











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
Los bosques tropicales son uno de los ecosistemas más diversos sobre el planeta, además de proporcionar diversos servicios ecosistémicos (de Beenhouwer, Aerts, y Honay, 2013). Sin embargo, acciones como la extracción de recursos, los asentamientos humanos, el exceso de residuos y la modificación de grandes áreas de producción (sistemas agroforestales, monocultivos, pastos para ganadería, etc.) degradan estos ecosistemas y ponen en peligro la biodiversidad que contienen (de Beenhouwer et al, 2013). Así, los ecosistemas pierden capacidad de recuperación y los servicios que prestan se ven amenazados (Andrade, 2011).
La mayoría de esfuerzos por conservar esta biodiversidad, se han enfocado en aislar áreas relativamente bien conservadas de ecosistemas naturales bajo figuras de áreas protegidas (Sánchez-Clavijo, Durán, Vélez, García, y Botero, 2010). Sin embargo, estas áreas naturales protegidas no son suficientes, ya que cubren menos del 15% de la superficie terrestre mundial (UNEP-WCMC y IUCN, 2016). Por consiguiente, es necesario complementar las áreas naturales protegidas con otras manejadas por principios ecológicos, que garanticen tanto la autosuficiencia como la producción de recursos, así como la conservación de la biodiversidad (Moguel y Toledo, 1999; Zermeño-Hernández, Pingarroni, y Martínez-Ramos, 2016).
Los sistemas agroforestales se consideran una alternativa o complemento para amortiguar la pérdida de biodiversidad, ya que son cultivos agrícolas con un manejo intencional de árboles de sombra, siendo importantes puesto que no remueven la totalidad de árboles del bosque y conservan parches de bosque nativo. Estos sistemas pueden aliviar la presión sobre el uso de recursos en las áreas protegidas, aumentar la conectividad de los componentes del paisaje (Bhagwat, Willis, Birks, y Whittaker, 2008; Zermeño-Hernández et al, 2016) y además, por su estructura multiestratificada, pueden proporcionar hábitat (Hall, Krausman, y Morrison, 1997) y diversos recursos a una significativa diversidad de animales y plantas, dependiendo del mantenimiento de la cubierta forestal y la intensificación del uso de la tierra (Cassano, Barlow, y Pardini, 2012; Guiracocha, Harvey, Somarriba, Krauss, y Carrillo, 2001).
Uno de los cultivos desarrollados bajo esta modalidad es el de cacao, Theobroma cacao L.; en Colombia, el cacao tiene un área total sembrada de 176.050 ha, cultivándose en suelos con características de piso térmico cálido, que comprende la franja de tierras ubicadas desde el nivel del mar hasta los 1300 m s. n. m. El departamento que concentra la mayor producción es Santander con el 41,45% de participación, lo que equivale a 23.574 toneladas. Le siguen en importancia, con menor participación: Antioquia, Arauca, Huila, Tolima, Nariño y Cesar, los cuales, en conjunto representan el 40,85% del total nacional (Federación Nacional de Cacaoteros, 2018).
Debido a la gran área que abarcan estos cultivos, su proximidad con la región Andina y su alta biodiversidad, son inevitables los encuentros con la fauna silvestre (ungulados, primates, roedores, aves granívoras, entre otros) que persiste en estas zonas. Eventualmente, algunas de estas especies pueden ocasionar daños a los cultivos o entrar en conflicto con las personas y poblaciones que dependen de dichos cultivos como sus ingresos económicos primarios (Conover, 2001; Marchini, 2014; Sillero-Zubiri, Sukumar, y Treves, 2007).
La fauna silvestre, que cohabita con los agroecosistemas, puede generar problemas con los productores por el eventual consumo y destrucción de cultivos y alimentos almacenados o la transmisión de enfermedades a humanos u otros animales, entre otras interacciones (Sillero-Zubiri et al, 2007; Treves, Wallace, y White, 2009). Estudios previos han registrado casos de conflictos con fauna y evaluado el daño hecho por mamíferos en plantaciones. Por mencionar algunos, Hafidzi (1998) reportó ratones y ardillas como las mayores plagas para los cultivos de cacao en Malasia, realizando un estudio sobre patrones de actividad de ardillas en estos cultivos, para proponer estrategias de manejo. Mollineau, Bekele, y García (2008) reportan que, en plantaciones de cacao a nivel industrial, en Trinidad, la ardilla roja (Notosciurus granatensis) tiene una marcada preferencia por los frutos maduros, dejando cuantiosas pérdidas, ya que hasta los frutos ligeramente afectados se pierden debido a la posterior acción de hongos. También indicaron que las zonas más afectadas son los bordes de los cultivos, puesto que están próximos a áreas boscosas, lo que permite su movimiento entre las zonas de alimentación (los cultivos) y sus refugios (los bosques). Por lo tanto, el manejo de estos encuentros (conflictos), con la fauna silvestre se aborda desde diferentes perspectivas, resultando en diversas respuestas por parte de las especies nativas (Muñoz y Muñoz-Santibáñez 2016).
En Colombia, los estudios sobre fauna en sistemas agroforestales, específicamente sobre mamíferos no voladores asociados a estos sistemas son limitados y tampoco son muchos los que registran las afectaciones hechas por estas especies a los cultivos. Por ejemplo, una evaluación de la riqueza y abundancia relativa de mamíferos no voladores habitando un cultivo de palma africana, en el departamento de Casanare, encontró un total de 16 especies, donde la mayoría correspondían a mesodepredadores y generalistas (Pardo-Vargas y Payán, 2015). Sin embargo, estudios enfocados en ensamblajes de murciélagos asociados a cultivos de café (Castaño, Botero, Velásquez, y Corrales, 2004; Numa, Verdu, y Sánchez-Palomino, 2005; Ortegón-Martínez y Pérez-Torres, 2007), registran la abundancia de especies como Glossophaga soricina, Sturnira lilium, Artibeus lituratus y Carollia brevicauda; concluyendo que los cafetales con sombrío y las coberturas de bosque nativo (parches) constituyen una fuente importante de alimento y refugio para algunos murciélagos frugívoros y nectarívoros. Otros estudios en sistemas agro-productivos (Botero Echeverri, Lentijo Jiménez, y Sánchez-Clavijo, 2014; Sánchez-Clavijo et al, 2008, 2010), aunque no se enfocan en mamíferos, cobran importancia pues incluyen estudios donde se evalúa la biodiversidad asociada a cultivos de café en el país, enfocándose en aves, plantas y hormigas. Observaciones ocasionales también han mostrado algunos mamíferos involucrados en el daño a plantaciones de pino (Pinus patula) en el oriente de Antioquia (S. Solari, obs. pers.). Sin embargo, no se encontraron reportes publicados que abarquen la fauna asociada a cultivos de cacao realizados en Colombia.
Históricamente, las soluciones al conflicto con la fauna han sido letales, mediante cacería, envenenamiento, trampas o exclusión completa (Treves y Naughton-Treves, 2005). Sin embargo, los mamíferos considerados como potenciales plagas deben ser vistos, además, en términos de sus interrelaciones con los componentes del ecosistema y los potenciales roles que desempeñan (Bhagwat et al, 2008; Jose, 2009; Lacher et al, 2019; Rumiz, 2010). Algunos de estos roles ecológicos incluyen: influir en la composición vegetal del bosque (al depredar semillas y plántulas), en las propiedades químicas y físicas del suelo (aireación al excavar madrigueras), en la productividad primaria (consumo de hojas y frutos); en el reciclaje de nutrientes (carroñeros); al proporcionar fuente de alimento base para carnívoros o el control de insectos y fauna menor, entre otras (Aguilera, 1985; Rumiz, 2010; Sieg, 1987; Solari, Rodríguez, Vivar, y Velazco, 2002; Voss, 1991). Todo lo anterior contribuye a la integridad de los ecosistemas y al flujo de materia y energía entre los diferentes ambientes (Feldhamer, Drickamer, Vessey, Merritt, y Krajewski, 2007; Nagy‑Reis, Ribeiro, Setz, y Chiarello, 2019; Norris, 2012; Rojas y Moreno, 2014). Por todo lo anterior, la alteración de las comunidades de mamíferos, a través de programas de control (exclusión y cacería), influye en otros componentes y, finalmente, sobre todo el sistema (Sieg, 1987). Para contrarrestarlo, se ha generado un interés por alternativas a las represalias, como el manejo no letal de fauna, elevando la tolerancia para la vida silvestre (Treves et al, 2009), que permita el funcionamiento de estas redes de interacción y el desempeño de sus roles ecológicos. Una visión integral al tema de los conflictos con fauna silvestre pasa necesariamente por entender estos roles a escala local y regional (Nagy-Reis et al, 2019), lo que puede generar mayor tolerancia a la presencia de estas especies y un manejo racional ambientalmente.
La Compañía Nacional de Chocolates ha mostrado interés en saber cuáles mamíferos no voladores persisten en la Granja Yariguíes, ubicada en el departamento de Santander, ya que al incluir parches de bosque natural como zonas de reserva alrededor de los cultivos experimentales de cacao, se estaría favoreciendo la permanencia de estos animales. Por otro lado, también existe el interés en cuantificar los daños causados a las mazorcas de cacao, principalmente por la ardilla roja (N. granatensis) e identificar qué otras especies pueden estar causando afectaciones negativas. En este sentido, el objetivo de este estudio fue identificar los roles ecológicos suministrados por mamíferos no voladores en un sistema agro-productivo de cacao, incluyendo los posibles daños que algunas especies ejercen sobre el cultivo.
MATERIALES Y MÉTODOS
Lugar de estudio
El lugar de muestreo fue la Granja Yariguíes, propiedad de la Compañía Nacional de Chocolates, ubicada en la vereda La Lejía, entre los municipios de San Vicente del Chucurí y Barrancabermeja, Santander. Con una extensión de 263 ha, donde 210 ha son destinadas al cultivo de cacao, mientras que 53 ha están destinadas a áreas de reserva y protección. La formación predominante (zona de vida) es bosque húmedo tropical (Hernández-Camacho, Hurtado, Ortiz, y Walschburger, 1992). La granja es atravesada por el río Oponcito y está sobre la vía Ruta del Sol (Figura 1). La zona de cultivo de cacao está distribuida en 39 lotes, de los cuales se escogieron dos áreas que cumplieran las siguientes condiciones: (1) que tuvieran borde colindante con el bosque de reserva; lote 10, con un área de 7,3 ha, y (2) que fueran netamente zona de cultivo; lotes 5 y 6, con un área de 3,5 ha y 3 ha, respectivamente, sumando un total de 6,5 ha. Se escogieron estos dos lotes juntos con el fin de tener áreas comparables, en cuanto al tamaño, con la parcela 10 (Figura 1).
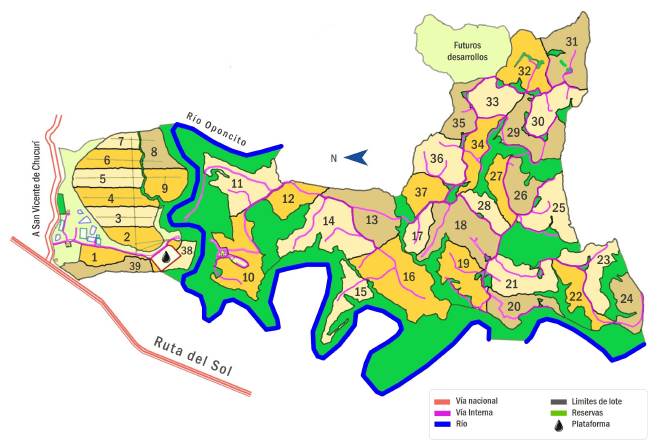
Diseño de muestreo
Los muestreos comprendieron cinco salidas de campo (una por mes) con una duración aproximada de cinco días cada una, desde septiembre de 2017 a enero de 2018. Los muestreos se dividieron en identificación y cuantificación de daños a las mazorcas (septiembre - diciembre 2017) y registro de mamíferos (septiembre 2017 - enero 2018). En cada uno de los lotes se hicieron cinco recorridos lineales de 200 m de longitud (2 m a cada lado), ubicando los árboles con frutos afectados, para luego identificar y cuantificar los daños causados por roedores silvestres (ardillas, ñeques, puercoespines). Adicionalmente, estos recorridos permitieron identificar los lugares más afectados y evaluar si había alguna preferencia por el estado de maduración (inmaduro, medio o maduro) de los frutos. Los daños a las mazorcas se determinaron por observación directa, mientras que su cuantificación se calculó a partir del número de frutos afectados (mordidos o consumidos) por cada lote, así como mediante el porcentaje de estos respecto al total disponible (por lote). Finalmente, la identificación de la especie causante se hizo mediante comparación directa de las marcas de los incisivos con la dentición de especímenes depositados en la Colección Teriológica de la Universidad de Antioquia (CTUA).
Para determinar la presencia de los mamíferos que habitan la zona, se hicieron recorridos que permitieron encontrar rastros como huellas, madrigueras, excretas, entre otros (Aranda, 2012; Navarro y Muñoz, 2000), así como áreas de presencia potencial de especies menores, usualmente con buena cobertura original (bosque húmedo). Para obtener registros directos, se instalaron cuatro cámaras de detección automática (cámaras trampa), en lugares donde se consideró la potencial presencia de mamíferos (sin usar atrayentes), las cuales se cambiaron de lugar cada mes para abarcar nuevos lugares que permitieran obtener más registros. También se dispusieron 40 trampas de captura viva, Sherman ® (23 cm), con el fin de obtener registros de pequeños mamíferos, que se ubicaron en líneas a través del borde del bosque o cerca a quebradas, donde se esperaba la presencia de estas especies, separadas entre sí por aproximadamente 10 m. Se usó una mezcla de hojuelas de avena y mantequilla de maní como cebo.
Por otro lado, se encuestaron algunos colaboradores de la granja, incluyendo técnicos (4), personal de vigilancia (2) y supervisores (1), acerca de la presencia de los mamíferos que habitan la zona, ya que ellos tienen un conocimiento más amplio del lugar y podrían dar información sobre las especies que ven frecuentemente y los lugares que transitan. Para una mayor precisión en las identificaciones, se empleó el apoyo visual con dibujos en guías de mamíferos neotropicales (Eisenberg, 1989; Emmons y Feer, 1990).
El esfuerzo de muestreo se definió de diferentes formas según el método empleado; para las cámaras trampa se definió como la suma del número de horas que las cámaras estuvieron activas multiplicado por el número de cámaras utilizadas. Para las trampas Sherman ®, se definió como el número de noches que las trampas estuvieron activas multiplicado por el número de trampas (Voss y Emmons, 1996), mientras que para los recorridos libres como la suma del tiempo invertido por día muestreado (Pardo-Vargas y Payán-Garrido, 2015).
Roles ecológicos
Estos mamíferos, además de la posible afectación a los cultivos, también juegan un rol particular dentro del ecosistema estudiado, siendo necesario establecer cuáles son estas especies y con cuáles otras coexisten en el lugar de estudio. Una vez identificados taxonómicamente se estableció, mediante la revisión de literatura pertinente, sus roles ecológicos en este ambiente. Para determinar estos roles, las especies se dispusieron por gremios, término definido por Root (1967), el cual agrupa las especies que explotan la misma clase de recursos ambientales de una manera similar sin tener en cuenta la posición taxonómica. Para esto se tomó en cuenta la dieta de cada especie registrada durante este estudio, así como el sustrato utilizado por cada una y el horario predominante de su actividad (González-Salazar, Martínez-Meyer, y López Santiago, 2014):
Dieta: para clasificar las especies por su dieta se consideró el principal recurso alimenticio utilizado por los mamíferos encontrados como: vertebrados e invertebrados no-artrópodos (carnívoros), insectos (insectívoros), carroña (carroñeros), frutos (frugívoros), hojas, tallos o semillas (herbívoros).
Sustrato de forrajeo: se refiere al lugar donde los organismos obtienen el alimento. Se consideraron tres sustratos principales: el suelo, los árboles y los cuerpos de agua, donde se desempeñan como cazadores, recolectores y excavadores, entre otros.
Periodo de actividad: se dividió en dos clases: 1) Diurna: si el periodo de actividad se inició en la mañana y continuó durante el día (06:00 a 18:00 h). 2) Nocturna: si la actividad comenzó a altas horas de la tarde y continuó durante toda la noche (18:00 a 06:00 h).
Para asignar las especies a un gremio se consideraron todos los datos obtenidos, ya sea por observación durante el muestreo o por información obtenida de la literatura. Por otro lado, en los casos que no se pudo determinar el tipo de dieta para la especie, se consideró la información reportada para el género (González-Salazar et al, 2014).
Para evaluar las diferencias entre las especies en relación con los daños causados al cacao, así como entre la actividad de la ardilla entre parcelas y entre meses, utilizamos una prueba de Kruskal-Wallis (p=0,05) dado que los valores obtenidos no cumplieron los supuestos requeridos para una prueba de ANOVA (McDonald, 2014).
RESULTADOS
Daños en los cultivos
Se registró a la ardilla roja (N. granatensis) como la especie que más atacó las mazorcas de cacao (Kruskal-Wallis H=8,3434; gl=2; p=0,015), seguida del ñeque (Dasyprocta punctata) y del puercoespín (Coendou sp.), aunque estos dos últimos en una muy baja proporción (Tabla 1). La incidencia de daño en los lotes 5-6 fue muy baja y el porcentaje de daño no varió mucho entre muestreos (0 a 0,34%), excepto por septiembre, que mostró un porcentaje más alto (3,2%); un patrón similar se observó en el lote 10 (Figura 3). Sin embargo, cuando se combinaron ambos lotes, el número de frutos afectados en septiembre (86) no fue muy distinto al registrado en el mes de octubre (84), lo que evidencia que, en general, hay un comportamiento similar en estos dos meses. En general, se observó un decrecimiento en el número de frutos y el porcentaje de daño a medida que pasa el tiempo (Tabla 1), pero que no llega a ser significativo (H=3; gl=3; p=0,3916). También se pudo identificar que la zona más afectada fue el lote 10, (Figura 2), aunque las diferencias no resultaron significativas (H=2.1084; gl=1; p=0,1465).
Tabla 1 Número de frutos afectados por ardilla roja (Notosciurus granatensis), ñeque (Dasyprocta punctata) y puercoespín (Coendou sp.) en cada lote por mes de muestreo. Se observa la mayor incidencia de las ardillas en el lote 10, mientras que los ñeques y puercoespines no representan mayor daño. Para los lotes 5-6, los daños fueron comparativamente más bajos que los del lote 10.
| Especie causante de daños | ||||||
| Mes | Parcela 5 -6 | Parcela 5 -6 | Parcela 5 -6 | Parcela 10 | Parcela 10 | Parcela 10 |
| Ardilla | Ñeque | Puercoespín | Ardilla | Ñeque | Puercoespín | |
| Septiembre | 3 | 0 | 0 | 81 | 2 | 0 |
| Octubre | 5 | 0 | 0 | 79 | 0 | 0 |
| Noviembre | 0 | 0 | 0 | 43 | 0 | 1 |
| Diciembre | 3 | 0 | 0 | 12 | 2 | 0 |
| Total | 11 | 0 | 0 | 215 | 4 | 1 |
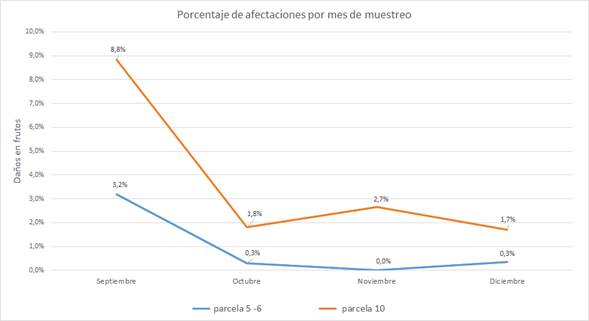
Figura 2 Porcentajes de afectaciones en frutos por mes de muestreo. La línea azul representa los daños para los lotes 5-6; la línea roja representa los daños para el lote 10.
En cuanto a la preferencia por algún estado de madurez del fruto, se evidenció una fuerte predilección por los frutos maduros, incluyendo en promedio más del 50% de los frutos afectados (Tabla 2). Sin embargo, no se encontraron diferencias significativas entre las tres categorías definidas (H=5,258; gl=2; p=0,0715).
Tabla 2 Frutos afectados por grado de madurez en cada lote durante los meses de muestreo, donde se observa una clara preferencia por los frutos maduros.
| Afectación según maduración del fruto | ||||||
| Mes | Parcela 5-6 | Parcela 5-6 | Parcela 5-6 | Parcela 10 | Parcela 10 | Parcela 10 |
| Maduro | Medio | Inmaduro | Maduro | Medio | Inmaduro | |
| Septiembre | 2 | 1 | 0 | 51 | 10 | 22 |
| Octubre | 3 | 2 | 0 | 54 | 14 | 11 |
| Noviembre | 0 | 0 | 0 | 29 | 15 | 0 |
| Diciembre | 3 | 0 | 0 | 10 | 4 | 0 |
Registros de mamíferos
Se registró un total de 17 especies en cinco órdenes empleando los diferentes métodos antes mencionados (cámara-trampa, rastreo de huellas, rastreo de madrigueras, trampas Sherman ®, entre otros). Como se esperaba, la mayoría de los registros se dieron en el bosque de protección (Tabla 3). Mediante conversaciones con el personal de la granja se confirmaron algunos registros que inicialmente se tomaron como potenciales (v.g., nutria, chigüiro) ya que la evidencia no era concluyente. En la mayoría de casos fue posible determinar con precisión la especie a partir de las fotografías o videos, o cuando se tuvo acceso a algún material (ejemplares en trampas, restos); solo en el caso del puercoespín (Coendou sp.), identificado a partir del tamaño de la huella de los incisivos en las mazorcas, fue imposible establecer la identidad de la especie.
Tabla 3 Listado de mamíferos registrados en la Granja Yariguíes, donde se observa los meses en los cuales fueron registrados cada uno y el lugar donde se obtuvo el registro. Las letras B, C, R, corresponden a los lugares de muestreo Bosque, Cultivo, y Río respectivamente. * Indica registros hechos por fuera de estas fechas, por personal de la granja o por subsecuentes visitas de los investigadores (en Bosque).
| Mes de muestreo | ||||||
|---|---|---|---|---|---|---|
| Especie | Nombre común | Septiembre | Octubre | Noviembre | Diciembre | Enero |
| Dasypus cf. novemcinctus | Armadillo | B, C | B, C | B, C | B | B |
| Didelphis marsupialis* | Zarigüeya | B | ||||
| Metachirus nudicaudatus* | Chucha mantequera | |||||
| Marmosa robinsoni | Marmosa | |||||
| Tamandua tetradactyla* | Oso mielero | |||||
| Bradypus variegatus* | Oso perezoso | |||||
| Cuniculus paca | Guagua | B, C | B | B, R | B, R | B, R |
| Hydrochoerus hydrochaeris | Chigüiro | R | R | R | R | R |
| Notosciurus granatensis | Ardilla roja | B, C | B, C | B, C | B, C | B, C |
| Coendou sp. | Puercoespín | C | ||||
| Dasyprocta punctata | Ñeque | C | C | |||
| Proechimys cf. chrysaeolus | Rata espinosa | C | B | B | ||
| Leopardus pardalis | Tigrillo | B | C | C | ||
| Lontra longicaudis | Nutria | R | ||||
| Procyon cancrivorus | Mapache | B | B, R | B, R | B, C, R | B, C, R |
| Cerdocyon thous | Zorro perro | B, C, R | B, C, R | B, R | B, R | B, R |
| Eira barbara | Tayra | B | B |
Representatividad de Gremios
Dado que la definición de gremio usada por nosotros incluye grupo alimenticio, sustrato y horario de actividad, las 17 especies registradas se incluyeron en 13 gremios que representan cinco grupos alimenticios (insectívoros, carnívoros, herbívoros, frugívoros y omnívoros). En los casos en que no fue posible definir el sustrato, dado que algunas especies son capaces de utilizar distintos estratos (suelo y vegetación arbórea), solo se indicó el grupo alimenticio y el patrón de actividad (Tabla 4).
Tabla 4 Especies registradas en la Granja Yariguíes, agrupadas por grupos tróficos y hábitos (tipo de sustrato y patrón de actividad).
| Grupo trófico | Hábito | Orden | Familia | Especie |
|---|---|---|---|---|
| Insectívoro | Terrestre nocturno | Cingulata | Dasypodidae | Dasypus cf. novemcinctus |
| Insectívoro | Arbóreo norcurno | Pilosa | Myrmecophagidae | Tamandua tetradactila |
| Insectívoro | Arbóreo norcurno | Didelphimophia | Didelphidae | Marmosa robinsoni |
| Herbívoro | Arbóreo norcurno | Pilosa | Bradypodidae | Bradypus variegatus |
| Herbívoro | Acuático nocturno | Rodentia | Caviidae | Hydrochuerus hydrochaeris |
| Fruguívoros | Terrestre diurno | Rodentia | Dasyproctidae | Dasyprocta punctata |
| Fruguívoros | Terrestre nocturno | Rodentia | Cuniculidae | Cuniculus paca |
| Fruguívoros | Terrestre nocturno | Rodentia | Echimyidae | Proechimys cf. chrysaeolus |
| Fruguívoros | Arbóreo diurno | Rodentia | Sciuridae | Notosciurus granatensis |
| Fruguívoros | Arbóreo nocturno | Rodentia | Erethizontidae | Coendou sp. |
| Carníviro | Terrestre nocturno | Carnivora | Felidae | Leopardus pardalis |
| Carnívoro | Acuático diurno | Carnivora | Mustelidae | Lontra longicaudis |
| Omnívoro | Diurno | Carnivora | Mustelidae | Eira barbara |
| Omnívoro | Terrestre nocturno | Carnivora | Procyonidae | Procyon cancrivorus |
| Omnívoro | Terrestre nocturno | Carnivora | Canidae | Cerdocyon thous |
| Omnívoro | Nocturno | Didelphimorphia | Didelphidae | Didelphis marsupialis |
| Omnívoro | Nocturno | Didelphimorphia | Didelphidae | Metachirus nudicaudatus |
DISCUSIÓN
Dado que en la granja Yariguíes la cobertura original de las zonas de bosque, aunque fragmentadas, se ha mantenido sin mayores cambios durante los últimos 20 años, consideramos que éstas siguen proporcionando hábitats favorables y recursos para las especies nativas que persisten en la zona. Las zonas de cultivo, por otro lado, también son aprovechadas directamente para alimentación por animales como ardillas y ñeques, así como indirectamente (como corredores para tránsito) por armadillos, mapaches y zorros, entre otros. Sin embargo, no se evidencia una oferta de refugios o hábitats favorables para estas especies en la zona de cultivo, a causa de podas continuas, poca diversidad de plantas y el paso constante de trabajadores, entre otros factores. El avistamiento de diferentes evidencias en zonas de cultivo (como huellas de zorro, mapache, tigrillo y guagua, entre otros) sugiere que estas zonas proporcionan, principalmente, conectividad entre los parches de bosque (Beer et al, 2003). De esta manera, el mantenimiento de los parches de bosque con características similares a ambientes poco intervenidos estaría ayudando a atenuar los efectos de la intensificación agrícola al proporcionar hábitats para poblaciones de mamíferos, contribuyendo al mantenimiento de la biodiversidad y las redes tróficas en estos ecosistemas modificados (Gómez, Coda, Serafini, Steinmann, y Priotto, 2017).
La presencia de especies generalistas de hábitat (sensu Rozenweig, 1981), tales como zorros, mapaches, zarigüeyas y armadillos eran esperadas puesto que estas poblaciones toleran mejor los cambios ambientales (Abba, Zufiaurre, Codesido, y Bilenca, 2015; Bhagwat et al, 2008; Gómez et al, 2017). Pero, la presencia de especies de hábitos carnívoros, como tigrillos y nutrias, que requieren condiciones específicas de hábitat debido a sus dietas y comportamiento, sugiere que al menos durante el estudio se dieron condiciones aceptables en los parches de bosque que permitieron la presencia de estas especies (Figura 3). Igualmente, esto podría tomarse como un posible indicador de las buenas prácticas de manejo que se llevan a cabo en la granja. Aunque estos gremios tróficos no son equivalentes a grupos funcionales, posibles de asociar a ciertos servicios ecosistémicos (Blondel, 2003), esta caracterización permite un primer acercamiento a la estructura del ensamblaje de mamíferos (Rumiz, 2010) en una zona productiva y transformada, incluyendo ciertos procesos a escala del ecosistema y cómo ciertas especies podrían estar cumpliendo roles similares y complementarios (Lacher et al, 2019).

Figura 3 Representación del entorno estudiado en la Granja Yariguíes, mostrando algunas de las especies registradas. De izquierda a derecha: Ocelote (Leopardus pardalis; carnívoro terrestre nocturno), Tamandúa (Tamandua tetradactyla; insectívoro arbóreo nocturno), Marmosa (Marmosa robinsoni; omnívoro arbóreo nocturno), Chigüiro (Hydrochoerus hydrochaeris; herbívoro acuático nocturno), Zorro perro (Cerdocyon thous; omnívoro terrestre nocturno), Armadillo (Dasypus novemcinctus; insectívoro terrestre nocturno), Guagua (Cuniculus paca; frugívoro terrestre nocturno), Ardilla de cola roja (Notosciurus granatensis; frugívoro arbóreo diurno).
En cuanto a los daños en los cultivos, en los lotes 5-6 se registraron pocos daños (máximo cinco frutos por mes muestreado), mientras que el lote 10 tuvo los mayores registros de daño (con un máximo de 83 frutos afectados y un mínimo de 14), sobre todo en las mazorcas ubicadas en las primeras líneas de cultivo y que limitan con el bosque. Es posible que esto se deba principalmente a esa cercanía, ya que, como se ha mostrado previamente, el bosque ofrece un espacio seguro para los mamíferos, de manera que éstos solo utilizan los cultivos para alimentación o conexión con otras zonas por esa conveniencia (de Beenhouwer et al, 2013; Mollineau et al, 2008). La progresiva disminución de las afectaciones podría deberse a las dinámicas de la finca, donde los meses de noviembre y diciembre son las de mayor cosecha, por lo que la disponibilidad de mazorcas se ve reducida y al mismo tiempo aumenta la presencia de personal a través de los sembríos.
En general, estos valores sugieren que los mamíferos no están representando grandes problemas ni pérdidas económicas importantes en estos cultivos. La baja incidencia puede darse por una moderada población de las especies que afectan los cultivos (ardillas, ñeques, puercoespines), que tal vez está siendo controlada por depredadores que persisten en la zona (como zorros y tigrillos), lo que es importante pues dicho equilibrio ayuda a mantener reguladas las poblaciones de potenciales plagas como los roedores (Nagy-Reis et al, 2019; Rumiz, 2010), evitando así algunos conflictos mayores (daños severos) que se pudieran tener con la fauna local. Sin embargo, la diversidad de pequeños y medianos mamíferos también ayuda a sostener otras funciones básicas del ecosistema, ya sea conectando parches de hábitats diferentes o similares mediante el transporte de materia orgánica, nutrientes y minerales a lo largo del paisaje (Lacher et al, 2019; Priotto, 2017; Rumiz, 2010).
En cuanto a los gremios identificados, estos representan roles ecológicos que, dependiendo del recurso que explotan y la forma en que lo hacen, pueden traer varios beneficios tanto para los bosques como para los cultivos (Lacher et al, 2019). Por ejemplo, las especies de hábitos insectívoros pueden ejercer control poblacional sobre insectos que afectan las plantas; mientras que, los armadillos al ser semifosoriales airean y remueven el suelo al hacer excavaciones continuas en busca de alimento o elaboración de madrigueras, lo que permite la aireación y remoción del suelo evitando la compactación (Abba et al, 2015; Bilenca et al, 2017; Rumiz, 2010). De otro lado, animales como tigrillos (carnívoros), zorros, mapaches y tayras (omnívoros), pueden estar influenciando las poblaciones de pequeños mamíferos que pueden ejercer daños en los cultivos (Nagy-Reis et al, 2019) o que pueden afectar el ecosistema si sus poblaciones aumentan considerablemente, por ejemplo, consumiendo semillas y plántulas que podrían perjudicar la dispersión y por ende, la composición del bosque (Rojas-Robles, Stiles, y Muñoz-Saba, 2012). Igualmente, herbívoros como el chigüiro y la guagua se alimentan sobre la tierra y defecan en el agua, favoreciendo el enriquecimiento de los ambientes acuáticos con materia orgánica y semillas, de lo cual se beneficiarían los peces y otros organismos (Beck, von Helversen, y Beck-King, 1999). Adicionalmente, para las especies que explotan los mismos recursos, es posible reducir la competencia directa (p. ej., por alimentos, en caso de su escasez) ya sea utilizando un estrato o un horario de actividad diferente (González-Salazar et al, 2014).
Debe recordarse también que, roedores como las ardillas, ñeques y guaguas (gremio de frugívoros), también influyen en la composición de los bosques, ya que la depredación de los frutos permite el transporte de semillas (Lacher et al, 2019; Rumiz, 2010). Los frugívoros consumen los frutos cerca de la planta parental y en algunos casos alejan y entierran semillas para consumo posterior, las cuales en gran parte son olvidadas, lo que permite la germinación de las plantulas, aumentando las áreas ocupadas por estas y sus rangos de distribución (Rojas-Robles et al, 2012). Esto cobra importancia teniendo en cuenta que en los bosques tropicales, más del 75% de las especies leñosas dependen de animales para dispersar sus semillas (Janzen y Vázquez-Yanes, 1991). Estas especies, junto con otras como el puercoespín (Coendou sp.), pueden estar actuando directamente sobre la regeneración natural del bosque ya que, al ramonear algunas especies de plantas de rápido crecimiento, ayudan a reducir la capacidad competitiva de estas, favoreciendo el crecimiento de plantas con más lento crecimiento (Lacher et al, 2019; Rumiz, 2010).
Por último, cabe resaltar que la Compañía Nacional de Chocolates no hace control o manejo de animales considerados “plagas”, y por el contrario, al mantener una zona de reserva en esta granja ayuda con la preservación de estas poblaciones. El rol ecológico de las especies presentes en este ecosistema productivo, complementado con medidas relativas de su valor funcional como reguladores de otras poblaciones, y sobre la dinámica general del ecosistema, resulta altamente provechoso para clasificar y categorizar especies o grupos de especies, tanto con fines de investigación como de conservación (Lacher et al, 2019; Nagy-Reis et al, 2019). Por ello, este proceso resulta crítico para el mantenimiento de esta biodiversidad y su dinámica, y favorece un uso racional y adecuado de los servicios ecosistémicos asociados.

