peso total y eviscerado), medido en su longitud estándar e inmediatamente se le realizó un corte uroventral para exponer su cavidad visceral; el contenido fue removido y sus partes fueron identificadas. Las gónadas se removieron cuidadosamente y se describió su morfología determinando el sexo, el estado de madurez y registrando su peso
Las escalas de madurez basadas en la información macroscópica siguieron la propuesta de Vazzoler (1996) la cual define cinco estadios: A= inmaduro; B = en maduración; C = maduro; D = desovado; E = reposo.
Análisis de la información
Para definir los periodos climáticos de mayor actividad reproductiva del ensamblaje, se relativizó el número de individuos maduros (estadio C + D) teniendo en cuenta el total de individuos analizados de cada taxón, por periodo climático en cada sistema.
Para determinar los sistemas donde el ensamblaje se reproduce y en qué temporadas climáticas, se utilizaron dos indicadores de la reproducción de los peces: el primero es el de actividad reproductiva en términos del porcentaje de individuos maduros de los taxones y el segundo es el de Intensidad Reproductiva (IR).
Para determinar las categorías y los valores de la IR del ensamblaje se empleó la propuesta de Vazzoler y Agostinho (1991) descrita en Vazzoler (1996). Esta propuesta se fundamenta en la asociación de la frecuencia de hembras maduras de cada especie (estadios C y D) y el porcentaje que la relación gonadosomática promedio (RGSpromedio) representa de la RGSmáxima, en cada sistema y momento climático.
La ecuación de la intensidad reproductiva (IR) utilizada fue la siguiente:
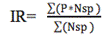
donde:
Nsp=número de especies en las que estuvo presente una categoría en un periodo de muestreo particular.
P=representa una categoría de IR que puede ser:
P=4 desove masivo
P=2 desove ocasional
P=1 maduración incipiente
P=0 sin actividad reproductiva
La RGSpromedio que se utilizó en los análisis fue el resultado de la diferencia entre la RGS corregida (RGS2) y la RGS (RGS1). RGS1=WG/WT; RGS2=WG/(WT-WG) en donde, WG=peso de la gónada, WT=peso total del pez (Vazzoler 1996).
Para probar si hay dependencia o asociación de la IR del ensamblaje de cada sistema acuático con los periodos climáticos, se hizo una prueba estadística de correlación de Spearman, ya que, en esta correlación se comparan variables cualitativas y cuantitativas; además, para aplicar esta prueba no es necesario hacer hipótesis sobre la distribución de los datos. Para demostrar si había diferencias en la IR entre los cinco sistemas, se aplicó la prueba de Kruskal Wallis, dado que, previamente a través de las pruebas de Kolmogorov-Smirnov y Shapiro-Wilk, se comprobó que los datos no presentaban distribución normal. Adicionalmente, como complemento a la prueba de Kruskal Wallis, se realizó la prueba de homogeneidad por pares de Smirnov Kolmogorov, considerando que este análisis tiene la ventaja de medir diferencias no solo en la tendencia central sino también en la dispersión y la distribución de los datos. Todas las pruebas estadísticas se realizaron con el software Statistica 13.0.
Para los análisis de este estudio, el ensamblaje estuvo compuesto por las especies de mayor ocurrencia en los cinco sistemas definidos en el área de influencia de los embalses Porce II y Porce III (Tabla 2).
Tabla 2 Especies de mayor ocurrencia del ensamblaje de peces del área de influencia de los embalses de Porce II y Porce III
| Familia | Nombre científico | Sistema | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Embalse Porce II | Embalse Porce III | Quebradas Porce II | Quebradas Porce III | Río Porce | ||||||||
| Chraracidae | Astyanax microlepis | x | x | x | x | |||||||
| Hemibrycon sp. | x | x | x | |||||||||
| Bryconamericus caucanus | x | x | x | |||||||||
| Roeboides dayi | x | x | x | |||||||||
| Bryconidae | Brycon henni | x | x | x | x | x | ||||||
| Parodontidae | Parodon magdalenensis | x | ||||||||||
| Chrenuchidae | Characidium phoxocephalum | x | ||||||||||
| Poecilidae | Poecilia caucana | x | x | |||||||||
| Cichlidae | Andinoacara latifrons | x | x | x | x | |||||||
| Coptodon rendalli | x | x | ||||||||||
| Oreochromis niloticus | x | x | ||||||||||
| Parachromis loisellei | x | x | ||||||||||
| Astroblepidae | Astroblepus trifasciatus | x | x | |||||||||
| Astroblepus unifasciatus | x | |||||||||||
| Loricariidae | Chaetostoma thomsoni | x | x | x | ||||||||
| Callicthydae | Hoplosternum magdalenae | x | ||||||||||
Resultados
Intensidad reproductiva del ensamblaje
Los sistemas de embalses presentaron altas intensidades reproductivas (IR) durante casi todos los periodos climáticos muestreados, con valores entre 2,8 y 4; sin embargo, los valores más bajos de este indicador se observaron en Porce II Lluvias II-2012 (IR=1) y Porce III Estiaje II - 2016 (IR=2) (Figura 2). En contraste, en el río y quebradas tributarias a Porce II y Porce III, se observó una mayor IR en los periodos de estiaje, aunque los valores en los periodos de lluvias también fueron, en su mayoría, altos (Figura 3 y Figura 4).
Actividad reproductiva de los taxones
La familia Characidae presentó el mayor porcentaje de individuos maduros en la mayoría de los periodos climáticos muestreados, en los embalses, en el río y en las quebradas tributarias a Porce III. En el embalse Porce II, entre los años 2011 y 2014, la familia Characidae presentó los mayores porcentajes de individuos maduros seguida de las familias Callychthydae y Cichlidae; en los años 2015 y 2016, las familias con mayor actividad reproductiva fueron Characidae, Cichlidae y Bryconidae, aunque los análisis de esta última familia estuvieron basados en un bajo número de individuos. En el embalse Porce III, las familias Characidae y Cichlidae, seguida de Bryconidae, aportaron en este orden al porcentaje de individuos maduros. A partir del año 2014 no se volvieron a registrar capturas de la familia Callychthydae (Figura 5 y Figura 6).
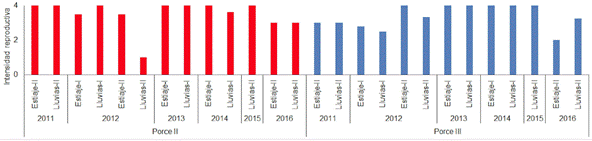
Figura 3 Intensidad Reproductiva (IR) de la comunidad de peces en los embalses Porce II y Porce III entre los años 2011 y 2016.
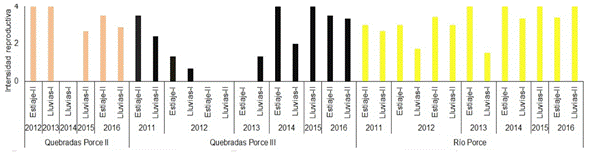
Figura 4 Intensidad Reproductiva (IR) de la comunidad de peces en las quebradas tributarias al embalse Porce II y Porce II y en el río Porce entre los años 2011 y 2016
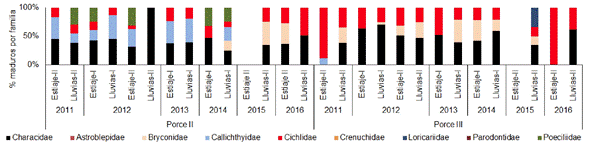
Figura 5 Actividad reproductiva (% de individuos maduros) de cada una de las familias que conforman el ensamblaje de peces en los embalses Porce II y Porce III entre los años 2011 y 2016.
En las quebradas tributarias de Porce II, la familia Characidae presentó individuos maduros en la mayoría de los periodos, sin embargo, sus porcentajes solo fueron dominantes en dos de estos (lluvias 2014 y lluvias 2015); para los monitoreos correspondientes al año 2016, los porcentajes de maduros estuvo distribuido entre cinco familias con bajo dominio de alguna de estas. En contraste, en las quebradas que tributan al embalse Porce III, la mayor actividad reproductiva estuvo representada principalmente por las familias Characidae, Bryconidae y, en menor porcentaje, por Astroblepidae. En el sistema de río, las familias Characidae y Poecilidae presentaron altos porcentaje de individuos maduros en la mayoría de los periodos climáticos y, al igual que para el resto de las familias, no se observó alguna tendencia relacionada con la temporalidad pluviométrica (Figura 6).
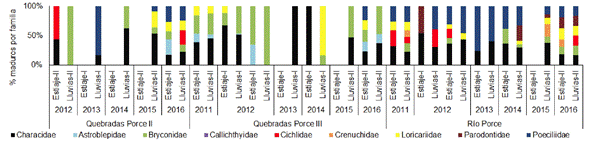
Figura 6 Actividad reproductiva (% de individuos maduros) de cada una de las familias que conforman el ensamblaje de peces en las quebradas tributarias a los embalses Porce II y Porce II y en el río Porce entre los años 2011 y 2016.
Las especies del embalse Porce II registraron en su mayoría altos porcentajes de individuos maduros en todos los periodos climáticos. La especie Andinoacara latifrons (Steindachner 1878) mostró porcentajes de individuos maduros cercanos al 25% hasta el año 2012, y a partir de ese año sus porcentajes estuvieron en su mayoría por encima del 50%. Los taxones Astyanax microlepis Eigenmann 1913, Parachromis loisellei (Bussing 1989), Brycon henni Eigenmann 1913, Roeboides dayi (Steindachner 1878) y Hoplosternum magdalenae Eigenmann 1913, presentaron porcentajes de individuos maduros cercanos o iguales al 100%. A partir del año 2014, la especie R. dayi se capturó de manera ocasional y H. magdalenae no se volvió a capturar. En general, no se observó sincronía de la proporción de individuos maduros y los periodos climáticos (Figura 7).
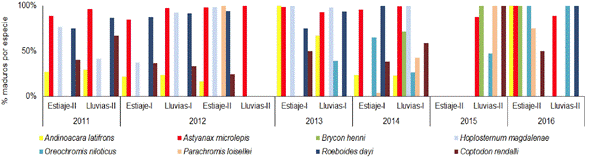
Figura 7 Porcentaje de individuos maduros (estadios C y D) de cada una de las especies de peces en el embalse Porce II en diferentes periodos climáticos entre los años 2011 y 2016.
En el embalse Porce III las especies A. microlepis, R. dayi y Coptodon rendalli (Boulenger 1897) presentaron porcentajes de maduros superiores al 85% en la mayoría de los periodos muestreados. La especie B. henni contó con mayores porcentajes de individuos maduros en los periodos de lluvias entre los años 2011 a 2013. Para el año 2016, Oreochromis niloticus (Linnaeus 1758), C. rendalli, A. latifrons, P. loisellei y A. microlepis mostraron actividad reproductiva alta, ya que registraron porcentajes de individuos maduros por encima del 98% (Figura 8).
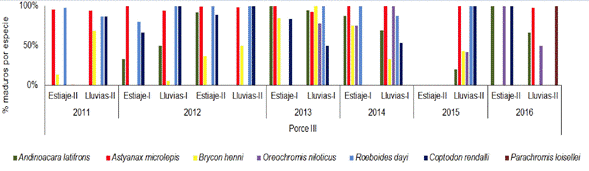
Figura 8 Porcentaje de individuos maduros (estadios C y D) de cada una de las especies de peces en el embalse Porce III en diferentes periodos climáticos entre los años 2011 y 2016.
En las quebradas tributarias del embalse de Porce II, las especies A. microlepis, P. caucana y A. latifrons presentaron porcentajes de maduros superiores al 64% en la mayoría de los periodos que registraron madurez gonadal. En los años 2013 y 2014, B. henni y Bryconamericus caucanus Eigenmann 1913 mostraron actividad reproductiva en el periodo correspondiente a lluvias I. Para el año 2016 se observó, particularmente, que los mayores porcentajes de maduración se presentaron en el periodo pluviométrico de las lluvias (Figura 9).
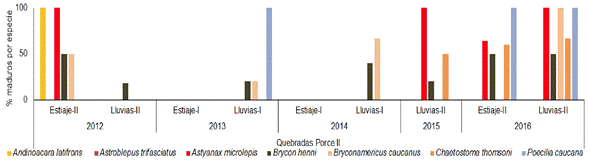
Figura 9 Porcentaje de individuos maduros (estadios C y D) de cada una de las especies de peces presentes en quebradas tributarias a Porce II entre los años 2012 y 2016.
En quebradas Porce III se observó que B. henni tuvo un mayor porcentaje de individuos maduros en la época de lluvias durante todos los años muestreados 2011-2016. Algo similar se evidenció con B. caucanus que mostró sincronía de su reproducción con la época de lluvias en los años 2011 a 2013 y 2015, aunque para el primer semestre del año 2014 mostró mayor actividad reproductiva en la época de estiaje; no obstante, en las lluvias de ese año no se realizaron análisis reproductivos puesto que el número de individuos capturados fue muy bajo. En el año 2016, B. henni y Hemibrycon sp, presentaron altos porcentajes de individuos maduros en los dos periodos climáticos (Figura 10).
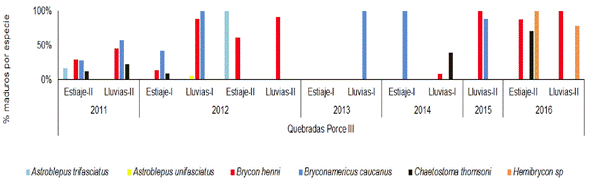
Figura 10 Porcentaje de individuos maduros (estadios C y D) de cada una de las especies de peces presentes en quebradas tributarias a Porce III entre los años 2011 y 2016.
En general, para el sistema de río se encontró que A. microlepis, B. caucanus y Poecilia caucana (Steindachner 1880) presentaron altos porcentajes de individuos maduros en la mayoría de los periodos climáticos muestreados. En el año 2015, A. microlepis fue la única especie que presentó altos porcentajes de individuos maduros. Para el año 2016, nueve especies analizadas mostraron la totalidad de individuos maduros en los periodos que fueron capturados; en el caso de B. henni, registrada en ambos periodos, si bien los análisis fueron realizados con pocos individuos se determinó que su actividad fue más alta en lluvias (Figura 11).
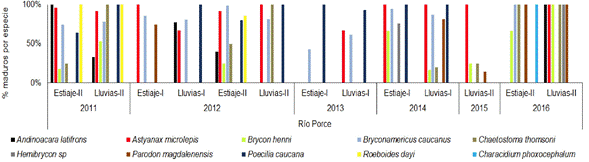
Figura 11 Porcentaje de individuos maduros (estadios C y D) de cada una de las especies de peces presentes en el río Porce entre los años 2011 y 2016.
De acuerdo con los resultados de la prueba estadística del coeficiente de correlación de Spearman, no hubo correlación significativa (p>0,05) entre la IR y los periodos climáticos en ninguno de los sistemas analizados (embalses, quebradas tributarias y río). Además, los resultados de la Anova de Kruskal Wallis y la prueba de la mediana indicaron que no hubo diferencias significativas (p>0,05) en la IR entre los cinco sistemas analizados; sin embargo, al comparar por pares a través del test de Smirnov Kolmogorov, se encontró que la IR del sistema de Quebradas de Porce III, presentó diferencias significativas (p<0,05) con los embalses de Porce II y Porce III.
Discusión
La intensidad reproductiva (IR) del ensamblaje de peces en los embalses de Porce II y Porce III fue alta en la mayoría de los periodos climáticos, al igual que el porcentaje de individuos maduros de la mayoría de las especies. Con excepción de la especie B. henni, estos resultados son coherentes con Vazzoler (1996) quien afirma que aquellas especies que permanecen en los embalses se reproducen permanentemente en ellos y cumplen todas las fases de su ciclo de vida en estos ambientes. La reproducción permanente en estos sistemas acuáticos es explicada por Godinho (2010) quien afirma que los embalses tienen disponibilidad de alimento y altos niveles de agua, y no están restringidos a los periodos de lluvias. Por lo tanto, los peces pueden reproducirse durante todo el año debido a que hay condiciones favorables de manera permanente para su descendencia.
Los mayores valores de IR en el río y las quebradas tributarias a los embalses fueron detectados en los periodos de estiaje, sin embargo, esto no quiere decir que la reproducción del ensamblaje ocurra solo en ese periodo ya que en la mayoría de los muestreos correspondientes a lluvias se registran valores que están por encima de los desoves ocasionales, siendo algunos iguales o cercanos a los desoves masivos. Estos resultados difieren a lo encontrado por Agostinho et al. (2004), Braga (2001), Gomiero y Braga (2005) y Rindonelli y Braga (2010) en ríos de Brasil, donde el ensamblaje de peces presentó mayor intensidad reproductiva en el periodo de lluvias; no obstante, para este estudio, en el periodo de lluvias también se registraron desoves masivos, aunque no fue con la misma regularidad o magnitud del periodo de estiaje.
El resultado de la prueba de Spearman indica que no hay correlación significativa entre la reproducción y los periodos climáticos en ninguno de los sistemas acuáticos estudiados y, por tanto, en términos estadísticos, la intensidad reproductiva del ensamblaje de peces es independiente al estiaje o las lluvias. La prueba por pares de Smirnov Kolmogorov, demostró que las quebradas tributarias a Porce III tienen una intensidad reproductiva que es particular a los dos embalses, esto puede deberse a que dentro de los tres sistemas loticos este conjunto de quebradas son más representativas de un ambiente natural, ya que, el sistema de río Porce tiene un caudal que está influenciado por la generación de electricidad, y en el caso de las quebradas tributarias de Porce II, estas tienen un mayor grado de intervención antrópica; la condición singular de las Quebradas de Porce III, concuerda con Munro et al. (1990) quienes plantean que los ríos y quebradas naturales son afectados por cambios estacionales asociados con la llegada y retirada de las lluvias. Es importante aclarar que cuando se hacen análisis de actividad reproductiva a nivel de especies, se determinó que si hay especies con una mayor reproducción durante las lluvias, como es el caso de B. henni y con menor tendencia algunas otras especies pertenecientes a la familia Characidae.
Aunque la temporalidad del desove es una estrategia que tiene como finalidad hacer coincidir la presencia de larvas y juveniles con adecuadas condiciones ambientales y de disponibilidad de recursos (Criscuolo-Urbinati 2005), es posible que en el embalse Porce II y Porce III se mantengan unas condiciones adecuadas para la reproducción durante las lluvias y los estiajes. Además, según Kramer (1978) en un ambiente menos estacional, los peces del trópico pueden tener temporadas extendidas de reproducción. Por otro lado, Winemiller (1996) afirma que la reproducción puede estar influenciada por la disponibilidad de espacio y otros recursos, como competición, depredación y otras interacciones bióticas.
En general, el ensamblaje de peces perteneciente al río y a las quebradas tributarias de los embalses de Porce II y Porce III se reproduce tanto en las lluvias como en el estiaje, siendo más intensa la reproducción en este último periodo. Este comportamiento puede ser explicado según Winemiller (1989) por las diferencias entre las especies que componen el ensamblaje de un mismo río, que pueden diferir en la sincronía y la duración de la reproducción.
Es importante resaltar que en el embalse Porce III y sus quebradas tributarias, B. henni presenta mayor porcentaje de individuos maduros en los periodos de lluvias, lo cual coincide con Usma (2001) quien afirma que, de acuerdo con el sitio, la reproducción de esta especie puede ocurrir en diferentes épocas del año, asociada en su mayoría al periodo de lluvias; Mancera-Rodríguez (2017) también encontró, en este mismo periodo climático, un mayor desarrollo gonadal y un alto porcentaje de individuos maduros de B. henni en las quebradas afluentes a los ríos Nare y Guatapé.
El comportamiento reproductivo de B. henni concuerda con lo expuesto por Kerguelen- Durango y Atencio-García (2015) quienes afirman que la selección del momento de reproducción de los peces en los sistemas acuáticos naturales andinos, está determinada por los patrones hidrológicos y el transporte de nutrientes asociado con el periodo de lluvias. Debido a la recurrencia de B. henni en el embalse Porce III, es importante anotar que, aunque este embalse es un sistema acuático artificial, presenta características hidráulicas de un río lento, debido a su bajo tiempo de retención de tan solo ocho días.
La familia Characidae presenta mayor porcentaje de individuos maduros en el área de influencia de los embalses Porce II y Porce III. Astyanax microlepis y R. dayi pertenecen a esta familia y son las especies más abundantes de la zona, superando el 80% de individuos maduros en la mayoría de capturas; sin embargo, en los años 2015 y 2016, la captura de R. dayi en el sistema de embalses descendió considerablemente. Según Loaiza-Santana y Jiménez-Segura (2013), considerando la categorización de Winemiller (1989), las poblaciones de estas dos especies en los embalses del río Porce exhiben estrategia reproductiva tipo oportunista, caracterizada por un esfuerzo reproductivo alto y sostenido.
Aunque dentro de los análisis solo se tuvo en cuenta como pez migrador a B. henni, es importante aclarar que durante las tres jornadas de rescate peces en el túnel de fuga de la central hidroeléctrica Porce III, realizadas en los años 2012 y 2015, se rescataron y reubicaron en el río Porce especies migratorias, como, Prochilodus magdalenae Steindachner 1879, Ichthyoelephas longirostris (Steindachner 1879) y Pimelodus blochii Valenciennes 1840; en uno de los rescates del año 2015, el 80% de los individuos rescatados pertenecían a P. magdalenae, por lo tanto, es altamente probable que para ese momento estuviera activa la migración reproductiva de esa especie por el río Porce (Jiménez-Segura et al. 2018).
El estudio de la reproducción se planteó a la escala de ensamblaje para este artículo, considerando que su objetivo fue estudiar la dinámica reproductiva de los peces, buscando algún tipo de patrón reproductivo característico de cada uno de los tres ambientes analizados (embalses, quebradas tributarias y río).
Los porcentajes de individuos maduros de las especies se ajustan, en cierta medida, a los resultados obtenidos a través de la IR, sin embargo, se debe tener en cuenta que el ensamblaje está compuesto por diferentes especies que pueden tener diferentes estrategias reproductivas no detectables analizando la reproducción a nivel de ensamblaje.
En general, los peces de los ambientes de embalses tienen altas intensidades reproductivas durante todos los periodos climáticos, presentando valores bajos en ciertos periodos, pero sin seguir algún tipo de tendencia relacionada con las lluvias.
Los sistemas loticos de quebradas tributarias a los embalses y río Porce tienen su mayor IR en el periodo de estiaje.
Entre los cinco sistemas analizados, el sistema lotico de quebradas de Porce III presenta un comportamiento, en su intensidad reproductiva, diferente estadísticamente al ambiente lenítico de los embalses de Porce II y Porce III.
Cuando se analiza la actividad reproductiva por especie, en los sistemas de Quebradas tributarias a los embalses y el río Porce, hay algunas especies de peces que tienen una sincronía de su reproducción con los periodos de lluvias.

