











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkINTRODUCCIÓN
TerpsinoëEhrenberg 1843, es un género de diatomea céntrica perteneciente a la familia Biddulphiaceae (Bacillariophyta), morfológicamente caracterizado por tener valvas elípticas, con márgenes ondulados, barras transversales prominentes, un rimoportula de posición central y un pseudocelo en cada ápice (Round et al. 1990, Sterrenburg 1994). Es un género con un número reducido de especies, en general citado para ambientes continentales tropicales a subtropicales, aunque también se registra en ambientes marino costeros (Carmona-Jiménez et al. 2017, Round et al. 1990, Tuji 2018).
Terpsinoë musicaEhrenberg (1843) es la especie tipo del género y debe su nombre a que las barras transversales rectas se hallan engrosadas hacia el centro, concediéndole un aspecto semejante a las notas musicales cuando los frústulos se observan en vista conectival. Las células vivas están unidas entre sí por un mucílago secretado por los pseudocelos formando colonias en zig-zag (Boyer 1900, Carmona-Jiménez et al. 2017, Sterrenburg 1994). En su descripción original Ehrenberg mencionó que fue hallada en materiales colectados en el municipio de Atotonilco El Grande (México), sin incluir referencia alguna respecto al sitio de recolección. Posteriormente se registró en diversos sistemas continentales y marinos costeros de América (Carmona-Jiménez et al. 2017, Luttenton et al. 1986, Sterrenburg 1994, Vouilloud 2003), Europa (Peragallo y Peragallo 1897, Van Heurck 1899), África (Baudrimont 1973, El-Awamri et al. 2003), e islas del Pacífico (Ehrenberg 1843, Navarro y Lobban 2009). Por registrarse en ambientes tan diversos es considerada una especie eurihalina (Baudrimont 1973, Navarro y Lobban 2009, Van Heurck 1899), que prolifera con mayor frecuencia en aguas duras con temperaturas cálidas (Sterrenburg 1994). Puede colonizar una gran diversidad de sustratos, ya que se ha encontrado sobre rocas (El-Awamri et al. 2003), en raíces de árboles sumergidos y briófitas acuáticas (Luttenton et al. 1986), posada en otras algas filamentosas como Cladophora (El-Awamri et al. 2007) y Bostrychia (Azevedo 1999) y también formando parte del perifiton de los ríos (Carmona-Jiménez et al. 2017).
La recopilación de los trabajos taxonómicos realizada por Montoya et al. (2013) y la revisión de trabajos posteriores publicados para Colombia, mostraron que no existe ningún registro previo de alguna especie de este género para el país.
MATERIALES Y MÉTODOS
Este trabajo forma parte de una investigación que abarcó 18 sistemas lóticos de las zonas biogeográficas de los Andes (en el departamento de Santander), Caribe (en el departamento de Magdalena), Orinoquía (en el departamento de Arauca) y Pacífico Colombiano (en el departamento de Nariño) (Figura 1). El material analizado en este estudio se recolectó en dos sistemas lóticos de la región de la Orinoquía (Río Lipa confluencia con el arroyo Primores y Río Lipa aguas arriba; estos sitios se ubicaron a 134 m s. n. m. y geográficamente entre los 6º 47’ 6,7” N y 71º 1’ 34” W, y 6º 49’ 33,2” N y 71º 2’ 16,1” W, respectivamente) y en dos del Caribe (en el municipio de Ciénaga, departamento de Magdalena, en los arroyos Canta Rana (910 m s. n. m., 11º 0’ 13,9’’ N y 74º 3’ 46,8’’ W) y El Padre (928 m s. n. m., 10º 58’ 36,1’’ N y 74º 4’ 22,7’’ W) (Figura 1).
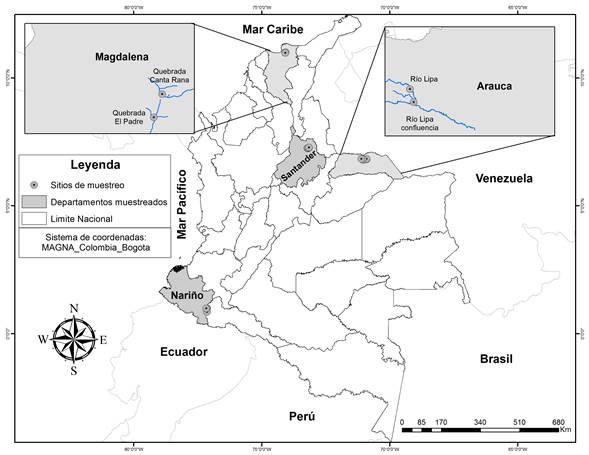
Figura 1 Ubicación geográfica de los sitios de muestreo en las cuatro regiones biogeográficas estudiadas. Regiones donde se registró Terpsinoë musica: Región Caribe: departamento del Magdalena y Región Orinoquía: departamento de Arauca.
En campo, las muestras del perifiton se recolectaron sobre sustratos rocoso y macrófitas (10 sustratos en cada sitio), cepillando una superficie de 90 cm2 por sitio. Cada muestra se fijó con Lugol (al 10%) y 6 a 8 gotas de formalina. Adicionalmente se midió la elevación sobre el nivel del mar con un GPS (Garmin etrex 20x), el caudal por medio del método de flotación (ISO 1998), la temperatura con un termómetro de mercurio, el pH con un pH-metro Martini (pH56 - 0,01 pH), la conductividad eléctrica con un medidor de conductividad Martini (Conductividad 0-3.999 μS/cm) y el oxígeno disuelto por medio de titulación con Winkler (Merck) (Tabla 1).
En el laboratorio las muestras se trataron con agentes oxidantes para eliminar la materia orgánica siguiendo el método descrito en CENT/TC 230 (2002). Para observar las muestras en el microscopio óptico (MO) el material tratado se montó en Naphrax®, y para el microscopio electrónico de barrido (MEB), se montó sobre tacos de vidrio y se recubrió con una capa fina de oro paladio en un metalizador Jeol JFC-1100 (fine coat ion sputter). Las observaciones se realizaron con un microscopio 500 LM Leica DM y en un microscopio electrónico de barrido Jeol JSM-6490LV MEB de la Universidad de Antioquia, Colombia. Las muestras fueron depositadas en el Herbario de la División Ficología del Museo de La Plata (LPC). En la Tabla 1 se alistan las muestras empleadas y los números de la colección.
Tabla 1 Valores de las variables físicas y químicas medidas en los sitios de muestreo de las regiones biogeográficas del Caribe y Orinoquía, Colombia, donde se registró Terpsinoë musica
| LPC* | Colector | Sistema | Coordenadas | Altitud (m s. n. m.) | Temperatura (°C) | pH (Unidades) | Conductividad (μS/cm) | Oxígeno disuelto (mg/l) | Caudal (m3.seg-1) | Sustrato |
|---|---|---|---|---|---|---|---|---|---|---|
| 5572 | Mario Medina | Río Lipa confluencia arroyo Primores | 6º 47’ 6,7’’ N 71º 1’ 34’’ W | 134 | 31,4 | 8,04 | 140 | 5,5 | 0,76 | Epifiton |
| 5571 | Río Lipa aguas arriba | 6º 49’ 33,2’’ N 71º 2’ 16,1’’ W | 134 | 29,5 | 7,4 | 100 | 5,5 | 0,46 | Epifiton | |
| 5568 | Arroyo Canta Rana | 11º 0’ 13,9’’ N 74º 3’ 46,8’’ W | 910 | 20,3 | 8,1 | 63 | 8 | 1,67 | Epiliton | |
| 5570 | Arroyo El Padre | 10º 58’ 36,1’’ N 74º 4’ 22,7’’ W | 928 | 18,3 | 7,9 | 92 | 7,5 | 0,15 | Epiliton |
* División Ficología del Museo de La Plata
La terminología utilizada para la descripción de los taxones es la sugerida en Barber y Harworth (1981). Para el ordenamiento sistemático se siguió a Cox (2015).
RESULTADOS
División Bacillariophyta
Clase Mediophyceae
Subclase Biddulphiophycidae
Orden Biddulphiales
Familia Biddulphiaceae
Género Terpsinoë
Especie Terpsinoë musicaEhrenberg 1843, p.425, Pl. 3, figs. IV-1 - VII- 30. Figuras 2 (A-I), 3 (A-E).
Valvas elípticas con márgenes triondulados; las ondulaciones o ensanchamientos tienen aproximadamente la misma proporción entre sí (Figuras 2A-E) a veces con una ligera reducción a la altura de las penúltimas barras transversales. Ápices capitados a rostrados con un pseudocelo formado por finos poroides organizados en hileras irregulares (Figura 2I). La superficie externa de la valva está cubierta por areolas fácilmente visibles al MO (Figuras 2F-H), dispuestas irregularmente en el centro y formando líneas irregulares más o menos longitudinales hacia los extremos (Figura 2G); al MEB se distinguen como rugosidades que se disponen de forma irregular por toda la valva (Figuras 2H-I). La valva tiene un rimoportula posicionado excéntricamente en el lóbulo central (Figuras 2F y G). Cada valva presenta seis barras transversales que se ordenan transapicalmente entre las porciones ensanchadas y los pseudocelos (Figuras 2A-D, 2G).
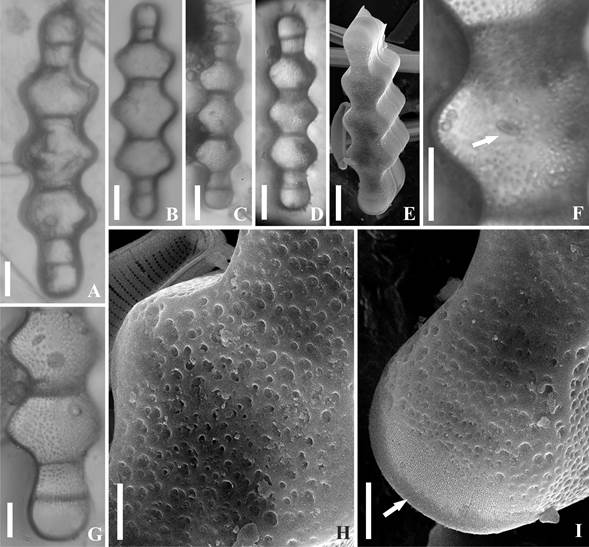
Figura 2 Terpsinoë musica. Especímenes en vista valvar. A-D, F-G. MO. E, H-I. MEB. A-E. Vistas generales. F. Detalle del centro mostrando el rimoportula (flecha). F-H. Detalle de las areolas en la superficie valvar. I. Detalle del pseudocelo (flecha). Escalas: A-E: 20 µm; F: 10 µm; G-I: 5 µm.
Frústulos rectangulares en vista conectival (Figuras 3A y B). Las barras transversales se observan en esta vista ocupando hasta 3/4 de la altura del manto, con los extremos engrosados de modo que en conjunto presentan aspecto de nota musical. Las barras transversales que se encuentran en las porciones ensanchadas son más largas que las que se encuentran cerca de los pseudocelos (Figuras 3A y B). Manto valvar alto, ornamentado con areolas irregularmente dispuestas (Figura 3C). Cingulum compuesto por pocas bandas cerradas (Figura 3C). Valvocópula con poroides pequeños dispuestos irregularmente; el borde de la pars interior presenta fimbrias (Figuras 3D y E).

Figura 3 Terpsinoë musica. Especímenes en vista conectival. A-B. MO. C-E. MEB. A-B. Note el ensanchamiento de las barras transversales. C. Detalle del manto valvar y del cingulum. D-E. Detalle de la valvocópula, las puntas de flecha indican las fimbrias. Escalas: A-C: 20 µm; D: 10 µm; E: 5 µm.
Datos morfométricos (n=30): Eje apical: 105-170 (m ( x = 120,2 (m; CV = 8,30); eje transapical (centro): 26,3-38,5 (m ( 𝑥 = 33,3 (m; CV = 15,26); L/A: 2,9-4,2; eje pervalvar: 43,5-106,4 (m ( 𝑥 = 62,9 (m; CV = 42,99); Altura del manto: 17,1-25,5 µm ( 𝑥 = 20,9 (m; CV = 12,36); areolas: 7-9 en 10 (m. En el ejemplar fotografiado al MEB, la densidad de los poroides del pseudocelo fue de 66-70 en 10 µm y la densidad de poroides en la valvocópula fue de 15-18 en 10 µm.
En este estudio esta especie se encontró asociada a comunidades propias del epifiton (sobre Eichhornia crassipes y Lenna minor) del río Lipa, perteneciente a la cuenca del río Orinoco, y del epiliton en los arroyos Canta Rana y El Padre, ubicados en la región Caribe. Como se detalla en la Tabla 1, en los sitios mencionados las aguas tienden a la alcalinidad con valores de pH entre 7,4 y 8,1, valores de conductividad eléctrica entre 60 y 140 μS/cm y bajos valores de caudal (entre 0,15-1,67 m3.seg-1) (Tabla 1).
Tabla 2 Comparación de los datos morfológicos y morfométricos de poblaciones de Terpsinoë musica registradas en Colombia y en otras regiones del mundo
| Referencia | Localidad | Eje apical (µm) | Eje trans apical (µm) | L/A | Profundidad del manto (µm) | Eje pervalvar (µm) | Núm. barras trans-versales | Densidad de areolas en 10 µm | Rimoportula | Pseudocelo | Espinas |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Carmona-Jiménez et al. (2017) | Arroyos tropicales de México | 80,2-135,6 | 33,6-50 | - | 16-24† | 64-122 | 2-6 | - | 1-(2-3), apertura interna en forma de S | poroides aleatoriamente distribuidas | pequeñas espinas marginales |
| El-Awamri et al. (2003) | El Cairo, Egipto | 111-140 | 35-49 | 3 | 12-17 | 64-105 | 6 | 12-14 | 1, apertura interna en forma de S | poroides irregularmente dispuestas | no |
| Wu (2013) | - | 56-133 | 32-44 | 1,6-3,7 | 21-27 | 52-99 | 2-6 | - | 1 | - | no |
| Baudrimont (1973) | Argelia | 40-200 | - | - | - | 35-40 | 6 | 10 | 1 | - | no |
| Luttenton et al. (1986) | Oklahoma, EE.UU | 49-116 | 37-43 | - | - | - | 2-6 | - | 1 | - | no |
| Navarro y Lobban (2009) | Guam | 100-110 | 85-95 | - | - | - | - | 8-10 | - | - | no |
| Este estudio | Regiones de Orinoquia y El Caribe | 105-170 | 25-30 | 4,2-5,6 | 14-26 | 68-88 | 6 | 8-10 | 1 | 66-70* poroides dispuestos en hileras irregulares | no |
† En la descripción de T. musica, Carmona-Jiménez et al. (2017), expresan que la profundidad del manto es de 64,4-125 µm, casi el mismo rango que para la medida del eje pervalvar; consideramos que se trata de un error y por tanto las medidas incluidas en la presenta tabla son las resultantes de medir los ejemplares de las fotos publicadas.
*Dato tomado a partir de un único ejemplar.
DISCUSIÓN
La comparación de los ejemplares de T. musica hallados en Colombia respecto a otras poblaciones de T. musica registradas en otros sitios, muestra que estos especímenes tienen una amplia variabilidad morfológica, particularmente en la longitud del eje apical y del eje pervalvar (Tabla 2). Coinciden en aquellos aspectos que definen la especie tales como contorno de la valva, presencia y posición del rimoportula, caracteres de las barras transapicales y pseudocelos. Una diferencia destacable es la disposición de las aréolas en la superficie valvar; aparentemente aleatoria o irregular en el centro (Boyer 1900, Carmona-Jiménez et al. 2017, El-Awamri et al. 2003, Frenguelli y Cordini 1937, Luttenton et al. 1986 y nuestro material) o radiales desde un punto central y luego paralelas/longitudinales hacia los extremos (Baudrimont 1973). Por otra parte, Carmona-Jiménez et al. (2017) documentaron algunos especímenes que presentaron pequeñas espinas en el margen valvar y más de un rimoportula.
Respecto a las características ecológicas, en general se la considera una especie de ambientes continentales salobres (aunque pueden ocurrir también en aguas dulces o marinas costeras) con una distribución tropical a subtropical (Baudrimont 1973, Carmona-Jiménez et al. 2017, El-Awamri et al. 2003, Luttenton et al. 1986, Round et al. 1990, Sterrenburg 1994). Según El-Awamri et al. (2003), la especie se desarrolla mejor en aguas duras, ligeramente alcalinas y salobres, con una conductividad eléctrica relativamente alta y temperaturas moderadamente altas. Por otra parte, Baudrimont (1973) la registró en aguas bicarbonatadas, sulfatadas y clorosulfatadas y la clasificó como una especie ( mésohalobe y alcalófila, que crece sobre filamentos de Cyanophyceae y Stigeoclonium. Carmona-Jiménez et al. (2017) la observaron en aguas neutras a ligeramente alcalinas (pH 7-8 unidades), con conductividades eléctricas entre medias a altas (268-2640 μS cm-1) y dominadas por sulfato/bicarbonato y calcio/magnesio. Asimismo, Luttenton et al. (1986) la registraron en aguas alcalinas, de conductividades moderadamente altas y altos valores de dureza. En Colombia se hallaron en el epifiton y el epiliton de sistemas lóticos en ambientes con aguas ligeramente alcalinas, baja conductividad y de bajo caudal.
Si bien la mayoría de las citas se refieren, en efecto, a localidades tropicales a subtropicales, la especie también ha sido citada en ambientes templados de agua dulce, como es el caso del río Delaware (Boyer 1900) y el lago Michigan en USA (Wujek y Welling 1981), en arroyos de la provincia de Buenos Aires, Argentina (Vouilloud 2003) y en sedimentos cuaternarios de la llanura pampeana, Argentina (Frenguelli y Cordini 1937, Frenguelli 1945).
En la descripción original de T. musica, Ehrenberg (1843) destacó la presencia de las estructuras con forma de “notas musicales” como característica de la especie y la ilustró con dibujos de dos ejemplares observados en vista conectival. No aportó ningún detalle de la vista valvar ni parámetros morfométricos, por lo que la descripción resulta insuficiente al momento de establecer comparaciones con poblaciones registradas en otros estudios. El material tipo de T. musica no ha sido estudiado posteriormente por ningún otro autor. Wu (2013), basado en la descripción original de T. musica y considerando la gran variabilidad morfológica entre diferentes poblaciones de la especie, planteó que es posible que estemos frente a un complejo de especies (flock), y en este sentido la localidad tipo es un dato importante al momento de analizar su taxonomía. Tuji (2018), empleando técnicas moleculares, comparó la población tipo de T. muninensis Tuji con una población de T. musica de Texas (sitio cercano a la localidad tipo de T. musica); el autor interpretó que las diferencias halladas en las secuencias apoyan la justificación de la descripción de su nuevo taxón, resaltando la importancia de considerar la distribución geográfica como factor relevante en el tratamiento taxonómico.
Para resolver esta cuestión es imprescindible, en primer lugar, analizar el material tipo de T. musica con técnicas modernas de análisis morfológico, a fin de establecer su identidad y así comparar la morfología fina de las distintas poblaciones y determinar si estamos frente a un taxón con una gran variabilidad morfológica y amplia distribución geográfica o, por el contrario, si se trata de un complejo de especies con requerimientos ecológicos y distribuciones más restringidas.

