










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.27 no.1 Bogotá June 2005
LAS ESPECIES COLOMBIANAS DEL GÉNERO DIGITARIA (POACEAE: PANICOIDEAE: PANICEAE)
The Colombian species of the genus Digitaria (Poaceae: Panicoideae: Paniceae)
DIEGO GIRALDO CAÑAS
Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Apartado 7495, Bogotá D. C., Colombia. dagiraldoc@unal.edu.co
RESUMEN
Se presenta un estudio sistemático, macro y micromorfológico de las especies colombianas del género Digitaria. Se analizan diversos aspectos relativos a la clasificación, la nomenclatura, las afinidades genéricas y la variación macro y micromorfológica de los caracteres, así como algunos aspectos anatómicos de la espiguilla y el antecio superior. El género Digitaria está representado en Colombia por 18 especies distribuidas en cinco secciones: Digitaria sect. Calvulae (una especie), Digitaria sect. Digitaria (siete especies), Digitaria sect. Ischaemum (tres especies), Digitaria sect. Ternatae (cuatro especies) y Digitaria sect. Trichachne (tres especies). Dos especies son endémicas de Colombia (Digitaria cardenasiana y Digitaria andicola). Las especies Digitaria argillacea, Digitaria neesiana y Digitaria fragilis se excluyen de la flora de Colombia. Se presentan las claves para reconocer las secciones y las especies presentes en Colombia, así como también las descripciones de éstas, la iconografía, los usos, los nombres vulgares, la distribución geográfica y se comentan algunas observaciones morfológicas y ecológicas. Se documenta la presencia de espiguillas trifloras en el género Digitaria, situación observada en las especies Digitaria dioica y Digitaria nuda.
Palabras clave. Digitaria, Poaceae, Paniceae, Gramíneas neotropicales, Flora de Colombia.
ABSTRACT
A systematic study of the Colombian species of the genus Digitaria is presented. The aspects related to the classification, nomenclature, generic affinities, and macro and micromorphological variation are discussed. Some anatomical features of the spikelet and upper anthecium are analyzed. The genus Digitaria is represented in Colombia by 18 species and five sections: Digitaria sect. Calvulae (one species), Digitaria sect. Digitaria (seven species), Digitaria sect. Ischaemum (three species), Digitaria sect. Ternatae (four species), and Digitaria sect. Trichachne (three species). Two species are endemic to Colombia (Digitaria cardenasiana and Digitaria andicola). Digitaria argillacea, Digitaria neesiana, and Digitaria fragilis are excluded from Colombian Flora. Keys to recognize all sections and species that inhabit Colombia are given, as well as description of the species, iconography, uses, vernacular names, specimens examined, geographical distribution, and morphological and ecological observations. The phenomenon of 3-flowered spikelets is recorded for the genus Digitaria and it was verified in two species: Digitaria dioica and Digitaria nuda.
Key Words.Digitaria, Poaceae, Paniceae, Neotropical grasses, Flora of Colombia.
INTRODUCCIÓN
El género Digitaria Haller pertenece a la tribu Paniceae de la subfamilia Panicoideae y éste se caracteriza por presentar la lema superior cartilagínea, glabra y con los bordes membranáceos, generalmente hialinos y plegados sobre la pálea superior. Digitaria es reconocido como uno de los géneros más difíciles de la tribu Paniceae y la dificultad taxonómica radica en su gran número de especies, su amplia distribución geográfica, su complejidad de caracteres y su pobre conocimiento de las relaciones entre las especies (Webster 1983, Webster & Hatch 1990). Recientemente se confirmó la monofilia de Digitaria con base en análisis morfológico-anatómicos (Filgueiras et al. 2001) y moleculares (Duvall et al. 2001, Giussani et al. 2001). Duvall et al. (2001) y Giussani et al. (2001) comentan que Digitaria es uno de los géneros derivados dentro de la tribu Paniceae, a diferencia de lo expuesto por Clayton & Renvoize (1986), quienes lo ubicaron como uno de los más basales dentro de la tribu.
Digitaria se distribuye, en su mayoría, en regiones tropicales y subtropicales, con algunos representantes en regiones templadas de ambos hemisferios; en el continente americano se distribuye desde los Estados Unidos de América y las Antillas hasta la provincia de Río Negro (Argentina) y sus especies crecen principalmente por debajo de los 1000 m de altitud, aunque algunas pueden alcanzar ca. 3500 m. De las 170-330 especies conocidas para el género, 101 habitan en el Nuevo Mundo (Zuloaga et al. 2003) y 55 están en Sudamérica (Killeen & Rúgolo de Agrasar 1992) y de éstas, 18 especies están en Colombia, constituyendo el 7,5% de las especies de la tribu Paniceae presentes en el territorio colombiano (Giraldo-Cañas 2001a). En su trabajo sobre las Paniceae de Colombia, Pinto-Nolla (1997) sólo había registrado ocho especies de Digitaria para el país.
El presente aporte pretende contribuir al conocimiento de la rica flora de Colombia y en especial al de su diversa aunque escasamente conocida flora agrostológica, estimada en 153 géneros (Giraldo-Cañas & Mayorga 2001) y unas 751 especies (Giraldo-Cañas, observaciones personales). Esta monografía hace parte de los estudios de gramíneas colombianas que viene adelantando el autor desde hace algunos años (García-Ulloa et al. 2005, Giraldo-Cañas 1998, 1999a, 1999b, 2000a, 2000b, 2000c, 2001a, 2001b, 2002a, 2002b, 2002c, 2002d, 2003a, 2004a, 2004b, 2004c, Giraldo-Cañas & Mayorga 2001, Peñaloza-Jiménez et al. 2002).
MATERIALES Y MÉTODOS
Estudios exomorfológicos
La terminología de las estructuras morfológicas planas y tridimensionales está basada en los términos propuestos por Font Quer (1993), excepto en lo que tiene que ver con los términos “panoja” [= paniculodio sensu Rua (1999), panícula sensu Font Quer (1993)] y “espiguilla” [= florescencia sensu Rua (1999), espícula sensu Font Quer (1993)], siguiéndose en este caso a Vegetti & Pensiero (1993), ya que éstas son las formas adoptadas en la agrostología moderna. Por su parte, la terminología sexológica de las plantas está basada en Cocucci (1980), mientras que para la definición de antecio se sigue a McClure & Soderstrom (1972) y Zuloaga & Soderstrom (1985). Se sigue a Ellis (1979) para la terminología de las estructuras anatómicas, excepto en lo que tiene que ver con los microfitolitos (cuerpos de sílice), siguiéndose en este caso a Zucol (1996, 1998, 2000) y para referirse en forma conjunta a los aguijones y ganchos, se empleó el término “asperezas” según el criterio sustentado por Aliscioni & Arriaga (1998), puesto que la distinción entre estas dos estructuras es arbitraria (Snow 1996). Además, Metcalfe (1960) sugirió que los aguijones son homólogos a los ganchos. El eje principal de la panoja corresponde a la longitud medida desde el punto de origen del racimo basal al punto de origen del último racimo. La medida de las espiguillas está dada por la longitud de la lema inferior y, en unos pocos casos por la longitud del antecio superior cuando éste es de mayor tamaño que la lema inferior. El color del antecio superior corresponde al color cuando éste alcanza su madurez.
Se estudiaron las colecciones de diferentes herbarios, tanto nacionales como del exterior. Asimismo, se realizaron varias salidas de campo a diferentes regiones del país con el fin de enriquecer las colecciones de gramíneas colombianas. Se sigue a Rúgolo de Agrasar (1974) para la delimitación infragenérica de Digitaria, pues considera unas secciones más naturales, con excepción de Digitaria sect. Phaeotrichae Rúgolo, siguiéndose en este caso a Killeen & Rúgolo de Agrasar (1992), quienes retoman Digitaria sect. Calvulae (Stapf) Henrard. Las obras originales de las descripciones de las especies aquí consideradas se tomaron de la bibliografía disponible, del “International Plant Name Index Query IPNI” (www.ipni.org), de la base de datos “W3-TROPICOS” del Jardín Botánico de Missouri (EE.UU, www.mobot.org) y del Catalógo de las gramíneas del Nuevo Mundo (Zuloaga et al. 2003). Véanse Veldkamp (1973), Rúgolo de Agrasar (1974), Webster (1983) y Wipff & Hatch (1994) para un completo panorama de la historia nomenclatural del género Digitaria.
En ocasiones se detectaron mezclas de especies en una misma recolección. En tales casos se consideraron el recolector, el número de recolección y el número consecutivo de herbario [e.g. J. Betancur et al. 355 (COL: 327701 y HUA: 35388)]. Véanse Bor (1956), Rosengurtt et al. (1970), Veldkamp (1973), Rúgolo de Agrasar (1974), Pohl (1980), Webster (1983, 1987), Renvoize (1984, 1998), Judziewicz (1990), Tovar (1993), Pohl & Davidse (1994), Poilecot (1999), Vega & Rúgolo de Agrasar (2002a) y Zuloaga et al. (2003) para una lista actualizada de los sinónimos de las especies aquí tratadas.
Véanse Chase & Luces de Febres (1972), Nicora & Rúgolo de Agrasar (1987), Dávila Aranda & Manrique Forceck (1990), Judziewicz (1990), Dávila Aranda et al. (1993) y Pinto-Escobar (2002) para un completo glosario de la morfología de gramíneas. Para los aspectos tipológicos de las inflorescencias véanse Gram (1961), Cámara Hernández & Rua (1991), Vegetti & Pensiero (1993), Rua & Weberling (1995), Vegetti & Anton (1995, 1996), Rua & Boccaloni (1996), Vegetti & Weberling (1996), Rua & Gróttola (1997), Rua (1999, 2003), Giraldo-Cañas (1999b, 2000b) y Cámara Hernández (2001). Para la información concerniente a la variación estructural de las espiguillas en la tribu Paniceae se recomienda ver a Cialdella & Vega (1996), Vega & Rúgolo de Agrasar (2001) y Giraldo-Cañas (2003b).
Estudios micromorfológicos y anatómicos del antecio superior y la espiguilla
El antecio se define como la casilla floral formada por la lema y la pálea, dentro de la cual se halla una única flor (McClure & Soderstrom 1972), cuya función consiste en proteger la flor y fundamentalmente la formación de la cariopsis (véase Roth 1977) y, su estudio es un complemento importante para el conocimiento de las relaciones en la tribu Paniceae. Así, se abordó el estudio de la epidermis de la lema y la pálea superiores (antecio superior) por medio del microscopio electrónico de barrido (MEB), lo que permitió comparar el tipo de estructura y ornamento presentes en las especies de Digitaria. Este método ha sido utilizado con frecuencia en los últimos años en la tribu Paniceae, revelando importantes datos para la clasificación dentro de la tribu y a nivel subgenérico (Hsu 1965, Clark & Gould 1975, Rost & Simper 1975, Johnston & Watson 1981, Shaw & Smeins 1979, 1981, Shaw & Webster 1983, Renvoize & Zuloaga 1984, Zuloaga & Soderstrom 1985, Thompson & Estes 1986, Zuloaga 1985, 1987a, 1987b, Zuloaga et al. 1986, 1987, 1989, 1992, 1998, Arriaga 1987, Webster 1992, Zuloaga & Sendulsky 1988, Morrone & Zuloaga 1991a, 1991b, 1992, Filgueiras et al. 1993a, 1993b, 2001, Morrone et al. 1993, 1996, 1998, Cialdella et al. 1995, Zuloaga & Morrone 1996, Giraldo-Cañas 2000c, 2001b, 2002d, 2002e, 2003b, Costea & Tardif 2002). La morfología y el ornamento del antecio superior fueron destacados como dos de los caracteres más importantes a nivel genérico entre las Paniceae, pues éstos sirven para establecer relaciones inter e infragenéricas en esta tribu [véanse Chase (1911), Hsu (1965), Zuloaga & Soderstrom (1985) y Webster (1988, 1992)].
Las muestras para las observaciones con el microscopio electrónico de barrido, se obtuvieron de material seco de herbario. Las observaciones se realizaron en un microscopio ambiental electrónico de barrido Philips XL30-ESEM, perteneciente al Instituto de Investigación e Información Geocientífica, Minero, Ambiental y Nuclear “INGEOMINAS” de la ciudad de Bogotá. Las muestras se sometieron a una limpieza previa a la metalización sumergiéndolas en xileno y mediante la técnica de ultrasonido por espacio de 15 minutos, con lo que se logra, en la mayoría de los casos, la remoción de ceras epicuticulares e impurezas (véase Dávila & Clark 1990); luego las muestras se dejaron secar a temperatura ambiente por espacio de ocho horas. Posteriormente, éstas se metalizaron con una lluvia de oro en un metalizador Fisons-Polaron, modelo SC-500. Las microfotografías al MEB de las especies aquí tratadas pueden ser consultadas en Giraldo-Cañas (2004a). La anatomía foliar no se abordó en la presente investigación, pues hay numerosas contribuciones al respecto que involucran la mayoría de las especies presentes en el territorio colombiano (véanse Metcalfe 1960, Türpe 1966, Edwards & Black 1971, Mayne et al. 1971, Sangster 1977, Srivastava 1978, Webster 1983, 1987, Nicora & Rúgolo de Agrasar 1987, Rúgolo de Agrasar & Sánchez 1989, Watson & Dallwitz 1992, Krishnan 2000).
IMPORTANCIA ECONÓMICA
Digitaria constituye, junto con Axonopus P. Beauv., Panicum L., Paspalum L. y Setaria P. Beauv., un importante grupo no sólo a nivel económico, siendo algunos de sus representantes importantes forrajeras, sino también a nivel ecológico, dada su abundancia y diversidad en algunos ecosistemas (sabanas naturales, cerrados, afloramientos rocosos de los escudos precámbricos sudamericanos, áreas degradadas, áreas de cultivos de regiones montañosas como de sabanas y selvas basales). Digitaria constituye el tercer género más diverso en la tribu Paniceae, después de Panicum y Paspalum y uno de los géneros más ricos de las gramíneas.
Muchas de las especies de Digitaria son muy importantes, pues protegen el suelo de la erosión y ayudan a estabilizar y recuperar suelos de zonas inestables y con pronunciadas pendientes (Rúgolo de Agrasar 1974, Crins 1991, Watson & Dallwitz 1992), toda vez que constituyen elementos muy dinámicos en el proceso de la sucesión vegetal temprana. Algunas especies son empleadas como cereales por ciertas comunidades humanas (Veldkamp 1973, Rúgolo de Agrasar 1974, Watson & Dallwitz 1992). Otras son conocidas por sus cualidades como formadoras de céspedes (Veldkamp 1973). Por último, unas pocas especies constituyen plantas arvenses, las que pueden llegar a representar serios problemas en cultivos de arroz en algunas regiones colombianas (Prof. Cilia Fuentes, Facultad de Agronomía, Universidad Nacional de Colombia, sede Bogotá, com. pers.) y otras, en razón a la belleza de sus inflorescencias, pueden representar valiosos elementos ornamentales, principalmente las especies de Digitaria sect. Trichachne (Stapf) Henrard.
AFINIDADES GENÉRICAS
Digitaria tiene gran afinidad con los géneros Axonopus, Anthaenantia P. Beauv. (= Leptocoryphium Nees), Panicum y Paspalum de los que se diferencia fundamentalmente por tener la lema superior con los bordes membranáceos, generalmente hialinos y plegados sobre la pálea superior. Los citados géneros poseen la lema superior con los bordes cartilagíneos, al igual que su dorso y generalmente enrollados sobre la pálea superior. Digitaria se diferencia de Axonopus, pues en este género la lema superior se dispone con su dorso hacia afuera del raquis, mientras que en Anthaenantia, Digitaria, Panicum y Paspalum, la lema superior se dispone con su dorso contra el raquis (Giraldo-Cañas 2001b).
Por su parte, Anthaenantia y Digitaria son géneros que se confunden fácilmente, diferenciándose el primero básicamente por la ausencia de gluma inferior, por tener la gluma superior y la lema inferior isomorfas, por presentar el antecio superior abierto en la extremidad, dejando salir fácilmente la cariopsis y por su lema superior ciliada, en tanto que Digitaria presenta el antecio superior cerrado en su extremo, encerrando la cariopsis y su lema superior es glabra (Rúgolo de Agrasar 1974).
Igualmente, Digitaria y Reimarochloa Hitchc. son géneros fenéticamente muy similares. No obstante, Reimarochloa lleva glumelas poco endurecidas y se separa de Digitaria por una serie de caracteres derivados, tales como la ausencia de ambas glumas, la reducción de las piezas del microsporofiloceo (que es dímero), la soldadura basal de los estilos y el antecio superior abierto en el ápice (Watson & Dallwitz 1992).
Frecuentemente algunas especies de Digitaria son confundidas con miembros del género neotropical Mesosetum Steud. (observaciones personales con base en determinaciones erróneas en los herbarios visitados). Sin embargo, ambos géneros pueden reconocerse fácilmente por la naturaleza de las espiguillas (binadas o en grupos y conspicuamente pediceladas en Digitaria vs. solitarias y sésiles a subsésiles en Mesosetum), por la gluma inferior (enervia en Digitaria vs. 1-3-nervia en Mesosetum), por el antecio superior (cartilagíneo en Digitaria vs. coriáceo en Mesosetum), por la lema superior (de bordes membranáceos y cubriendo la pálea superior en Digitaria vs. de bordes coriáceos y aplicada sobre la pálea superior en Mesosetum) y por el tipo de hilo de la cariopsis (aovado u obovado en Digitaria vs. linear en Mesosetum) (véase Zuloaga & Morrone 1994: 200).
Ahora bien, Centrochloa Swallen, Spheneria Kuhlm. (géneros monotípicos de hábitats abiertos de Brasil), Axonopus (géneros que carecen de gluma inferior) y Digitaria son, por su parte, géneros muy relacionados, pero Centrochloa se diferencia por las modificaciones de su gluma superior, la que se extiende debajo del callo como una larga espuela cónica. En cuanto a Spheneria, su espiguilla es abaxial y su pálea superior es biaquillada, mientras que Axonopus y Centrochloa poseen espiguillas adaxiales y sus páleas superiores son enteras (Watson & Dallwitz 1992, Giraldo-Cañas 2001b). A continuación se presenta una clave para diferenciar los géneros similares a Digitaria.
Clave para diferenciar los géneros fenéticamente similares a Digitaria presentes en Sudamérica
1. Glumas ausentes; estambres 1 ó 2; espiguillas caedizas junto con el pedicelo.
Reimarochloa
1’. Glumas presentes o sólo la gluma superior presente; estambres 3; espiguillas caedizas sin el pedicelo.
2. Lema superior con el dorso cartilagíneo y los bordes membranáceos, hialinos a blanquecinos y planos, cubriendo casi completamente la pálea superior.
Digitaria
2’. Lema superior con el dorso y los bordes cartilagíneos o coriáceos, enrollados sobre la pálea superior.
3. Espiguillas con el dorso de la lema superior hacia fuera del raquis.
4. Gluma superior extendiéndose por debajo del callo como una larga espuela cónica.
Centrochloa
4’. Gluma superior sin ninguna extensión. Axonopus
3’. Espiguillas con el dorso de la lema superior hacia adentro del raquis.
5. Antecio superior abierto en su porción distal; inflorescencia con espiguillas no dispuestas en racimos unilaterales.
Anthaenantia lanata
5.’ Antecio superior cerrado en su porción distal.
6. Espiguillas sésiles a subsésiles; cariopsis con hilo linear.
Mesosetum
6’. Espiguillas pediceladas; cariopsis con hilo punctiforme a oblongo.
7. Pálea superior biaquillada. Spheneria
7.’ Pálea superior entera.
8. Espiguillas plano-convexas, dispuestas en racimos espiciformes; pálea inferior ausente.
Paspalum
8’. Espiguillas biconvexas, dispuestas en racimos no espiciformes; pálea inferior presente.
Panicum
RESULTADOS Y DISCUSIÓN
Caracteres exomorfológicos
Hábito. Las plantas pueden ser anuales o perennes, cespitosas o rizomatosas a raramente estoloníferas [D. eriantha Steud., D. fuscescens (J. Presl) Henrard], de porte grácil a robusto y con cañas simples o ramificadas y erectas o decumbentes. No obstante, el determinar la longevidad de las especies tiene varios inconvenientes, pues los ejemplares pueden estar incompletos o mal herborizados y en otros casos, la longevidad puede ser una respuesta ecológica, por lo que individuos de una misma especie pueden ser anuales o perennes, situación exhibida por otros miembros de la tribu Paniceae (Giraldo-Cañas 2001b). Es por esto que aquí no se considera la longevidad como un carácter importante para determinar ejemplares.
En varias especies las bases de las cañas y los rizomas están “protegidos” por catafilos, los que son por lo regular corta y densamente pilosos. Esta situación es particularmente frecuente en los miembros de Digitaria sect. Trichachne. El número de entrenudos por caña presenta una gran variación y va desde tres hasta trece o un poco más, siendo mayor en D. nuda Schumach. Por su parte, las vainas pueden variar de glabras a pilosas, en las que los pelos son unicelulares (macropelos) de paredes engrosadas y asociados en su base con células epidérmicas sobreelevadas con respecto al resto de la epidermis [= pelos de base tuberculada o papilosa, sensu Rúgolo de Agrasar (1974) y Snow (1996)].
Por su parte, la lígula puede variar de membranácea a escariosa y a su vez puede ser ciliada o glabra, y su borde puede ser entero, irregular, eroso, denticulado o laciniado (Fig. 1). Las láminas siempre son lanceoladas o lineares, planas, glabras o pilosas, en las que los pelos son de la misma naturaleza de los de las vainas. La abundancia o densidad del indumento no es tenida en cuenta aquí para fines de separación de las especies, pues es bien sabido que su variación en densidad corresponde a una respuesta ecológica a determinadas condiciones ambientales y no a una particularidad del genotipo. Ahora bien, la densidad del indumento puede variar al interior de un mismo ejemplar, siendo por lo regular las hojas de las porciones basales de las cañas más pilosas que las hojas distales. Es por esto que aquí no se le da importancia taxonómica a la densidad del indumento.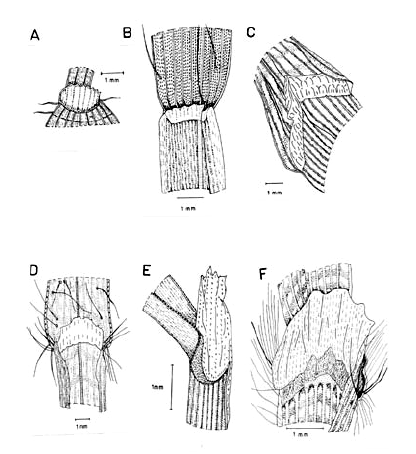
Figura 1. Tipos de lígula en las especies colombianas de Digitaria. A. Digitaria fuscescens (J. Presl) Henrard (Davidse & Llanos 5126, COL). B. Digitaria violascens Link (Giraldo-Cañas 3433, COL). C. Digitaria lehmanniana Henrard (Fuentes & Amaya 764, COL). D. Digitaria eriantha Steud. (Dugand 5900, COL). E. Digitaria similis Beetle ex Gould (Etter et al. 610, COL). F. Digitaria insularis (L.) Fedde (Giraldo-Cañas 3296, COL).
Inflorescencia. En general, la inflorescencia en las especies colombianas de Digitaria corresponde a una panoja abierta y exerta con las espiguillas adpresas a los racimos, y estos últimos no se ramifican en órdenes consecutivos (Fig. 2), o sea, los racimos son de primer orden, excepto en D. insularis (L.) Fedde, en la que varios ejemplares muestran que algunos racimos proximales pueden ramificarse en órdenes consecutivos, situación novedosa para Digitaria sect. Trichachne, ya que la presencia de racimos de segundo orden sólo se había documentado para Digitaria laxa (Rchb.) Parodi (Rua 2003). La inflorescencia presenta un eje principal desarrollado, en el que los racimos se disponen en forma verticilada o alterna, ya bien sea cercanos entre sí a conspicuamente distanciados, o nulo, caso en el cual los racimos son conjugados o digitados, como es el caso de D. longiflora (Retz.) Pers., y algunos ejemplares de D. andicola Giraldo-Cañas, D. fuscescens, D. sanguinalis (L.) Scop. y D. violascens Link. Por su parte, el número de racimos por panoja presenta una gran variación (uno a numerosos), incluso en una misma especie, por lo que su importancia taxonómica es nula.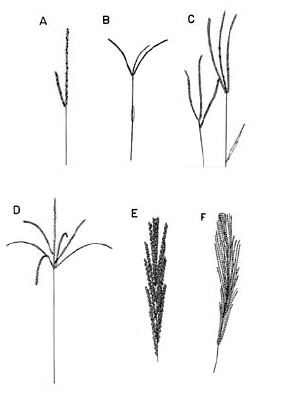
Figura 2. Tipos de inflorescencia en las especies colombianas de Digitaria. A. Digitaria andicola Giraldo-Cañas (Giraldo-Cañas 3319, COL). B. Digitaria fuscescens (J. Presl) Henrard (Davidse & Llanos 5126, COL). C. Digitaria violascens Link (Giraldo-Cañas 3433, COL). D. Digitaria eriantha Steud. (Dugand 5900, COL). E. Digitaria insularis (L.) Fedde (Giraldo-Cañas 3296, COL). F. Digitaria lehmanniana Henrard (Fuentes & Amaya 764, COL).
Raquis. Por lo regular, el raquis es glabro a glabrescente o escabroso a escabriúsculo. No obstante, una de las especies (D. horizontalis Willd.) presenta el raquis con escasos pelos unicelulares hialinos y de base tuberculada como los exhibidos por los representantes de las secciones Cabrera, Lappagopsis y Senescentia del género Axonopus (Giraldo-Cañas 2000c, 2001b, 2002e, 2003b) y por algunos miembros de Panicum (Zuloaga et al. 1992). Por otra parte, el raquis puede ser áptero y triquetro como sucede en la mayoría de las especies o alado como en D. fuscescens, D. longiflora y D. violascens (Fig. 3). A diferencia de las estructuras antes mencionadas, la forma del raquis sí tiene importancia taxonómica, pues cada especie sólo presenta un tipo de raquis.
Figura 3. Tipos de raquis y pedicelos en las especies colombianas de Digitaria. A. Digitaria similis Beetle ex Gould (Etter et al. 610, COL). B. Digitaria eriantha Steud. (Dugand 5900, COL). C. Digitaria lehmanniana Henrard (Fuentes & Amaya 764, COL). D. Digitaria andicola Giraldo-Cañas (Giraldo-Cañas 3319, COL). E. Digitaria insularis (L.) Fedde (Giraldo-Cañas 3296, COL). F. Digitaria violascens Link (Giraldo-Cañas 3433, COL).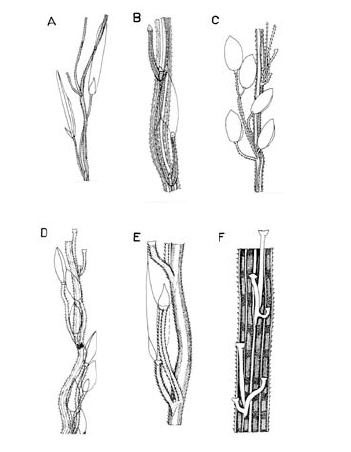
Pedicelo. Los pedicelos se disponen en el raquis en grupos de dos, tres, cuatro o cinco por nudo y los de un mismo nudo presentan longitudes diferentes (Fig. 3). Éstos pueden ser subtrígonos, trígonos o filiformes a capilares, rectos a flexuosos, glabrescentes a más comúnmente escabriúsculos o escabrosos, raras veces escasamente pilosos (D. horizontalis). La porción distal (articulación pedicelo-espiguilla) puede ser truncada (Digitaria sect. Digitaria) o discoide a cupuliforme en el resto de las especies. En las especies que presentan raquis alado (D. fuscescens, D. longiflora, D. violascens), el pedicelo de la espiguilla distal de cada nudo presenta su porción basal adnata al raquis.
Espiguilla. Las espiguillas son bifloras, aunque funcionalmente unifloras, pues la flor inferior está representada únicamente por la lema inferior y, en algunos casos por la pálea inferior y a menudo se observan una o dos lodículas. Éstas están dispuestas en dos hileras alternas sobre dos lados del raquis y son comprimidas dorsiventralmente, y su forma puede variar desde lanceolada, elíptica hasta ovada, y su porción distal puede ser aguda o acuminada (Fig. 4 y Fig. 5). Las espiguillas pueden ser glabras, glabrescentes o pilosas, situación en la cual el indumento puede ser muy variable, tanto en densidad, distribución y tipo de pelos, por lo que esta característica tiene mucha importancia taxonómica para separar especies y/o grupos de especies. Más adelante se hablará en detalle acerca de la naturaleza del indumento y de los tipos de pelo. El tamaño de las espiguillas también varía considerablemente, encontrándose las más pequeñas en Digitaria sect. Ischaemum Ohwi y Digitaria sect. Ternatae Hack. (1.2-2.5 mm de longitud), mientras que las más grandes se encuentran en Digitaria sect. Trichachne (3-5.1 mm de longitud), siendo intermedias en Digitaria sect. Calvulae y Digitaria sect. Digitaria (2-3.6 mm de longitud).

Figura 4. Tipos de espiguilla y lema inferior en las especies colombianas de Digitaria. A. Digitaria andicola Giraldo-Cañas (Giraldo-Cañas 3319, COL). B. Digitaria insularis (L.) Fedde (Giraldo-Cañas 3296, COL). C. Digitaria sanguinalis (L.) Scop. (Archer 3315, COL). D. Digitaria violascens Link (Giraldo-Cañas 3433, COL). E. Digitaria lehmanniana Henrard (Fuentes & Amaya 764, COL). F. Digitaria cardenasiana Giraldo-Cañas (Rangel & Espina 1418, COL).
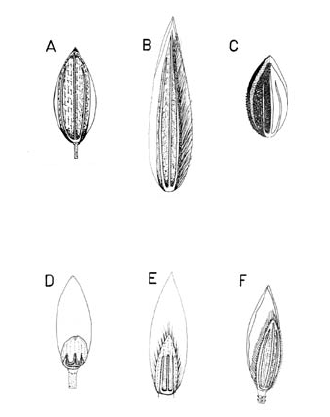
Figura 5. Tipos de gluma superior en las especies colombianas de Digitaria. A. Digitaria fuscescens (J. Presl) Henrard (Davidese 5126, COL). B. Digitaria insularis (L.) Fedde (Giraldo-Cañas 3296, COL). C. Digitaria cardenasiana Giraldo-Cañas (Rangel & Espina 1418, COL). D. Digitaria andicola Giraldo-Cañas (Giraldo-Cañas 3319, COL). E. Digitaria sanguinalis (L.) Scop. (Archer 3315, COL)). F. Digitaria eriantha Steud. (Dugand 5900, COL).
La gluma inferior puede estar ausente (D. longiflora, D. setigera Roth ex Roem. & Schult.) o ser muy reducida (Fig. 4), caso en el cual es enervia y glabra y presenta una textura membranácea a escariosa y su porción distal varía de triangular a truncada, emarginada o redondeada. Por su parte, la gluma superior puede ser tan larga como la lema inferior o más corta que ésta (Fig. 5). En las especies de Digitaria sect. Digitaria, la gluma superior presenta dimensiones mucho más cortas que la lema inferior, por lo regular ca. ½ o menos que ésta, mientras que en las otras secciones, generalmente, la gluma superior es tan larga o casi tan larga como la lema inferior. La gluma superior es de la misma naturaleza de la de la lema inferior y su diferencia fundamental radica en el número de nervios [menor en la gluma superior, (0--) 3 (--5)-nervia] y en la longitud. Los nervios pueden ser pronunciados a débiles y algunas veces no llegan al extremo distal de la gluma superior, caso en el cual son anastomosados hacia su porción distal, como en algunas espiguillas de D. fuscescens, D. lehmanniana Henrard y D. violascens. En la mayoría de los casos, la longitud de la lema inferior es análoga a la longitud del antecio superior (Fig. 4), salvo en algunas especies en las que la lema inferior puede ser ligeramente más corta o más larga que éste (ca. 0.2 mm). El número de nervios de la lema inferior oscila entre 5 y 7 y son similares a los de la gluma superior, salvo en D. sanguinalis, en la que los nervios son escabrosos en la porción distal de la lema inferior.La distancia de las piezas que componen la espiguilla sobre la raquilla, es más conspicua en los miembros de Digitaria sect. Trichachne que en las demás secciones. En esta sección, las glumas, la lema inferior y el antecio superior se insertan sobre la raquilla distanciados entre sí, por lo que los internodios son notables, tal como lo habían detallado Rúgolo de Agrasar (1974) y Webster (1983).
Naturaleza de los pelos de la espiguilla. Los pelos pueden ser bicelulares (= micropelos) y unicelulares (= macropelos). Los micropelos están presentes en todas las especies, aunque son muy escasos y éstos corresponden al tipo panicoide, donde la célula basal es más corta que la distal (Ellis 1979, Shaw & Smeins 1981, Thompson & Estes 1986, Snow 1996). Éstos se distribuyen en forma irregular tanto en la gluma superior como en la lema inferior. Por su parte, los macropelos son más interesantes dada su variación y sólo se encuentra un tipo de macropelo por especie, por lo que tienen una gran importancia taxonómica. Éstos pueden ser muy cortos (Digitaria sect. Digitaria, Digitaria sect. Ternatae), largos (Digitaria sect. Digitaria, Digitaria sect. Ternatae, Digitaria sect. Ischaemum, Digitaria sect. Calvulae) a muy largos (Digitaria sect. Trichachne), acuminados, agudos o con el ápice redondeado (Digitaria sect. Calvulae, Digitaria sect. Digitaria y Digitaria sect. Trichachne) o claviformes (Digitaria sect. Ternatae) tanto corta (D. cardenasiana Giraldo-Cañas) como largamente estipitados [D. andicola, D. filiformis (L.) Koeler y D. lehmanniana], o verrugosos y largos (Digitaria sect. Ischaemum), o sea, aquellos que presentan la superficie con pequeñas prominencias.Los macropelos se localizan en la gluma superior y en la lema inferior (nunca en la gluma inferior), aunque su disposición puede ser sólo en las márgenes de éstas o en toda su superficie o en los espacios internervales. Éstos varían en densidad, desde escasos hasta numerosos, constituyendo un indumento muy denso. En todos los casos, los macropelos no presentan una base tuberculada como sucede con los macropelos del raquis de D. horizontalis o los de las hojas de las especies de Digitaria. Caso contrario exhiben algunos representantes de géneros de la tribu Paniceae como Anthaenantiopsis Mez ex Pilger, Axonopus sect. Lappagopsis, Gerritea Zuloaga, Morrone & Killeen, Ichnanthus P. Beauv., Panicum y Paspalum, en los que los macropelos de las espiguillas presentan base tuberculada (Zuloaga & Sendulsky 1988, Morrone et al. 1993, Rúgolo de Agrasar & Nicora 1993, Zuloaga et al. 1993, 1994, Giraldo-Cañas 2002e, 2003b). Sin embargo, los macropelos en Axonopus y Panicum siempre son largos, lisos, agudos y no son dilatados, mientras que en Paspalum los puede haber como los anteriores de Axonopus y Panicum o capitados a clavados (véanse Veldkamp 1973, Davidse & Zuloaga 1992, Rodríguez et al. 2000, Giraldo-Cañas 2002e, 2003b).
En este sentido, el indumento de la espiguilla de Digitaria, cuando éste es simple (Digitaria sect. Calvulae, Digitaria sect. Digitaria y Digitaria sect. Trichachne), presenta más afinidades con Anthaenantia lanata (Kunth) Benth. [= Leptocoryphium lanatum (Kunth) Nees] (espiguillas pilosas, los pelos simples), y cuando el indumento está constituido por macropelos claviformes (Digitaria sect. Ternatae) se acerca más a Paspalum (Paspalum grupo Parviflora, pelos capitados). Por último, el indumento compuesto de macropelos verrugosos (Digitaria sect. Ischaemum) es poco frecuente en la familia y particularmente escaso en la tribu Paniceae, según la amplia bibliografía consultada. Los macropelos verrugosos exhibidos por las especies de Digitaria sect. Ischaemum son semejantes a los presentados por tres especies de Coelachyrum Hochst. & Nees de la subfamilia Chloridoideae, denominados por Snow (1996) como macropelos crispados.
Espiguillas trifloras. Se observaron espiguillas con dos antecios fértiles y uno neutro en D. dioica Killeen & Rúgolo [ejemplares Córdoba et al. 1511 (COL), Escobar 402 (VALLE)] y D. nuda [ejemplar Echeverry 2551 (COL)] (Tabla 1). En D. dioica, la única espiguilla de esta naturaleza fue observada en el tercio superior de un racimo distal, mientras que en D. nuda, estas espiguillas son numerosas y están distribuidas en todas las inflorescencias. Así, esta situación corresponde al caso de espiguillas trifloras, donde el antecio inferior está representado por la lema inferior o por la lema inferior, la pálea inferior y las lodículas inferiores reducidas (antecio inferior neutro, flor inferior neutra), más las dos flores de los antecios fértiles. Las espiguillas con dos antecios fértiles se asemejan a las propias del género Isachne R. Brown de la tribu Isachneae de las Panicoideae (Hsu 1965, Palacios 1968, Connor 1987, Kellogg & Campbell 1987, Zuloaga 1987a, Cialdella & Vega 1996), considerado como el “precursor” de la tribu Paniceae.
Tabla 1. Tipos de espiguillas trifloras en la tribu Paniceae y distribución de la sexualidad de las flores. (1): tomado de Morrone & Zuloaga (1989); (2): tomado de Cialdella & Vega (1996); (3): tomado de Morrone & Zuloaga (1991b); (4): tomado de Giraldo-Cañas (2003b); (5): el presente estudio.
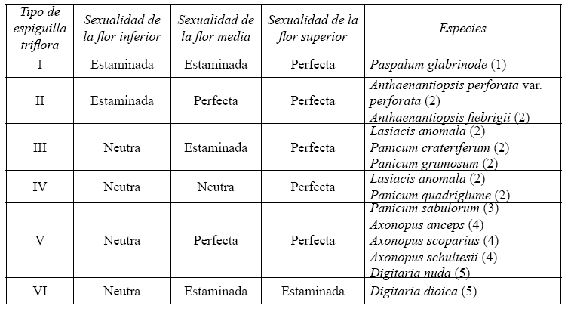
Por lo tanto, D. dioica y D. nuda son las únicas especies en el género que presentan espiguillas trifloras. Así pues, éstos son otros casos de espiguillas trifloras registrados en la tribu Paniceae. La presencia de espiguillas trifloras sólo ha sido citada para las especies Anthaenantiopsis perforata (Nees) Parodi var. perforata, Anthaenantiopsis fiebrigii Parodi, Axonopus anceps (Mez) Hitchc., Axonopus scoparius (Flüggé) Kuhlm., Axonopus schultesii G. A. Black, Panicum crateriferum Sohns, Panicum quadriglume (Döll) Hitchc., Panicum grumosum Nees, Panicum sabulorum Lam., Paspalum glabrinode (Hack.) Morrone & Zuloaga y para el género Lasiacis (Griseb.) Hitchc. (Morrone & Zuloaga 1989, 1991b, Cialdella & Vega 1996, Giraldo-Cañas 2003b). Con el hallazgo de estas espiguillas se adiciona un sexto tipo de espiguillas trifloras en la tribu Paniceae (véanse Morrone & Zuloaga 1989, 1991b, Cialdella & Vega 1996, Giraldo-Cañas 2003b). Véase la Tabla 1 para la distribución de los tipos sexuales de las flores en las espiguillas trifloras en la tribu Paniceae.
Antecio superior. Su color en la madurez varía de pajizo (secciones Calvulae, Digitaria, Ischaemum, Trichachne) a castaño o negruzco (Digitaria sect. Ternatae y D. violascens de Digitaria sect. Ischaemum) pasando por grisáceo o azuloso (e.g. algunos ejemplares de D. sanguinalis) a cobrizo [e.g. algunos ejemplares de D. californica (Benth.) Henrard y de D. insularis]. Algunas espiguillas de D. fuscescens aunque presentan el antecio superior pajizo, en ciertas ocasiones la porción distal de éste puede ser vinácea. Así, el color del antecio superior tiene importancia taxonómica a nivel de grupos o secciones.
En cuanto a la forma del antecio superior, éste puede variar de elíptico a lanceolado u ovado y de agudo a acuminado o apiculado (Fig. 6). Por otra parte, su consistencia es cartilagínea y su textura está caracterizada por papilas (papilas aparentemente simples al estereomicroscopio), las cuales se disponen en filas longitudinales y presentan diferentes dimensiones o son más pronunciadas en unas especies, por lo que algunas texturas pueden aparecer más rugosas que otras. En todos los casos el antecio superior es glabro (Fig. 6), aunque visto a gran aumento, éste puede presentar algunas asperezas distales.
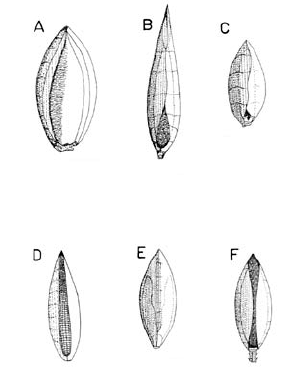
Figura 6. Tipos de antecio superior en las especies colombianas de Digitaria, vistos desde el lado de la pálea superior; nótense las diferentes proporciones de cubrimiento de la pálea superior por parte de la lema superior. A. Digitaria violascens Link (Giraldo-Cañas 3433, COL). B. Digitaria similis Beetle ex Gould (Etter et al. 610, COL). C. Digitaria cardenasiana Giraldo-Cañas (Rangel & Espina 1418, COL). D. Digitaria eriantha Steud. (Dugand 5900, COL). E. Digitaria lehmanniana Henrard (Fuentes & Amaya 764, COL). F. Digitaria andicola Giraldo-Cañas (Giraldo-Cañas 3319, COL).
Como se anotó anteriormente, el género Digitaria se caracteriza por presentar la lema superior con los bordes membranáceos, blanquecinos a más comúnmente hialinos y plegados sobre la pálea superior y cubriéndola casi completamente o cubriendo ca. 1/3-4/5 de ésta (Fig. 6). La lema superior es siempre enervia, excepto en D. nuda y D. similis Beetle ex Gould, en las que es 3-nervia (véase el capítulo de observaciones dado para estas especies).
La histología del antecio superior es muy interesante ya que sus células epidérmicas varían en forma y tamaño y su ornamento está constituído por diversas microestructuras. La epidermis abaxial de la lema y de la pálea superiores posee papilas, cuerpos de sílice, asperezas, aparatos estomáticos y puede presentar micropelos bicelulares. A continuación se destaca la naturaleza de diferentes estructuras presentes en el antecio superior.
Las células epidérmicas largas, tanto de la lema como de la pálea superiores, son rectangulares, más de tres a ocho veces más largas que anchas, agrupadas en hileras longitudinales, de paredes anticlinales longitudinales onduladas a marcadamente sinuosas (D. bicornis (Lam.) Roem. & Schult., D. californica, D. ciliaris (Retz.) Koeler, D. fuscescens, D. horizontalis, D. insularis, D. nuda, D. similis, D. violascens) o plegadas (D. andicola, D. cardenasiana, D. dioica, D. eriantha, D. filiformis, D. lehmanniana, D. longiflora, D. sanguinalis, D. setigera), con una (-3) papilas por célula.
Por su parte, las papilas son simples (aunque en D. californica éstas aparecen verrugosas, principalmente hacia la porción proximal del antecio superior), de tamaño variable, numerosas a muy numerosas y están dispuestas regularmente en hileras longitudinales sobre toda la superficie de la lema y de la pálea superiores, confiriéndole un aspecto rugoso a éstas. En todas las especies las papilas presentan su porción distal redondeada, excepto en D. californica, en la que las papilas de la porción proximal del antecio superior aparecen verrugosas como se indicó anteriormente. Por lo regular, las papilas están sobreelevadas con relación a la epidermis, aunque en D. filiformis, D. fuscescens y D. lehmanniana éstas aparecen ligeramente hundidas ya que se disponen en surcos longitudinales. Las papilas simples son la condición más común entre las Paniceae. No obstante, hay varias especies que presentan papilas verrugosas y/o compuestas en el antecio superior, las cuales son semejantes a las encontradas en D. californica (véanse Clark & Gould 1975, Shaw & Smeins 1981, Zuloaga & Soderstrom 1985, Thompson & Estes 1986, Morrone et al. 1998, Zuloaga et al. 1998).
Los cuerpos de sílice, aunque muy escasos, son circulares a redondeados. Éstos son exfoliados y se localizan principalmente en la porción distal de la lema superior. En la mayoría de las Paniceae, los cuerpos de sílice se localizan en la porción distal del antecio superior, tanto en la lema como en la pálea (Hsu 1965, Zuloaga & Soderstrom 1985, Giraldo-Cañas 2002d, 2003b). Algunas excepciones son comentadas por Morrone et al. (1996) para las especies Paspalum longicuspe Nash y Paspalum repens Berg., en las que los cuerpos de sílice están distribuidos sobre toda la superficie de la lema y la pálea superiores. Los cuerpos de sílice circulares a redondeados corresponden a la clase Pooideae (Zucol 1996) y es raro que para las especies de Digitaria aquí examinadas no se hallan encontrado cuerpos de sílice del tipo Panicoideae [halteriformes de varios tipos, los cuales son muy comunes en los representantes de la tribu Paniceae (véanse Zuloaga & Soderstrom 1985, Giraldo-Cañas 2002d, 2003b)], aunque éstos si fueron observados en la lámina foliar, en los pedicelos y en la gluma superior y la lema inferior de las especies colombianas de Digitaria.
Los micropelos pueden estar presentes o ausentes, cuando presentes éstos son bicelulares, fusiformes, lanceolados u oblongos, escasos a muy escasos y se localizan principalmente hacia la porción apical de la lema superior en dirección distal, y éstos se distribuyen irregularmente. Sus células distal y basal pueden estar o no colapsadas, y sus paredes son delgadas; la célula distal es de ápice ligeramente agudo a redondeado. Estos micropelos corresponden al tipo panicoide, donde la célula basal es más corta que la distal (Ellis 1979, Shaw & Smeins 1981, Zuloaga & Soderstrom 1985, Thompson & Estes 1986, Snow 1996, Giraldo-Cañas 2002d, 2003b).
Entre tanto, los macropelos (pelos unicelulares) no existen en el antecio superior de Digitaria, los que son frecuentes en varios miembros de las Paniceae (e.g. Axonopus, Panicum, Paspalum, entre otros, véase Giraldo-Cañas 2002d, 2003a, 2003b). Por tal razón, se dice que el antecio superior de Digitaria es glabro. No obstante, las especies aquí estudiadas presentan asperezas (aguijones y ganchos) en el extremo distal de la lema superior, por lo que se debería corregir la descripción genérica, toda vez que siempre se encuentra referida a que su antecio superior es glabro.
Cabe destacar que en la región proximal de la lema superior se halla diferenciada la porción de germinación, lugar por donde emerge la raíz embrionaria (Rost & Simper 1975, Johnston & Watson 1981, Shaw & Smeins 1981). La porción de germinación observada en las especies de Digitaria corresponde a la forma típica encontrada en las Paniceae (Rost & Simper 1975, Johnston & Watson 1981, Giraldo-Cañas 2002d), en las que la línea de ruptura es conspicua y semilunar. En ninguna de las especies estudiadas se observó espolón (prolongación endurecida de la base de la lema, véase Morrone & Zuloaga 1992).
Ahora bien, en todas las especies examinadas se observaron aparatos estomáticos en los antecios superiores, aunque éstos son muy escasos, los cuales se ubican principalmente en el extremo distal de la lema superior y éstos son paracíticos y superficiales o pueden estar ligeramente sobreelevados con relación a la epidermis. Las células subsidiarias son en forma de domo a subtriangulares, condición común entre las Panicoideae (véanse Ellis 1979, Giraldo-Cañas 2001c, 2003b). La presencia de aparatos estomáticos en el antecio superior es hallada con cierta frecuencia en otros géneros de la tribu Paniceae, aunque la mayoría de las especies de esta tribu no los presentan (Giraldo-Cañas 2002d, 2003b). Es así como varios investigadores observaron aparatos estomáticos en varios miembros de Paniceae: Panicum subgén. Dichanthelium (Clark & Gould 1975), Eriochloa Kunth (Shaw & Smeins 1981), Ichnanthus (Shaw & Webster 1983), Panicum grupo Lorea (Renvoize & Zuloaga 1984), Ichnanthus grandifolius (Döll) Zuloaga & Soderstrom, Panicum aristellum Döll, Tatianyx arnacites (Trin.) Zuloaga & Soderstrom (Zuloaga & Soderstrom 1985), Acroceras zizanioides (Kunth) Dandy (Zuloaga et al. 1987), Panicum sect. Stolonifera (Zuloaga & Sendulsky 1988), Panicum sabulorum (Morrone & Zuloaga 1991b), Panicum sect. Laxa (Zuloaga et al. 1992) y en Brachiaria (Trin.) Griseb. y Urochloa P. Beauv. (Morrone & Zuloaga 1992). Con respecto a la presencia de aparatos estomáticos en el antecio superior de las Paniceae, Shaw & Webster [1983, citando a Shaw & Gould (1979)] comentan que este carácter es más común en las Paniceae C3. No obstante, Digitaria es un género C4. La presencia de aparatos estomáticos en las lemas puede ser un carácter simplesiomórfico, toda vez que las lemas son homólogas a las hojas (Dr. L. E. Mora-Osejo, Universidad Nacional de Colombia, com. pers.).
De acuerdo con las características epidérmicas, tanto de la lema como de la pálea superiores, Digitaria presenta un “dermotipo panicoide” (Metcalfe 1960, Shaw & Smeins 1981, Thompson & Estes 1986, Zuloaga et al. 1989, Zuloaga & Morrone 1992, Filgueiras et al. 1993b, 2001, Giraldo-Cañas 2002d, 2003b), el que se caracteriza por presentar células largas rectangulares de paredes onduladas, presencia de micropelos bicelulares de paredes delgadas del tipo panicoide y cuerpos de sílice halteriformes (en mayor proporción), aunque éstos últimos no fueron observados en el antecio superior de ninguna de las 18 especies aquí examinadas. No obstante, algunos cuerpos de sílice halteriformes se observaron en las láminas foliares y en la gluma superior y la lema inferior de varias especies (véase Giraldo-Cañas 2002a).
Flor. La flor superior de las Paniceae es siempre perfecta (algunas excepciones son comentadas en Connor 1981, 1987, Vega 1996), mientras que la flor inferior es imperfecta o neutra como en todas las especies del género Digitaria. Las flores imperfectas se originan por reducción de flores perfectas (Cocucci & Anton 1988). Estos autores plantean la hipótesis de que tal reducción es debida a la inhibición ejercida por la pálea sobre el ápice floral. Siguiendo este razonamiento, la existencia de flores neutras podría explicarse como el resultado de la inhibición extrema del ápice floral (Cialdella & Vega 1996). Esta inhibición trae como resultado una reducción en el número de flores funcionales de la espiguilla.
Así, las espiguillas bifloras (normales) del género Digitaria son funcionalmente unifloras, dado que la flor inferior es neutra o imperfecta. Por lo tanto, la flor superior es siempre perfecta (excepto en D. dioca, véanse la descripción y el capítulo de observaciones dados para esta especie) y está constituida por tres estambres (los cuales emergen apicalmente), por un ovario oblongo y glabro, el cual remata en dos estilos y dos estigmas plumosos. En cuanto a su biología reproductiva, las especies del género Digitaria son autocompatibles (Connor 1987).
Cariopsis. La cariopsis es oblonga y comprimida dorsiventralmente, presentando un escudete embrional de ½ de la longitud de la cariopsis o un poco menor y con un surco poco notable, mientras que el hilo puede ser aovado u obovado.
TRATAMIENTO TAXONÓMICO
Digitaria Haller, Hist. Stirp. Helv. 2: 244. 1768 (nom. cons.), emend. A. S. Vega & Rúgolo, Amer. J. Bot. 88 (9): 1671. 2001.
Valota Adans., Fam. Pl. 2: 495. 1763 [non Valota Dumort. (Amaryllidaceae), Anal. Fam. Pl.: 58. 1829], nom. rejic. Especie tipo: Valota insularis (L.) Chase [≡ Andropogon insularis L. = Digitaria insularis (L.) Fedde].
Syntherisma Walter, Fl. Carol. 76. 1788. Especie tipo: Syntherisma praecox Walter [≡ Digitaria praecox (Walter) Willd. = Digitaria sanguinalis (L.) Scop.].
Acicarpa Raddi, Agrostogr. Bras. 31. 1823 [non Acicarpha Juss. (Calyceraceae), Ann. Mus. Natl. Hist. Nat. 2: 347. 1803], nom. illeg. hom. Especie tipo: Acicarpa sacchariflora Raddi [= Milium hirsutum P. Beauv. = Trichachne sacchariflora Nees = Digitaria sacchariflora (Nees) Henrard].
Paspalum L. sect. Digitaria (Haller) Nees, Fl. Bras. Enum. Pl. 2: 20. 1829. Basónimo: Digitaria Haller.
Trichachne Nees, Fl. Bras. Enum. Pl. 2: 85. 1829. Especie tipo: Trichachne sacchariflora Nees [= Digitaria sacchariflora (Nees) Henrard].
Gramerium Desv., Mem. Soc. Agric. Angers 1: 165. 1831. Especie tipo: Gramerium convolutum Desv.
Panicum L. sect. Digitaria (Haller) Trin., Gram. Panic. 48, 76. 1826. Basónimo: Digitaria Haller.
Paspalum L. sect. Digitaria (Haller) Nees, Fl. Bras. Enum. Pl. 2: 20. 1829. Basónimo: Digitaria Haller.
Panicum L. subgen. Digitaria (Haller) A. Gray, Manual 611. 1848. Basónimo: Digitaria Haller.
Panicum L. sect. Trichachne (Nees) Steud., Syn. Pl. Glumac. 1: 38. 1855 [1853]. Basónimo: Trichachne Nees (= Digitaria Haller).
Eriachne Phil., Anales Univ. Chile 36: 207. 1870. Especie tipo: Eriachne rigida Phil. [= Digitaria californica (Benth.) Henrard].
Panicum L. ser. Digitaria (Haller) Benth., Fl. Austral. 7: 464. 1878. Basónimo: Digitaria Haller.
Panicum L. ser. Digitarieae (Haller) Benth., Fl. Austral. 7: 464. 1878. Basónimo: Digitaria Haller.
Panicum L. ser. Trichachne (Nees) Benth., Fl. Austral. 7: 464. 1878. Basónimo: Trichachne Nees (= Digitaria Haller).
Panicum L. subgen. Digitaria (Haller) Hack., Oesterr. Bot. Z. 51: 290. 1901. Basónimo: Digitaria Haller.
Leptoloma Chase, Proc. Biol. Soc. Wash. 19 (34): 191-192. 1906. Especie tipo: Leptoloma cognata (Schult.) Chase [≡ Panicum cognatum Schult. = Digitaria cognata (Schult.) Pilg.].
Paspalum L. subgen. Digitaria A. Camus, Notul. Syst. (París) 2: 216. 1912. Basónimo: Digitaria Haller.
Digitaria subsect. Trichachne (Nees) Stapf, Fl. Trop. Afr. 9: 424. 1919. Basónimo: Trichachne Nees.
Digitariopsis C. E. Hubb., Hooker’s Icon. Pl. 35: t. 3420. 1940. Especie tipo: Digitariopsis redheadii C. E. Hubb.
Plantas anuales o perennes, cespitosas o rizomatosas, raramente estoloníferas (D. eriantha, D. fuscescens). Cañas simples o ramificadas, erectas o postradas. Vainas abiertas, glabras o pilosas. Lígula membranácea a escariosa, ciliada o glabra, de borde entero a irregular, denticulado o laciniado. Láminas lanceoladas o lineares, planas, glabras o pilosas. Inflorescencias terminales, erectas, exertas; panojas laxas a densas, constituidas por (1--) 2 a numerosos racimos unilaterales, éstos conjugados, verticilados a más comúnmente alternos sobre un eje, los racimos sin ramificarse en órdenes consecutivos (excepto en algunas inflorescencias de D. insularis); raquis trígono, áptero o alado, de bordes escabrosos, raramente piloso (D. horizontalis), terminando en una espiguilla; articulación pedicelo-espiguilla truncada, discoide o cupuliforme. Espiguillas pediceladas, binadas, ternadas o en grupos de 4 ó 5 por nudo, raramente solitarias, los pedicelos de un mismo nudo de diferente longitud, dispuestas en dos hileras alternas sobre dos lados del raquis, comprimidas dorsiventralmente, ovadas, lanceoladas, elípticas, agudas o acuminadas, glabras o pilosas, entonces los pelos de diferente naturaleza; 2-floras (ocasionalmente se encuentran espiguillas trifloras, e.g. D. dioica y D. nuda), antecio inferior estéril, el superior fructífero; raquilla articulada por debajo de las glumas, cayendo en conjunto a la madurez; gluma inferior ausente o reducida, enervia, triangular, redondeada o truncada, membranácea o hialina, glabra; gluma superior tan larga como la espiguilla o mucho más corta que ésta, (0-) 3 (-5)-nervia, glabra o pilosa; lema inferior tan larga como la espiguilla, 5-7-nervia, glabra o pilosa; pálea inferior ausente o reducida; flor inferior no desarrollada, con 1 ó 2 lodículas rudimentarias libres o soldadas; antecio superior tan largo o más corto que la espiguilla, dorso aplicado contra el raquis, cartilagíneo, glabro aunque con asperezas en su porción distal (lema superior), papiloso, ovado, elíptico a lanceolado, acuminado, apiculado o agudo, pajizo, violáceo, castaño o negruzco a la madurez, lema superior enervia a raramente 3-nervia (D. nuda, D. similis), estriada (las papilas dispuestas en filas longitudinales), de márgenes membranáceas, blanquecinas a hialinas, plegadas sobre la pálea superior y cubriéndola casi completamente en la mayoría de los casos o cubriendo ca. 1/3-2/3 de ésta; lodículas 2, cuneadas, cordiformes o truncadas, nerviadas; flor superior perfecta, raro unisexual (D. dioica); estambres 3, emergentes apicalmente; ovario con dos estilos y estigmas plumosos. Cariopsis comprimida dorsiventralmente; escudete embrional ½ de la longitud de la cariopsis o un poco menor, surco poco notable, hilo aovado u obovado.
Especie tipo. Digitaria sanguinalis (L.) Scop. (= Panicum sanguinale L.).
Etimología. Del latín digitus (= dedo), aludiendo a la posición de los racimos en la inflorescencia.
Número cromósomico básico. x = 9, 15 y 17 (Watson & Dallwitz 1992: 313).
Tipo fotosintético. C4, Kranz del subtipo anatómico MS NADP-me (Hattersley 1987, Rúgolo de Agrasar & Sánchez 1989). La vaina mestomática es la que aloja los cloroplastos especializados que caracterizan a las plantas C4. Esta vaina es homóloga a la endodermis por presentar sus células las paredes engrosadas, resistentes al ácido sulfúrico concentrado y bandas de Caspary (Rúgolo de Agrasar & Sánchez 1989). Estos caracteres se observan en los haces vasculares primarios; los haces terciarios presentan una sola vaina. La posición de los cloroplastos especializados de la vaina Kranz es centrífuga (Rúgolo de Agrasar & Sánchez 1989).
Observaciones. Algunos miembros del género Digitaria (principalmente ejemplares de Digitaria sect. Digitaria) son frecuentemente confundidos con el género Cynodon A. Rich. de la subfamilia Chloridoideae. Dicha confusión la he verificado en determinaciones erróneas en diferentes herbarios y quizás, este error frecuente es debido a la similitud de sus inflorescencias, en las que los racimos (Digitaria) y las espigas (Cynodon) se disponen en uno o varios verticilos, mostrándose como inflorescencias digitadas o verticiladas. No obstante, ambos géneros se pueden diferenciar fácilmente, pues Cynodon posee espiguillas sésiles (pediceladas en Digitaria), comprimidas lateralmente (comprimidas dorsiventralmente en Digitaria), solitarias (binadas, ternadas o en grupos de cuatro o cinco por nudo en Digitaria) y carinadas (no carinadas en Digitaria), con desarticulación arriba de las glumas (por debajo de las glumas en Digitaria) y cuya raquilla se extiende por detrás de la pálea como una cerda desnuda o con un pequeño rudimento (raquilla sin prolongación en Digitaria).
Clave para separar las secciones de Digitaria presentes en Colombia
1. Espiguillas binadas en el raquis.
2. Espiguillas glabras o pilosas, en este último caso el indumento nunca es lanoso; componentes de la espiguilla separados por internodios muy breves.
Digitaria sect. Digitaria
2’. Espiguillas conspicuamente pilosas, el indumento lanoso; componentes de la espiguilla separados por internodios muy evidentes.
Digitaria sect. Trichachne
1’. Espiguillas ternadas o en grupos de cuatro o cinco por nudo en el raquis.
3. Espiguillas dioicas, pilosas, los pelos simples y acuminados.
Digitaria sect. Calvulae (Digitaria dioica)
3. Espiguillas perfectas, glabrescentes a pilosas, los pelos verrugosos o claviformes.
4. Raquis alado; espiguillas con pelos verrugosos.
Digitaria sect. Ischaemum
4’. Raquis triquetro; espiguillas con pelos claviformes. Digitaria sect. Ternatae
Clave para separar las especies de Digitaria sect. Digitaria presentes en Colombia
1. Gluma inferior ausente.
Digitaria setigera
1’. Gluma inferior presente.
2. Espiguillas de 1.5-2.5 mm de longitud.
3. Raquis y pedicelos pilosos, los pelos hialinos de base tuberculada; lema superior enervia.
Digitaria horizontalis
3’. Raquis y pedicelos glabros; lema superior 3-nervia. Digitaria nuda
2’. Espiguillas 2.7-3.6 mm de longitud.
4. Plantas perennes, fuertemente estoloníferas. Digitaria eriantha
4’. Plantas anuales, no estoloníferas.
5. Gluma inferior triangular.
6. Nervios de la lema inferior lisos; lema superior cubriendo ca. 1/3-2/3 de la pálea superior. Digitaria ciliaris
6’. Nervios de la lema inferior escabrosos hacia su porción distal; lema superior cubriendo casi completamente o ca. 4/5 de la pálea superior. Digitaria sanguinalis
5’. Gluma inferior no triangular (ápice redondeado).
Digitaria bicornis
Clave para separar las especies de Digitaria sect. Ischaemum presentes en Colombia
1. Antecio superior castaño oscuro a negruzco. Digitaria violascens
1’. Antecio superior pajizo.
2. Espiguillas glabras a glabrescentes; plantas estoloníferas.
Digitaria fuscescens
2’. Espiguillas conspicuamente pilosas; plantas densamente cespitosas.
Digitaria longiflora
Clave para separar las especies de Digitaria sect. Ternatae presentes en Colombia
1. Espiguillas 1.2-1.7 mm de longitud.2. Eje principal de la inflorescencia de 0.5-5 cm de longitud; panojas constituidas por 2-18 (-20) racimos; antecio superior castaño oscuro; lígula de 0.7-2 mm de longitud.
Digitaria filiformis
2’. Eje principal de la inflorescencia de 7-16 cm de longitud; panojas constituidas por numerosos racimos (ca. 40); antecio superior castaño claro; lígula ca. 2 mm de longitud.
Digitaria lehmanniana
1’. Espiguillas de 1.8-2.5 mm de longitud.
3. Espiguillas densamente pilosas; gluma superior 0-0.25 mm más corta que la lema inferior. Digitaria cardenasiana
3’. Espiguillas glabrescentes a ligeramente pilosas; gluma superior 0.5-1.4 mm más corta que la lema inferior.
Digitaria andicola
Clave para separar las especies de Digitaria sect. Trichachne presentes en Colombia
1. Espiguillas con indumento ocráceo. Digitaria insularis
1’. Espiguillas con indumento blanquecino a blanco-plateado.
2. Espiguillas de 3-3.5 mm de longitud; gluma inferior triangular; gluma superior triangular y tan larga como la lema inferior o ligeramente más corta que ésta; antecio superior ovado y 0.5-0.8 mm más corto que la lema inferior; lema superior enervia.
Digitaria californica
2’. Espiguillas de 4.1-5.1 mm de longitud; gluma inferior lanceolada; gluma superior lanceolada y ca. 1 mm más corta que la lema inferior; antecio superior lanceolado y tan largo como la lema inferior; lema superior 3-nervia. Digitaria similis
Descripción de las especies
Digitaria sect. Calvulae (Stapf) Henrard
1. Digitaria dioica Killeen & Rúgolo, Syst. Bot. 17 (4): 603. 1992. TIPO. Bolivia. Santa Cruz: Prov. Ñuflo de Chávez, Estancia Salta, 10 km S of Concepción, 500 m, 11 sep 1985, T. Killeen 1192 (holotipo F; isotipos CTES, ISC, US, no vistos, MO!, SI!). Fig. 7.
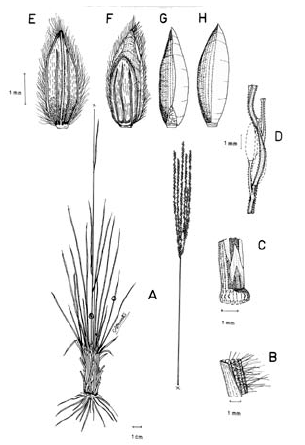
Figura 7. Digitaria dioica Killeen & Rúgolo: A. Hábito; B. Detalle del indumento de la lámina; C. Detalle de la lígula; D. Porción de un racimo; E. Espiguilla, vista desde la lema inferior; F. Espiguilla, vista desde la gluma superior; G. Antecio superior, visto desde la pálea superior; H. Antecio superior, visto desde la lema superior. Todo de Hermann 11091 (COL).
Plantas dioicas, perennes, moderada a densamente cespitosas; catafilos ausentes, muy raramente presentes, cuando presentes, escasos, membranáceos, con venación conspicuamente impresa, glabros, lustrosos, hasta de 12 mm de longitud; follaje denso, principalmente basal, formando engrosamientos basales a manera de bulbos. Cañas erguidas, simples, hasta de 60 cm de alto x ca. 1.5 mm de ancho; entrenudos regularmente 3, pajizos, lustrosos u opacos, lisos a levemente surcados, cilíndricos a canaliculados, hasta de 15 cm de longitud, siendo los entrenudos inferiores más cortos que los distales; nudos glabros, castaño-oscuros. Vainas hasta de 10 (-15) cm de longitud, las basales densamente híspidas, los pelos unicelulares, rectos, adpresos, ascendentes, suaves al tacto, hialinos o blancos a castaño-claros, las vainas distales haciéndose progresivamente menos pubescentes a glabras, las vainas seniles persistentes en la base de la planta, rotas a manera de flecos, las vainas maduras enteras, redondeadas. Lígula membranácea a escariosa, erosa, glabra, traslúcida, ca. 0.8 mm de longitud. Láminas hasta de 35 cm de longitud x ca. 3-4 mm de ancho (las láminas de las hojas de los nudos superiores reducidas), lineares, planas, ápice acuminado, glabras a pilosas, o sólo pilosas en su porción proximal, los pelos de base tuberculada. Inflorescencias erectas, largamente exertas; pedúnculos hasta de 40 cm de longitud; eje principal hasta de 6 cm de longitud; panojas constituidas por (2-) 3-4 (-6) racimos erectos, ascendentes, éstos hasta de 17 cm de longitud, próximos entre si; raquis triquetro, glabrescente, escabriúsculo marginalmente, ca. 0.3-0.4 mm de ancho; espiguillas densamente dispuestas en los racimos y en ocasiones constituyendo racimos cortos de segundo orden hasta de 2 (-3) cm de longitud, regularmente 3 espiguillas por nudo con pedicelos de diferente longitud; pedicelos glabrescentes, hasta de 5 mm de longitud los más largos, hasta de 2.5 mm los más cortos. Espiguillas 2.9-3.3 mm de longitud, ca. 0.9 mm de ancho, las espiguillas estaminadas y pistiladas similares, elipsoides; gluma inferior reducida, escariosa, translúcida, entera, enervia, de 0.2-0.5 mm de longitud, de ápice obtuso; gluma superior ca. 2/3-3/4 la longitud de la lema inferior, lanceolada, acuminada, 3-nervia, hirsuta entre los nervios y las márgenes, los pelos simples, acuminados, hialinos, adpresos o levemente divergentes, hasta de 1.5 mm de longitud; lema inferior de 2.9-3.3 mm de longitud, lanceolada, acuminada, 5-7-nervia, hirsuta, los pelos como los de la gluma superior, aunque generalmente un poco más largos y más numerosos, hasta 2 mm de longitud; pálea inferior ca. 0.4 mm de longitud, escariosa, hialina, uni-nervia; lodículas reducidas, apenas perceptibles, menores que la pálea inferior; antecio superior ca. 0.2-0.3 mm más corto que la lema inferior, elipsoide, acuminado, estriado, cartilagíneo, glabro, pajizo a castaño, las márgenes de la lema superior planas y encerrando la pálea superior; lodículas ca. 0.3 mm de longitud; espiguillas pistiladas con estaminodios blanquecinos, ca. 1.8 mm de longitud. Cariopsis ca. 1.7 mm de longitud, dorsiventralmente comprimida, pajiza a castaño-clara; escudete embrional ca. 0.7 mm de longitud; hilo oval, castaño oscuro, ca. 0.5 mm de longitud.
Distribución geográfica y ecológica. Digitaria dioica presenta una distribución disyunta y sólo es conocida de algunas sabanas y cerrados de Bolivia, Brasil y Colombia. Según Killeen & Rúgolo de Agrasar (1992), la distribución disyunta de esta especie y de otras gramíneas neotropicales restringidas a sabanas bien drenadas puede reflejar una expansión pleistocénica de las sabanas neotropicales. Dichos autores comentan que los estudios de los cambios pleistocénicos de la vegetación han girado en torno de especies amazónicas, y que las especies de sabanas pueden proveer datos adicionales para las hipótesis sobre cambios pleistocénicos de la vegetación.
Esta especie es localmente abundante en sabanas al pie de afloramientos rocosos. Crece en sabanas arboladas y no arboladas, junto con Byrsonima crassifolia Kunth (Malpighiaceae), Curatella americana L. (Dilleniaceae), Palicourea sp. (Rubiaceae), Sapium sp. (Euphorbiaceae), Clusiaceae spp., Xylopia sp. (Annonaceae), con diversas especies rupícolas de los géneros Vellozia (Velloziaceae), Navia, Aechmea (Bromeliaceae), Paspalum, Andropogon, Anthaenantia, Axonopus, Panicum, Setaria, Trachypogon (Poaceae), Bulbostylis (Cyperaceae), Syngonanthus, Paepalanthus (Eriocaulaceae), Drosera (Droseraceae), entre otras. Según se infiere de los diferentes ejemplares examinados, D. dioica siempre está asociada con sabanas sujetas a quemas frecuentes. En Colombia, D. dioica crece en los departamentos de Guaviare, Meta y Vichada, entre los 100 y 500 m de altitud.
Observaciones. Digitaria dioica es la única especie dioica en el género, característica que permite reconocerla fácilmente. En la tribu Paniceae la dioecia es muy rara y sólo se había registrado en tres géneros, los cuales están restringidos a Australia (véanse Connor 1981, Killeen & Rúgolo de Agrasar 1992, Connor et al. 2000): Pseudochaetochloa Hitchc., Spinifex L. y Zygochloa S. T. Blake.
Digitaria dioica es fácil y frecuentemente confundida con una especie del cerrado brasileño de Digitaria sect. Calvulae, D. neesiana Henrard. No obstante, ambas especies pueden reconocerse principalmente por su sexualidad [plantas dioicas (D. dioica) vs. plantas hermafroditas (D. neesiana)] y por el tamaño de sus espiguillas (2.9-3.3 mm de longitud en las poblaciones colombianas de D. dioica y 2.8-3.8 mm de longitud en las demás poblaciones sudamericanas de D. dioica vs. 4.0-5.5 mm de longitud en D. neesiana).
Por otra parte, D. dioica es frecuentemente confundida con la especie Anthaenantia lanata (= Leptocoryphium lanatum), situación que he podido verificar en las numerosas determinaciones erradas en los ejemplares de herbario. Sin embargo, estas dos especies pueden separarse fácilmente ya que A. lanata carece de gluma inferior, además tiene la gluma superior y la lema inferior isomorfas y presenta el antecio superior abierto en la extremidad, dejando salir fácilmente la cariopsis, siendo su lema superior ciliada, en tanto que D. dioica presenta el antecio superior cerrado en su extremo, encerrando la cariopsis y su lema superior es glabra.
Los ejemplares Córdoba et al. 1511 (COL) y Escobar 402 (VALLE) presentaron cada uno una espiguilla triflora, la cual estaba ubicada en el tercio superior de un racimo distal. Véase la Tabla 1 para ampliar la información y el significado de la presencia de espiguillas trifloras.
Ejemplares examinados
COLOMBIA. Guaviare: Mun. San José del Guaviare, serranía La Lindosa (Escudo Guayanés), inmediaciones de los afloramientos rocosos de “Ciudad de Piedra”, 250 m, nov 1995, D. Giraldo-Cañas & R. López 2506 (COAH, COL, SI). Meta: Llanos de Carimagua, 230 m, jun 1988, E. Escobar 402 (VALLE). Sabanas de El Vergel, feb 1937, H. García 4958-A (COL, paratipo). Al sur de la sierra de La Macarena, margen derecha del caño Losada, cerca de la confluencia con el río Guayabero, sabanas de La Cristalina, 1 km al S de La Peña de Los Conejos, cerca de la quebrada La Cristalina, 250 m, 11 mar 1959, R. Jaramillo Mejía & J. Hernández 2149 (COL). Vichada: Mun. Puerto Carreño, 16 km del Tapón, en altillanura disectada, 12 abr 1995, M. Córdoba et al. 1497 (COAH, COL). Mun. Puerto Carreño, vía a Puente Nuevo, en altillanura ondulada, 13 abr 1995, M. Córdoba et al. 1511 (COL). Territorio Faunístico El Tuparro, entre los ríos Tomo y Tuparro, El Tapón, 30 abr 1971, J. Daniel A-8 (COL, paratipo). 83 km E of Las Gaviotas, along unimproved dirt road to Santa Rita, below very small hills in sand, 150 m, 27 dic 1973, G. Davidse & F. Llanos 5240 (COL, paratipo). Along the Río Vichada at Bopimí, ca. 14 km NW de San José de Ocuné, dry llanos, 100 m, 24 ene 1944, F. Hermann 11091 (COL, paratipo).
Digitaria sect. Digitaria
2. Digitaria bicornis (Lam.) Roem. & Schult., Syst. Veg. 2: 470. 1817. Paspalum bicorne Lam., Tab. Encycl. 1: 176. 1791. TIPO. India, sin fecha, Sonnerat s.n. (holotipo P-LAM; isotipo US, no vistos). Fig. 8.
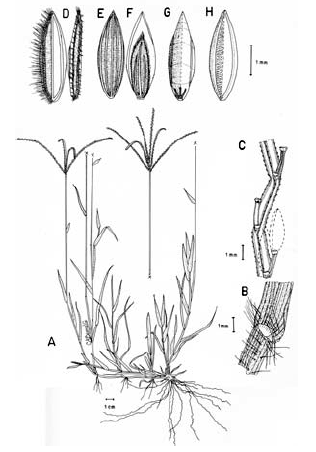
Figura 8. Digitaria bicornis (Lam.) Roem. & Schult.: A. Hábito; B. Detalle de la región ligular; C. Porción de un racimo; D. Detalle del indumento de la espiguilla en vistas frontal y lateral; E. Espiguilla, vista desde la lema inferior; F. Espiguilla, vista desde la gluma superior; G. Antecio superior, visto desde la lema superior; H. Antecio superior, visto desde la pálea superior. Todo de Romero-Castañeda 10358 (COL).
Plantas anuales, cespitosas, follaje laxo a denso, caulinar. Cañas simples o ramificadas, erectas a decumbentes en las porciones basales, caso en el cual las porciones decumbentes con raíces en los nudos inferiores y originando cañas floríferas ascendentes, cañas hasta de 60 cm de alto; entrenudos 3-9, pajizos a castaño claros, cilíndricos a canaliculados, hasta de 12 cm de longitud; nudos glabros o pilosos, castaño oscuros. Vainas hasta de 10 cm de longitud, aunque normalmente menores, glabrescentes a pilosas, principalmente hacia sus porciones proximal y distal, los pelos hialinos de base tuberculada. Lígula membranácea a escariosa, glabra, triangular o de ápice redondeado, éste entero o ligeramente dentado, 1-2.5 mm de longitud. Láminas hasta de 11 cm de longitud x 2-10 (-12) mm de ancho, planas, lanceoladas, ápice acuminado, glabrescentes a pilosas, los pelos de base tuberculada. Inflorescencias erectas, exertas a largamente exertas; pedúnculos hasta de 34 cm de longitud; eje principal de 0-3 cm de longitud; panojas constituidas por 2-8 racimos erectos, ascendentes, éstos hasta de 16 cm de longitud, raramente conjugados a alternos sobre el eje principal o dispuestos en 1 ó 2 verticilos; raquis triquetro, glabro, escabriúsculo marginalmente, 0.7-0.8 mm de ancho; espiguillas binadas, densamente dispuestas en los racimos, con pedicelos de diferente longitud, éstos trígonos, escabriúsculos, los más largos hasta de 3 mm de longitud, los más cortos hasta de 1 mm de longitud; articulación pedicelo-espiguilla truncada. Espiguillas 2.7-3.6 mm de longitud, lanceoladas, heteromorfas, caso en el cual la espiguilla conspicuamente pedicelada pilosa, la subsésil glabra a glabrescente, a homomorfas, entonces ambas espiguillas pilosas, los pelos simples, adpresos en las espiguillas jóvenes, patentes en las espiguillas muy maduras, hialinos a blanquecinos, brillantes, rectos a ligeramente flexuosos; gluma inferior glabra, enervia, membranácea, reducida, ca. 0.25 mm de longitud, de ápice redondeado; gluma superior de 1.7-2.4 mm de longitud, triangular, 3-nervia, pilosa; lema inferior de 2.7-3.6 mm de longitud, lanceolada, 5-7-nervia, pilosa hacia las márgenes; pálea inferior reducida, membranácea a escariosa; antecio superior ca. 0.3 mm más corto que la lema inferior, lanceolado, estriado, cartilagíneo, glabro, pajizo a castaño muy claro en la madurez, la lema superior con las márgenes membranáceas, hialinas a blancecinas y cubriendo ca. 1/3-4/5 de la pálea superior; lodículas ca. 0.25 mm de longitud, hialinas a pajizas, cordiformes. Cariopsis no vista.
Usos. Esta especie es muy apetecida como alimento por el chigüiro en los Llanos Orientales (Forero & Betancur 85, COAH).
Nombres vulgares. Guadín, guaduín, guardarrocío, guardarrocío colchado, hierba coneja (Tolima), mindaca (Sucre).
Distribución geográfica y ecológica. Digitaria bicornis se distribuye en las regiones tropicales y subtropicales de ambos hemisferios (Pohl & Davidse 1994), principalmente en áreas de cultivo de zonas bajas y en áreas degradadas con una fuerte intervención antrópica. En Colombia, esta especie está presente prácticamente en todo el país y se distribuye desde el nivel del mar hasta los 1800 m.
Observaciones. Digitaria bicornis presenta espiguillas heteromorfas, siendo la espiguilla conspicuamente pedicelada pilosa, mientras que la subsésil es glabra a glabrescente. Las espiguillas pilosas exhiben en su madurez los pelos en forma patente. La gluma inferior de esta especie presenta el ápice redondeado y una longitud de ca. 0.25 mm, mientras que las demás especies de Digitaria sect. Digitaria que poseen dicha estructura, ésta es triangular o truncada y de una longitud mucho mayor o menor a la de la gluma inferior de D. bicornis.
Ejemplares examinados
COLOMBIA. Antioquia: Mun. Dabeiba, 27 km después por la carretera al mar, 250 m, 28 sep 1986, J. Betancur et al. 265, 267 (HUA). Mun. Necoclí, El Totumo, carretera Turbo-Necoclí, km 41, 30 m, 29 sep 1986, J. Betancur et al. 282 (HUA). Mun. Necoclí, carretera Necoclí-Arboletes, km 42, 160 m, 29 sep 1986, J. Betancur et al. 300 (COL, HUA, MO). Mun. Valdivia, corregimiento Puerto Valdivia, 8 km de Puerto Valdivia, mina de oro “El 15”, 14 may 1987, R. Callejas et al. 3471 (HUA). Mun. Santo Domingo, vereda Piedras Gordas, quebrada Piedras Gordas, carretera Barbosa-Cisneros, a la altura de Pescadito Nro. 1, en inmediaciones de la finca de Don Manuel Arango-Rozzo, 1400-1500 m, 4 ene 2002, D. Giraldo-Cañas et al. 3303 (COL, HUA). Mun. Fredonia, La Botero, cerca de Puente Iglesias, 13 nov 1988, Grupo Medio Ambiente ISA 5 (HUA). Mun. Santa Fe de Antioquia, finca Cotové, 580 m, 24 sep 1986, R. Pohl 15595 (HUA). Mun. Betulia, La Barcaza, hacienda Cabildo, 19 nov 1987, Sección Medio Ambiente ISA 41 (HUA). Arauca: Mun. Arauca, Caño Limón, campo petrolero de la Occidental de Colombia, 230 m, 8 jun 1998, J. Forero & J. Betancur 67 (COAH), 9 jun 1998, J. Forero & J. Betancur 85 (COAH). Mun. Arauca, inmediaciones de las instalaciones de la Universidad Nacional de Colombia, hacienda El Cairo, formaciones de sabanas inundables con pequeñas “matas de monte”, carretera Arauca-Tame, km 9, 200-300 m, 13 jun 2003, D. Giraldo-Cañas et al. 3502, 3515 (COL). Atlántico: Usiacurí, arroyo Cagón, 100 m, 7 ene 1940, A. Dugand & H. García-Barriga 2358 (COL). Barranquilla, El Prado, 60 m, 20 dic 1958, A. Dugand 5072 (COL), 1 nov 1961, A. Dugand 5871 (COL), 8 mar 1962, A. Dugand 6070 (COL). Playa de Salgar, 0-5 m, 11 nov 1967, A. Dugand 7118 (COL). Puerto Colombia, 10 m, 19 oct 1963, P. Pinto 717 (COL). Bolívar: Cartagena, en las murallas, castillo de San Felipe, 17 ene 1941, A. Dugand & R. Jaramillo 2873 (COL). Cartagena, Bocagrande, 0-5 m, 15 oct 1984, J. Wood 4658 (COL). Golfo de Morrosquillo, archipiélago de San Bernardo, isla Múcura, 22 ago 1990, F. Flórez et al. 27 (COL). Vicinity of Turbaco, 200-300 m, 6-22 nov 1926, E. Killip & A. Smith 14190 (COL). Caldas: río Magdalena, km 224, 200 m, 3 mar 1961, J. Idrobo 4333 (COL). Casanare: entre Yopal y Aguazul, 400 m, año 1983, J. Wood 3828 (COL). Chocó: Puerto Pizarro, 2 km al N en el estero de José Ángel, río Baudó, 0-5 m, 3 ene 1967, H. Fuchs et al. 22148 (COL). Córdoba: Mun. Lorica, Estación Piscícola de la CVS, ca. 50 m, 22 feb 2005, Giraldo-Cañas et al. 3856 (COL). Mun. Montería, Aguas Negras, 12 km NE de Montería, 15 m, 1 feb 1970, B. Anderson 1909 (COL). Cundinamarca: Mun. Nariño, 340 m, 16 feb 1986, J. Fernández & R. Jaramillo 5214 (COL). Mun. Nilo, carretera Nilo-Melgar, en coluvios de los cortes de la carretera, ca. 200 m, 19 oct 2004, D. Giraldo-Cañas et al. 3793 (COL). Entre Viotá y Girardot, 320-560 m, ago 1964, C. Saravia 4626 (COL). Guaviare: Mun. San José del Guaviare, trocha Nuevo Tolima, en cercanías al Batallón José Joaquín París, 250 m, mar 1996, D. Giraldo-Cañas & R. López 2572 (COAH, SI). La Guajira: Mun. Barrancas, near Tabaco, El Cerrejón, 23 nov 1980, P. Bunch et al. 285 (HUA). Serranía La Macuira, entre el arroyo y la duna Arehuara, 290 m, 4 mar 1963, C. Saravia 2368 (COL). Magdalena: isla de Salamanca, en Los Cocos, 14 jul 1970, N. de López 414 (COL), 4 dic 1966, R. Romero-Castañeda 10535 (COL). Región de Santa Marta, Ciénaga, Aguacoca, 26 jul 1966, R. Romero-Castañeda 10358 (COL). Isla de Salamanca, cerca de La Tasajera, 17 ene 1967, M. Schnetter 51 (COL). Mun. Ciénaga, 14 may 1961, R. Romero-Castañeda 8903 (COL). San Andrés, Providencia y Santa Catalina: isla de Providencia, bahía Maracaibo, 18 sep 1984, J. Idrobo 11600 (COL). Santander: Region about Landázuri, 70 km N of Vélez, near Las Flores, 700 m, 9 jun 1944, N. Fassett 25304 (COL). Entre el caño de Chicamocha y el valle de Umpala, 1000 m, 14 jun 1962, C. Saravia 777 (COL). Carretera Piedecuesta-Pescadero, 1005 m, 18 jun 1962, C. Saravia et al. 976 (COL). Sucre: alrededores de Verrugas, finca La Aguada, 0-10 m, 21 ago 1979, H. Bernal 148 (COL). Mun. Tolú, arroyo Villero, carretera Tolú-Coveñas, km 20.5, litoral caribe, golfo de Morrosquillo, 0-5 m, 18 sep 1990, J. Betancur & M. Berrío 1987 (COL, HUA). Sin localidad, 137 m, 29 may 2003, D. Aldana 3 (COL). Tolima: Mun. El Espinal, vereda San Francisco, 438 m, 25 jul 1962, Instituto de Fomento Algodonero 11 (COL). Mun. Saldaña, finca Las Lagunas, 15 oct 1999, F. Montealegre s.n. (COL: 434863). Mun. El Espinal, vía El Espinal-Guamo, km 12, vereda Canasto, borde de lote de arroz, 400 m, 8 jul 2002, A. Osorrio et al. 166 (COL). Valle del Cauca: Cali, carrilera frente al edificio Unidad Vecinal Venezolana, 1000 m, 4 feb 1961, J. Idrobo 4212 (COL). Loboguerrero, 1050 m, 12-14 nov 1962, C. Saravia 1501 (COL). Vichada: Región Guayanesa, Mun. Puerto Carreño, afloramientos rocosos del tipo “lajas” entre Punta de Lajas y el cerro El Bita, ribera del río Orinoco, 40-100 m, 4-5 ene 2004, D. Giraldo-Cañas & C. Parra 3623, 3642, 3647 (COL).
3. Digitaria ciliaris (Retz.) Koeler, Descr. Gram. 27. 1802. Panicum ciliare Retz., Observ. Bot. 4: 16. 1786. Panicum sanguinale L. var. ciliare (Retz.) Vasey, U. S. Dep. Agr. Div. Bot. Bul. 8: 23. 1889. LECTOTIPO. China, Canton, Wennerberg s.n. (LD, no visto, designado por Veldkamp, Blumea 21: 32. 1973). Fig. 9.
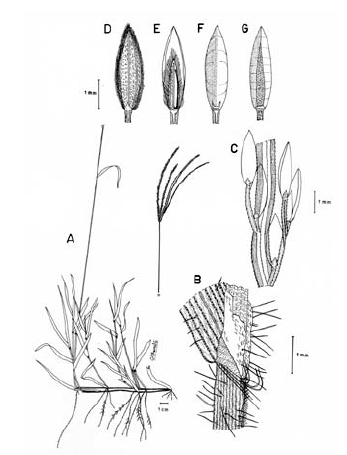
Figura 9. Digitaria ciliaris (Retz.) Koeler: A. Hábito; B. Detalle de la región ligular; C. Porción de un racimo; D. Espiguilla, vista desde la lema inferior; E. Espiguilla, vista desde la gluma superior; F. Antecio superior, visto desde la lema superior; G. Antecio superior, visto desde la pálea superior. Todo de Pohl 15548 (HUA).
Plantas anuales, cespitosas, follaje laxo a denso, caulinar. Cañas simples o ramificadas, erectas a decumbentes en las porciones basales, entonces originando raíces en los nudos inferiores y cañas ascendentes en éstos, cañas hasta de 80 cm de alto; entrenudos 3-11, pajizos a castaño claros, cilíndricos a canaliculados, hasta de 12 (-20) cm de longitud; nudos glabros o pilosos, castaño oscuros. Vainas hasta de 10 cm de longitud, aunque frecuentemente de menor longitud, glabrescentes a pilosas, entonces los pelos hialinos de base tuberculada. Lígula membranácea a escariosa, glabra, triangular, de borde finamente dentado, 1.5-2.9 mm de longitud. Láminas hasta de 18 cm de longitud x 2-9 mm de ancho, planas, lineares a lanceoladas, ápice acuminado, glabrescentes a pilosas en su porción proximal, los pelos de base tuberculada. Inflorescencias erectas, exertas a largamente exertas; pedúnculos hasta de 40 cm de longitud; eje principal de 0.2-7 cm de longitud; panojas constituidas por 3-9 (-12) racimos erectos, ascendentes, éstos hasta de 20 cm de longitud, subdigitados a alternos sobre el eje principal o dispuestos en 1-2 verticilos o ambas condiciones en una misma panoja; raquis triquetro, glabro, escabriúsculo marginalmente, 0.6-0.9 mm de ancho; espiguillas binadas, con pedicelos de diferente longitud, éstos trígonos, escabriúsculos, los más largos hasta de 2.5 mm de longitud, los más cortos hasta de 0.8 mm de longitud; articulación pedicelo-espiguilla truncada. Espiguillas 2.8-3.3 mm de longitud, lanceoladas, pilosas, los pelos simples y suaves, adpresos a levemente divergentes; gluma inferior glabra, enervia, membranácea, triangular, 0.4-0.5 mm de longitud; gluma superior de 1.8-2.2 mm de longitud, triangular, 3-nervia, pilosa; lema inferior de 2.8-3.3 mm de longitud, lanceolada, 5-7-nervia, nervios lisos, pilosa en las márgenes; pálea inferior ausente o escasamente visible; antecio superior tan largo como la lema inferior, lanceolado, estriado, cartilagíneo, glabro, pajizo a castaño muy claro, la lema superior con las márgenes membranáceas, hialinas a blanquecinas, plegadas sobre la pálea superior y cubriendo ca. 1/3-2/3 de ésta; lodículas ca. 0.3 mm de longitud, cordiformes. Cariopsis ca. 1.7 mm de longitud, comprimida dorsiventralmente; escudete embrional ca. ½ de la longitud de la cariopsis; hilo aovado.
Usos. Forragera, aunque puede constituir una maleza de primer grado en cultivos de tierras bajas.
Nombres vulgares. Alambrillo (Antioquia), grama alta (Santander), falsa pata de gallina (Tolima).
Distribución geográfica y ecológica. Digitaria ciliaris se distribuye en las regiones tropicales y subtropicales de ambos hemisferios (Pohl & Davidse 1994, Vega & Rúgolo de Agrasar 2002a). En Colombia, está presente prácticamente en todo el país y se distribuye desde el nivel del mar hasta los 2600 m. Esta especie es frecuente en terrenos modificados y en áreas de cultivo.
Observaciones. Digitaria ciliaris es frecuentemente confundida con D. horizontalis y con D. setigera. Pero el raquis y los pedicelos de D. ciliaris carecen de pelos de base tuberculada, característica propia de D. horizontalis. Además, D. ciliaris tiene espiguillas de mayor tamaño (2.8-3.3 mm de longitud vs. 1.5-2.5 mm en D. horizontalis) y su gluma inferior es triangular (truncada en D. horizontalis). De D. setigera se distingue fácilmente, pues ésta carece de gluma inferior. Véanse las observaciones dadas para D. sanguinalis.
Ejemplares examinados
COLOMBIA. Amazonas: corregimiento departamental La Pedrera, en el sitio denominado “Chorro de Córdoba”, zona de raudales del río Caquetá, en afloramiento rocoso del Escudo Guayanés, 230 m, 19 jul 1997, D. Giraldo-Cañas et al. 2720 (COAH, COL, HUA, SI). Antioquia: Fredonia, 1850 m, 3 ago 1930, W. Archer 504 (COL). Mun. Necoclí, sitio El Sucio, carretera Necoclí-Arboletes km 20, 80 m, 29 sep 1986, J. Betancur et al. 291 (COL, HUA, MO). Mun. Arboletes, orilla de playa, 30 sep 1986, J. Betancur et al. 311 (COL, HUA). Mun. Chigorodó, hacienda Montecristo, 25 km después de Apartadó, en cultivo de “chontaduro”, 40 m, 3 oct 1986, J. Betancur et al. 355 (COL: 327701). Grassland at Hacienda Montenegro near La Pintada, 600 m, 20 oct 1947, C. Blackman et al. 17C-283 (COL). Mun. Cáceres, El Doce, Bajo Cauca, 200-400 m, 25 mar 1978, R. Callejas 503 (COL, HUA, dos ejemplares). Yondó, 0-100 m, 28 jun 1989, M. de Fraume et al. 958 (COL). Mun. Itagüí, en jardín urbano, carrera 50 con calle 64, barrio Simón Bolívar, 1550 m, 16 jul 1992, D. Giraldo-Cañas 985 (HUA), 2 ene 2002, D. Giraldo-Cañas et al. 3292 (COL). Mun. Medellín, predios internos del jardín botánico “Joaquín Antonio Uribe”, 1550 m, 2 ene 2002, D. Giraldo-Cañas et al. 3294 (CAUP, COL, HUA). Mun. Caucasia, entre Caucasia y Nechí, por la troncal norte, ca. 20 km de Caucasia, hacienda La Candelaria, 17 ago 1986, R. Pohl et al. 15411 (HUA). Mun. Jardín, vereda Las Manguitas, 18 km al oeste de Jardín, 1600 m, 19 sep 1986, R. Pohl & J. Betancur 15548 (HUA, dos ejemplares). Bolívar: Mun. Turbaco, 12 ago 1980, J. Espina 522 (COL). Boyacá: Mun. Santa María, vereda La Esmeralda, km 1 entre Santa María y Juntas, cercanías al río Garagoa, 960 m, 25 mar 2001, J. Betancur et al. 9010 (COL). Caquetá: Quinche, 90 km dowstream of Araracuara, año 1977, M. Eden 48 (COL). Cauca: Mun. Popayán, predios internos y jardineras de la Facultad de Educación de la Universidad del Cauca, 1750 m, 7-11 nov 2004, D. Giraldo-Cañas et al. 3806 (COL). Córdoba: Montería, 25 m, 10 ago 1969, B. Anderson 1819 (COL). Mun. San Antero, cerro Buenos Aires, bosque seco secundario, 25 feb 2005, D. Giraldo-Cañas et al. 3861 (COL). Río San Jorge, margen izquierda, cerca de la ciénaga de Ayapel, 50 m, 12 jul 1973, J. Idrobo & A. Cleef 6606 (COL). Mun. Cereté, granja experimental Turipaná, jul 1971, F. Lozano 23 (COL). Cundinamarca: predios del jardín botánico “José Celestino Mutis”, 2550 m, 25 sep 1986, G. Morales et al. 697 (COL). Guaviare: Mun. San José del Guaviare, inspección La Carpa, vereda La Rompida, terrenos arenosos del río Guayabero, 250 m, 10 feb 1994, L. César 16 (COAH). Huila: a few kms SW of Gallego, 1400 m, 23 oct 1983, J. Wood 4053 (COL). Magdalena: Santa Marta, ca. 30 m, años 1898-1901, H. Smith 180 (COL); Santa Marta, años 1898-1901, H. Smith s.n. (tres ejemplares, COL: 253679, COL: 253824 y COL: 253850). Meta: 10 km de la carretera Villavicencio-Bogotá, 700 m, 13 jun 1989, F. Zuloaga 3948 (COL, dos ejemplares, SI). Nariño: Mun. Buesaco, 2 km antes de la población, 2000 m, 4 may 1989, B. Ramírez 1413 (COL, PSO). Carretera Túquerres-Pasto, cerca al río Juanambú, 1380-1820 m, 15 dic 1962, C. Saravia & R. Jaramillo 1893 (COL). Santander: Sabana de Torres, Aguas Claras, Salinas, 270 m, 17 nov 1985, J. Torres & P. Pinto 2823 (COL). Carretera Piedecuesta-Pescadero, 1005 m, 18 jun 1962, C. Saravia et al. 986 (COL). Coromoro, hacienda Los Arrayanes, 1300 m, 28 ene 1942, L. Wiedmam 2, 6 (COL). Sucre: Mun. San Onofre, corregimiento Las Brisas, carretera Las Brisas-Altos de Tulio, cuenca del arroyo Caricano, 6 sep 1996, A. Realpe 124 (HUA). Tolima: meseta de Ibagué, Picaleña, finca Corinto, 5 km vía Ibagué-El Espinal, 830 m, 11 jul 2002, A. Osorio et al. 268 (COL). Ibagué, San Jorge, 1250 m, 11 mar 1964, R. Russi 55 (COL). Valle del Cauca: Río Calima, margen derecha, playas en Pailón, 60 m, 23 may 1946, J. Cuatrecasas 21222 (VALLE).
4. Digitaria eriantha Steud., Flora 12: 468. 1829. TIPO. Sudáfrica, Cabo de la Buena Esperanza, von Ludwig s.n. (holotipo no localizado). Fig. 10.
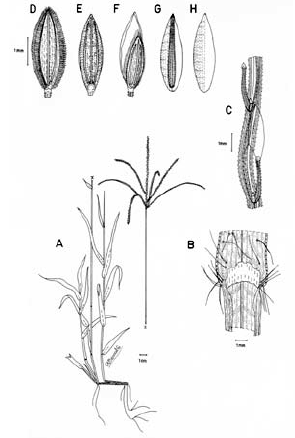
Figura 10. Digitaria eriantha Steud.: A. Hábito; B. Detalle de la región ligular; C. Porción de un racimo; D. Detalle del indumento de la espiguilla desde la lema inferior; E. Espiguilla, vista desde la lema inferior; F. Espiguilla, vista desde la gluma superior; G. Antecio superior, visto desde la pálea superior; H. Antecio superior, visto desde la lema superior. Todo de Dugand 5900 (COL).
Plantas perennes, cespitosas, estoloníferas, los estolones hasta de 1 m de longitud, follaje laxo. Cañas simples o ramificadas, erectas a decumbentes en las porciones basales, entonces originando raíces en los nudos inferiores y cañas ascendentes en éstos, cañas hasta de 100 cm de alto; entrenudos numerosos, pajizos a castaño claros, cilíndricos a canaliculados, hasta de 15 cm de longitud; nudos glabros, castaño oscuros. Vainas hasta de 12 cm de longitud, aunque frecuentemente de menor longitud, glabras, glabrescentes a finamente pubescentes, entonces los pelos hialinos de base tuberculada. Lígula membranácea a escariosa, glabra, triangular, con el extremo distal denticulado, 2-6 mm de longitud. Láminas hasta de 25 cm de longitud x 2-5 mm de ancho, planas, lineares a lanceoladas, ápice acuminado, glabras. Inflorescencias erectas, exertas a largamente exertas; pedúnculos hasta de 45 cm de longitud; eje principal de 0-3 (-4.5) cm de longitud; panojas constituidas por 3-12 racimos erectos, ascendentes, éstos hasta de 17 cm de longitud, subdigitados, digitados a alternos sobre el eje principal o dispuestos en 1-2 verticilos; raquis triquetro, escabriúsculo marginalmente, ca. 0.5 mm de ancho; espiguillas binadas, con pedicelos de diferente longitud, éstos trígonos, escabriúsculos, los más largos hasta de 1.6 (-2) mm de longitud, los más cortos hasta de 0.7 mm de longitud; articulación pedicelo-espiguilla truncada. Espiguillas 2.8-3.4 mm de longitud, lanceoladas, glabrescentes o cortamente pilosas, los pelos simples y suaves, adpresos a levemente divergentes; gluma inferior membranácea, glabra, enervia, triangular, 0.3-0.6 (-1) mm de longitud; gluma superior de 1.6-1.9 mm de longitud, triangular, 3-nervia, cortamente pilosa; lema inferior de 2.8-3.4 mm de longitud, lanceolada, 5-7-nervia, glabrescente a cortamente pilosa; pálea inferior ca. 0.2 mm de longitud, membranácea a escariosa; antecio superior tan largo como la lema inferior o ca. 0.25 mm más corta que ésta, lanceolado, estriado, cartilagíneo, glabro, pajizo a grisáceo, la lema superior con las márgenes membranáceas, hialinas a blanquecinas, plegadas sobre la pálea superior y cubriéndola casi completamente (ca. 4/5 de ésta); lodículas ca. 0.2 mm de longitud. Cariopsis no vista.
Usos. Esta especie es una destacada forrajera, principalmente para ganado vacuno (Mejía-Saulés 1986, Nicora & Rúgolo de Agrasar 1987, Pohl & Davidse 1994).
Nombres vulgares. Pangola (Antioquia, Atlántico, Cundinamarca, Magdalena).
Distribución geográfica y ecológica. Especie nativa de África, introducida en América por su importancia forrajera (Mejía-Saulés 1986, Beetle 1987, Nicora & Rúgolo de Agrasar 1987, Pohl & Davidse 1994). De Colombia sólo se tienen registros de Antioquia, Atlántico, Córdoba, Cundinamarca, Magdalena, Meta y Santander y crece entre el nivel del mar y los 1800 m.
Observaciones. De las especies de Digitaria sect. Digitaria presentes en Colombia, D. eriantha es la única que es perenne y fuertemente estolonífera. Estas características, sumadas a la longitud de su lígula (2-6 mm) y de sus glumas inferior (0.3-1 mm) y superior (1.6-1.9 mm), permite reconocerla fácilmente entre las especies de dicha sección.
Veldkamp (1973: 13) ubica a D. eriantha (tratada como D. pentzii Stent) en la sección Erianthae Henrard, quizás por su hábito perenne, mientras que la sección Digitaria está caracterizada por especies anuales (Webster 1987). No obstante, aquí se ha preferido considerarla en la sección Digitaria, ya que sus características morfológicas son compartidas con los representantes de dicha sección. Es por esto que el hábito anual o perenne no constituye un carácter taxonómico para separar entidades, pues se ha visto que incluso una misma especie puede presentar individuos anuales o perennes, como una estrategia para sobrevivir en un ambiente en particular. Por lo tanto, la longevidad de una planta es una respuesta ecológica que no puede ser usada como una característica diagnóstica (véase Giraldo-Cañas 2001b). Al respecto, Rúgolo de Agrasar (1974) ya había considerado a D. eriantha (tratada como D. decumbens Stent) en la sección Digitaria, lo que concuerda con la posición aquí adoptada. Digitaria eriantha aparece frecuentemente tratada en obras botánicas de diferente índole como D. decumbens o como D. pentzii, binomios considerados sinónimos de ésta (véase Zuloaga et al. 2003: 201).
Ejemplares examinados
COLOMBIA. Antioquia: granja experimental “Tulio Ospina”, Medellín, año 1971, C. Franco s.n. (HUA: 3501). Mun. Medellín, calle 48D, cerca de Almacenes Éxito, 1450 m, 24 ago 1986, R. Pohl 15433 (HUA, MO). Atlántico: Los Pendales, hacienda “Riodulce”, en dehesas, 10-15 m, 19 nov 1961, A. Dugand 5900 (COL). Córdoba: Mun. Cereté, granja experimental Turipaná, jul 1971, F. Lozano 19 (COL). Cundinamarca: Fusagasugá, 18 jul 1961, J. Idrobo 4647 (COL, dos ejemplares). Silvania, club El Bosque, 1300 m, año 1983, J. Wood 3852 (COL). Magdalena: Santa Marta, parque nacional Tairona, río Piedras, 7 m, dic 1991, P. Torrijos & Martínez 142 (COL). Meta: Puerto Gaitán, 150 m, 20 jun 1995, S. Lægaard & C. Mayorga 17617 (COL). Santander: ca. 2 km E of Suratá, towards California, 1900 m, 14 sep 1985, J. Wood 5058 (COL).
5. Digitaria horizontalis Willd., Enum. Pl. 92. 1809. TIPO. Hispaniola. Santo Domingo, Anón. (holotipo B-W; isotipos MVFA, US, no vistos). Fig. 11.
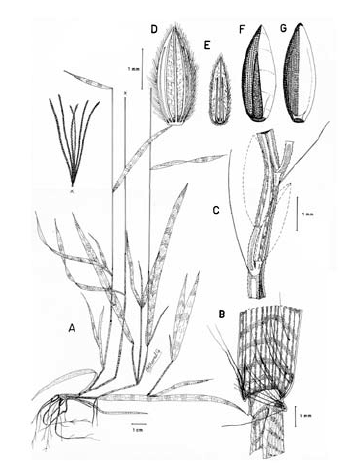
Figura 11. Digitaria horizontalis Willd.: A. Hábito; B. Detalle de la región ligular; C. Porción de un racimo (nótense los macropelos unicelulares en los racimos); D. Espiguilla, vista desde la lema inferior; E. Detalle de la gluma superior; F. Antecio superior, visto desde la pálea superior; G. Antecio superior, visto desde la lema superior. Todo de Instituto de Fomento Algodonero 48 (COL).
Plantas anuales, cespitosas, follaje laxo a denso, caulinar. Cañas simples o ramificadas, erectas a decumbentes en las porciones basales, entonces originando raíces en los nudos inferiores y cañas ascendentes en éstos, cañas hasta de 45 cm de alto; entrenudos 3-9, pajizos a castaño claros, cilíndricos a canaliculados, hasta de 10 cm de longitud; nudos glabros o pilosos, castaño oscuros. Vainas hasta de 9 cm de longitud, aunque frecuentemente de menor longitud, glabrescentes a pilosas, entonces los pelos hialinos de base tuberculada. Lígula membranácea a escariosa, glabra, obtusa, con el extremo distal denticulado, 1.2-1.9 mm de longitud. Láminas hasta de 11 cm de longitud x 2-9 mm de ancho, planas, lanceoladas, ápice acuminado, glabrescentes a pilosas en su porción proximal, los pelos de base tuberculada. Inflorescencias erectas, exertas a largamente exertas; pedúnculos hasta de 25 cm de longitud; eje principal de 0-2.5 (3) cm de longitud; panojas constituidas por (2-) 3-10 racimos erectos, ascendentes, éstos hasta de 15 cm de longitud, conjugados, subdigitados, digitados a alternos sobre el eje principal o dispuestos en 1-2 (4) verticilos; raquis triquetro, laxamente piloso, los pelos hialinos, tiesos y de base tuberculada, escabriúsculo marginalmente, 0.5-0.7 mm de ancho; espiguillas binadas, con pedicelos de diferente longitud, éstos trígonos, escabriúsculos, escasamente pilosos, los pelos hialinos, tiesos y de base tuberculada, pedicelos más largos hasta de 2.5 mm de longitud, los más cortos hasta de 0.7 mm de longitud; articulación pedicelo-espiguilla truncada. Espiguillas (1.5) 2-2.5 mm de longitud, lanceoladas, pilosas, los pelos simples y suaves, adpresos a levemente divergentes; gluma inferior glabra, enervia, reducida a un borde membranáceo, truncada, ca. 0.1 mm de longitud; gluma superior de 1-1.5 mm de longitud, triangular, 3-nervia, pilosa; lema inferior de (1.5) 2-2.5 mm de longitud, lanceolada, 5-7-nervia, pilosa marginalmente; pálea inferior no vista; antecio superior tan largo como la lema inferior o ca. 0.25 mm más corta que ésta, lanceolado, estriado, cartilagíneo, glabro, pajizo a castaño muy claro, la lema superior enervia, con las márgenes membranáceas, hialinas a blanquecinas, plegadas sobre la pálea superior y cubriéndola casi completamente; lodículas ca. 0.2 mm de longitud, truncadas. Cariopsis ca. 1.5 mm de longitud, comprimida dorsiventralmente; escudete embrional ca. ½ de la longitud de la cariopsis; hilo aovado.
Usos. Forraje.
Nombres vulgares. Guardarrocío, guadín, guaduín, hierba coneja (Huila, Tolima), conejo, pasto conejo (Valle del Cauca).
Nombres indígenas. Kanapiri (lengua Guahiba, Casanare).
Distribución geográfica y ecológica. Digitaria horizontalis se distribuye en las regiones tropicales de ambos hemisferios (Pohl & Davidse 1994). Vive en suelos arenosos y húmedos y en terrenos modificados y en áreas de cultivo. En Colombia, esta especie está presente prácticamente en todo el país y se distribuye desde el nivel del mar hasta los 2600 m.
Observaciones. Esta especie es fácilmente reconocible, pues es la única que presenta el raquis y los pedicelos laxamente pilosos, siendo los pelos tiesos, hialinos y de base tuberculada. Además, D. horizontalis presenta espiguillas pequeñas, de (1.5) 2-2.5 mm de longitud. Estas dos características permiten separarla fácilmente de las demás especies de Digitaria sect. Digitaria. Véanse las observaciones dadas para D. ciliaris y D. nuda.
Ejemplares examinados
COLOMBIA. Amazonas: Trapecio Amazónico, orilla del río Loretoyacu, 250 m, nov 1945, J. Duque-Jaramillo 2066 (COL). Quebrada Aduche, en yucal, 200 m, 5 ago 1977, J. Idrobo 8901 (COL). Río Caquetá, Araracuara, 1 abr 1976, C. Sastre & H. Reichel 5084 (COL). Atlántico: entre Baranoa y Galapa, 120-140 m, 4 nov 1961, A. Dugand 5878 (COL). Bolívar: islas del Rosario, mar Caribe, 2 m, 14-17 oct 1963, P. Pinto 711, 714 (COL). Caldas: Mun. Arauca, granja experimental Monte Lindo, 1200 m, 27 feb 1973, D. Soejarto 3896 (HUA, dos ejemplares). Casanare: Los Llanos, río Casanare, Esmeralda, en sabana, 130 m, 20 oct 1938, J. Cuatrecasas & H. García-Barriga 3887 (COL). margen derecha del río Casanare, caserío indígena Tsamani, 100 m, 28 jul 1977, F. Ortiz 266 (COL). Cundinamarca: near La Mesa on the road to El Colegio, 1000 m, año 1983, J. Wood 3589 (COL). Guaviare: Mun. San José del Guaviare, inspección La Carpa, vereda La Rompida, en la vega del río Guayabero, 250 m, 10 feb 1994, L. César 19 (COAH). Huila: Mun. El Agrado, quebrada La Yaguilga, campos de cacao, 700 m, sep 1986, J. Fernández & G. Morales 6886 (COL). Magdalena: sin localidad, 100 m, oct 1861, M. Lindig 1126 (COL). Santa Marta, parque nacional Tairona, río Piedras, 5 m, dic 1991, P. Torrijos & Martínez 27 (COL). Meta: río Guayabero, raudal Angostura Nro. 1, en las playas del río, 350 m, 20 ene 1959, P. Pinto 279 (COL). Nariño: Túquerres, 2500 m, may 1853, J. Triana s.n. (COL: 2009). Quindío: Mun. Quimbaya, rastrojos abiertos en los límites con cultivos de café, 1500 m, 20-25 mar 2005, D. Giraldo-Cañas & J. García-Ulloa 3881 (COL). Mun. Quimbaya, Parque Principal, 1500 m, 20-25 mar 2005, D. Giraldo-Cañas & J. García-Ulloa 3890 (COL). Mun. Circasia, Parque Principal, 1500 m, 20-25 mar 2005, D. Giraldo-Cañas & J. García-Ulloa 3897 (COL). Tolima: Armero, campo experimental Bledonia, 28 abr 1960, J. Barreneche 16 (COL). Mun. Armero, vereda Bledonia, 421 m, 16 ago 1962, Instituto de Fomento Algodonero 48 (COL). El Espinal, estación agrícola Nataima, 400 m, 10 may 1967, L. Jeffery 1 (COL). Mun. Saldaña, vereda Normandía, lote La Monterredondo, 17 abr 2002, J. L. Álvarez 11 (COL). Mun. Ibagué, barrio especial El Salado, lote Los Tres Carlos, 23 abr 2002, J. L. Álvarez 19 (COL).Mun. Saldaña, finca Las Lagunas, 15 oct 1999, F. Montealegre s.n. (COL: 434862). Mun. El Espinal, finca experimental Dupont, vereda Guayabal, variante El Espinal-Ibagué, al lado del seminario Diocesano, 320 m, 11 jul 2002, A. Osorio et al. 296 (COL). Valle del Cauca: Estación Piedras, entre Obando y Cartago, 980 m, 5 feb 1961, J. Idrobo 4241 (COL). Mun. Cali, Ingenio Meléndez, 1008 m, 18 jul 1966, J. Páez 7 (COL). Palmira, granja agrícola experimental, 5 ene 1957, A. Ramírez 11 (COL). Vaupés: río Kuduyarí, Pacuativa, 380 m, H. García-Barriga 14903 (COL, dos ejemplares). Río Piraparaná, tributary of Río Apaporis, headwaters of Caño Teemeeña, 10 sep 1952, R. Schultes & I. Cabrera 17385 (COL). Vichada: caño Urimica, 19 dic 1971, I. Cabrera 1686 (COL). Sin departamento ni localidad: años 1783-1808, J. Mutis 5456 (COL, dos ejemplares).
6. Digitaria nuda Schumach., Beskr. Guin. Pl. 45. 1827. TIPO. Ghana, P. Thonning 367 (holotipo C; isotipo L, no vistos). Fig. 12.
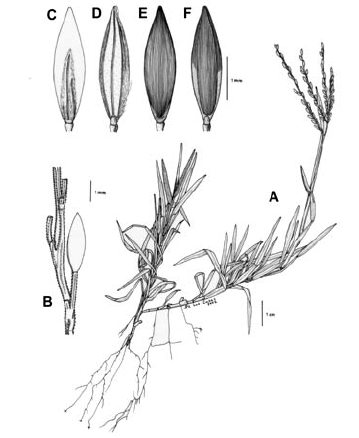
Figura 12. Digitaria nuda Schumach.: A. Hábito; B. Porción de un racimo; C. Espiguilla, vista desde la gluma superior; D. Espiguilla, vista desde la lema inferior; E. Antecio superior, visto desde la lema superior; F. Antecio superior, visto desde la pálea superior. Todo de Echeverry 2551 (COL).
Plantas anuales, gráciles, cespitosas, follaje denso, caulinar. Cañas ramificadas, erectas a decumbentes en la base, caso en el cual las porciones decumbentes hasta de 10 cm de longitud y con raíces en los nudos inferiores y originando numerosos tallitos ascendentes, cañas hasta de 15 cm de alto; entrenudos numerosos (entre 6 y 13 por caña sin considerar los de las porciones decumbentes), pajizos a castaño oscuros, lustrosos u opacos, glabros, lisos, cilíndricos a canaliculados, hasta de 2 cm de longitud aunque normalmente de 1 cm de longitud; nudos glabros, castaño oscuros. Vainas hasta de 4.5 cm de longitud, aunque normalmente no superan los 2.5 cm de longitud, pilosas con pelos de base tuberculada, principalmente en su porción distal. Lígula mambranácea a escariosa, glabra, levemente erosa, 1.5-2 mm de longitud. Láminas hasta de 4 cm de longitud x 2-3 mm de ancho, planas, lanceoladas, ápice acuminado, glabras a corta y laxamente pubescentes o sólo pilosas cerca a la región ligular. Inflorescencias erectas, largamente exertas; pedúnculos hasta de 10 cm de longitud; eje principal de 0.4-0.7 cm de longitud; panojas constituidas por 2-4 racimos erectos, ascendentes, éstos hasta de 4.3 cm de longitud, alternos en el eje principal o dispuestos en dos verticilos; raquis triquetro, glabro, escabriúsculo marginalmente, 0.5-0.6 mm de ancho; espiguillas densamente dispuestas en los racimos y binadas, raramente solitarias en las porciones basales de los racimos, con pedicelos de diferente longitud, éstos subtrígonos, escabriúsculos, los más largos hasta de 4 mm de longitud, los más cortos hasta de 1.3 mm de longitud; articulación pedicelo-espiguilla truncada; presencia de numerosas espiguillas trifloras con dos antecios perfectos y uno neutro. Espiguillas (2.3-) 2.5 mm de longitud, lanceoladas, pilosas, los pelos simples, agudos, tenues, flexuosos; gluma inferior glabra, enervia, membranácea a escariosa, truncada a obtusa, 0.25-0.4 mm de longitud; gluma superior 0.6-1.7 mm de longitud, triangular, 3-nervia, pilosa, los pelos tenues y simples; lema inferior de (2.3-) 2.5 mm de longitud, membranácea, lanceolada, 5-7-nervia, los nervios conspicuos, pilosa en las porciones marginales, los pelos simples y tenues; pálea inferior no observada; antecio superior tan largo como la lema inferior, lanceolado, acuminado a cortamente apiculado, estriado, cartilagíneo, glabro, pajizo o raramente castaño muy claro, la lema superior 3-nervia, con las márgenes membranáceas y cubriendo ca. 1/3 de la pálea superior; lodículas ca. 0.25 mm de longitud, cordiformes a rectangulares con la porción distal erosa, hialinas a pajizas. Cariopsis no vista.
Distribución geográfica y ecológica. Digitaria nuda es una especie tropical de ambos hemisferios, especialmente frecuente en África (Vega & Rúgolo de Agrasar 2002a). En el continente americano había sido registrada en Guayanas, Brasil, Bolivia, Paraguay, Argentina (Renvoize 1984, 1998, Rúgolo de Agrasar 1994, Poilecot 1999, Vega & Rúgolo de Agrasar 2002a), Estados Unidos de América, Canadá (Webster 1987), México, Measoamérica y Antillas (Webster & Hatch 1981, Pohl & Davidse 1994). Esta especie crece en terrenos húmedos y arenosos de tierras bajas. En Colombia sólo ha sido registrada una vez en la costa pacífica del Valle del Cauca.
Observaciones. Las inflorescencias del único ejemplar examinado presentan numerosas espiguillas trifloras con dos antecios perfectos y uno neutro (véase el capítulo “Caracteres exomorfológicos” para ampliar la información y el significado de las espiguillas trifloras). Así, esta especie, junto con D. dioica, son las únicas en el género que presentan espiguillas trifloras. Por otra parte, la gluma superior en las espiguillas trifloras es más pequeña que la de las espiguillas normales (bifloras).
Esta especie presenta la lema superior 3-nervia, la cual, además, sólo cubre ca. 1/3 de la pálea superior, lo que permite diferenciarla fácilmente de las demás especies cercanas. Digitaria nuda es afín a D. horizontalis. No obstante, éstas pueden distinguirse fácilmente, pues en la primera de ellas el raquis y los pedicelos carecen de pelos de base tuberculada, los cuales son conspicuos en la segunda. También puede ser confundida con D. sanguinalis, pero esta última posee espiguillas glabrescentes a inconspicuamente pilosas y de mayor tamaño. Tanto D. horizontalis como D. sanguinalis presentan la lema superior enervia, mientras que en D. nuda ésta es 3-nervia. Véase la clave de las especies de Digitaria sect. Digitaria.
Ejemplar examinado
COLOMBIA. Valle del Cauca: Océano Pacífico, orilla norte de La Barra, entre Juan Chaco y Bocas del San Juan, en arenales de la playa, 1 m, 25 may 1975, R. Echeverry 2551 (COL).
7. Digitaria sanguinalis (L.) Scop., Fl. Carniol. (ed. 2) 1: 52. 1771. Panicum sanguinale L., Sp. Pl. 1: 57. 1753. TIPO. Cultivado en Leiden, Van Royen s.n. (holotipo no localizado; isotipo L). Fig. 13.
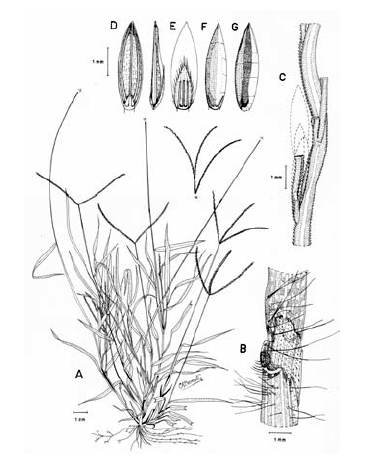
Figura 13. Digitaria sanguinalis (L.) Scop.: A. Hábito; B. Detalle de la región ligular; C. Porción de un racimo; D. Espiguilla, vista desde la lema inferior (vistas frontal y lateral); E. Espiguilla, vista desde la gluma superior; F. Antecio superior, visto desde la lema superior; G. Antecio superior, visto desde la pálea superior. Todo de Archer 3315 (COL).
Plantas anuales, cespitosas, follaje laxo a denso, caulinar. Cañas simples o ramificadas, erectas a decumbentes en las porciones basales, entonces originando raíces en los nudos inferiores y cañas ascendentes en éstos, cañas hasta de 50 cm de alto; entrenudos 3-9, pajizos a castaño claros, cilíndricos a canaliculados, hasta de 10 cm de longitud; nudos glabros, castaño oscuros. Vainas hasta de 10 cm de longitud, aunque frecuentemente de menor longitud, glabras, glabrescentes a finamente pubescentes, entonces los pelos hialinos de base tuberculada. Lígula membranácea a escariosa, glabra, rectangular a triangular, con el extremo distal dentado o eroso, 1-3 mm de longitud. Láminas hasta de 14 cm de longitud x 2-6 mm de ancho, planas, lanceoladas, ápice acuminado, glabras o pilosas hacia su porción proximal, los pelos de base tuberculada. Inflorescencias erectas, exertas a largamente exertas; pedúnculos hasta de 24 cm de longitud; eje principal de 0-2.5 cm de longitud; panojas constituidas por (2-) 3-12 racimos erectos, ascendentes, éstos hasta de 12 cm de longitud, conjugados, subdigitados, digitados a alternos sobre el eje principal o dispuestos en 1-2 verticilos; raquis triquetro, escabriúsculo marginalmente, 0.6-0.8 mm de ancho; espiguillas binadas, con pedicelos de diferente longitud, éstos trígonos, escabriúsculos, los más largos hasta de 2.6 mm de longitud, los más cortos hasta de 1 mm de longitud; articulación pedicelo-espiguilla truncada. Espiguillas 2.9-3.3 mm de longitud, lanceoladas, glabrescentes o muy cortamente pilosas, los pelos simples y suaves; gluma inferior membranácea, glabra, enervia, triangular, 0.3-0.5 mm de longitud; gluma superior generalmente ½ de la longitud de la espiguilla o hasta 2 mm de longitud, triangular, 3-nervia, cortamente pilosa; lema inferior de 2.9-3.3 mm de longitud, lanceolada, 5-7-nervia, los nervios escabrosos principalmente en su porción distal, glabrescente a cortamente pilosa; pálea inferior no vista; antecio superior tan largo como la lema inferior o ca. 0.2 mm más corto que ésta, lanceolado, estriado, cartilagíneo, glabro, pajizo, gris oscuro o azuloso, la lema superior con las márgenes membranáceas, hialinas a blanquecinas, plegadas sobre la pálea superior y cubriéndola casi completamente o 4/5 de ésta; lodículas ca. 0.3 mm de longitud, cordiformes. Cariopsis ca. 1.7 mm de longitud, comprimida dorsiventralmente; escudete embrional ca. ½ de la longitud de la cariopsis; hilo aovado.
Usos. Esta especie es apetecida por el ganado, especialmente el caballar. No obstante, D. sanguinalis crea problemas de invasión en cultivos de tierras bajas y medias por su lujurioso crecimiento (Luces de Febres 1963, Nicora & Rúgolo de Agrasar 1987, Rúgolo de Agrasar 1994).
Nombre vulgar. Los seis (Nariño).
Distribución geográfica y ecológica. Digitaria sanguinalis es nativa del Viejo Mundo y ha sido introducida en regiones cálidas y templadas de ambos hemisferios (Pohl & Davidse 1994). Ésta crece principalmente en áreas húmedas y alteradas y en campos de cultivo. En Colombia sólo ha sido registrada en los departamentos de Caquetá, Cauca, Cundinamarca, Magdalena, Nariño, Santander y Valle del Cauca, entre el nivel del mar y los 2000 m.
Observaciones. Digitaria sanguinalis es frecuentemente confundida con D. ciliaris. No obstante, D. sanguinalis presenta los nervios escabrosos en la porción distal de la lema inferior, característica que no está presente en D. ciliaris. Por otra parte, la gluma superior en D. sanguinalis es más pequeña que la de D. ciliaris. Véanse las observaciones dadas para D. nuda.
Ejemplares examinados
COLOMBIA. Caquetá: Morelia, 150-300 m, 2 oct 1941, K. Sneidern 1056 (COL, dos ejemplares). Cauca: Popayán, 5 may 1935, H. García-Barriga 4380 (COL). Cundinamarca: La Florida, Anolaima, abr 1935, Archer 3315 (COL). Magdalena: Sierra Nevada de Santa Marta, southeastern slopes, Hoya del Río Donachuí, below the village Donachuí near the river, 1230-1350 m, 24 sep 1959, J. Cuatrecasas & R. Romero 24396 (COL). Nariño: Mun. Imués, Pilcuán, 1910 m, 16 abr 1966, G. López & M. de la Rosa 57-F (COL). San José de Albán, 1800 m, oct 1966, C. Martínez & H. Martínez 45 (COL). Santander: Zapatoca, ca. 1800 m, 22 jul 1944, N. Fassett 25497 (COL).
8. Digitaria setigera Roth, Syst. Veg. 2: 474. 1817. TIPO. India, Heyne s.n. (holotipo B; isotipo K, no vistos; fotografía del holotipo publicada en Bor, Webbia 11: Tav. 2. 1956). Fig. 14.
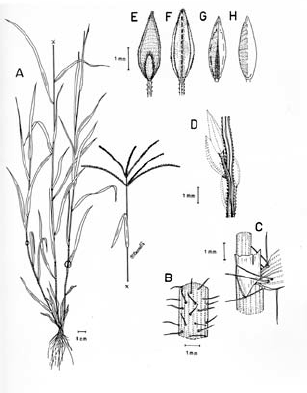
Figura 14. Digitaria setigera Roth: A. Hábito; B. Detalle del indumento de las vainas; C. Detalle de la región ligular; D. Porción de un racimo; E. Espiguilla, vista desde la gluma superior; F. Espiguilla, vista desde la lema inferior; G. Antecio superior, visto desde la pálea superior; H. Antecio superior, visto desde la lema superior. Todo de Fonnegra et al. 2476 (COL).
Plantas anuales, cespitosas, follaje laxo a denso, caulinar. Cañas simples o ramificadas, erectas a decumbentes en las porciones basales, entonces originando raíces en los nudos inferiores y cañas ascendentes en éstos, cañas hasta de 100 cm de alto; entrenudos 3-10, pajizos a castaño claros, cilíndricos a canaliculados, hasta de 10 cm de longitud; nudos glabros, castaño oscuros. Vainas hasta de 10 cm de longitud, aunque frecuentemente de menor longitud, glabras, glabrescentes o pilosas, entonces los pelos hialinos de base tuberculada. Lígula membranácea a escariosa, glabra, truncada a erosa, 1.8-3.4 mm de longitud. Láminas hasta de 15 cm de longitud x 3-8 (-10) mm de ancho, planas, lanceoladas, ápice acuminado, glabras o pilosas principalmente hacia su porción proximal, los pelos de base tuberculada. Inflorescencias erectas, exertas a largamente exertas; pedúnculos hasta de 40 cm de longitud; eje principal de 0-2.5 cm de longitud; panojas constituidas por 3-6 (-8) racimos erectos, ascendentes, éstos hasta de 12 (-15) cm de longitud, subdigitados, digitados a alternos sobre el eje principal o dispuestos en 1-3 verticilos; raquis triquetro, escabriúsculo marginalmente, 0.5-0.6 (-0.8) mm de ancho; espiguillas binadas, con pedicelos de diferente longitud, éstos trígonos, escabriúsculos, los más largos hasta de 2.6 mm de longitud, los más cortos hasta de 0.7 mm de longitud; articulación pedicelo-espiguilla truncada, raramente discoide. Espiguillas (2.7-) 2.8-3.3 mm de longitud, lanceoladas, glabrescentes a pilosas, los pelos simples y suaves; gluma inferior ausente; gluma superior 0.5-1 mm de longitud, truncada a triangular o de ápice obtuso, 0-3-nervia, glabrescente a muy cortamente pilosa, los pelos simples y suaves; lema inferior de (2.7-) 2.8-3.3 mm de longitud, lanceolada, 5-7-nervia, glabrescente a pubescente, los pelos suaves y muy cortos; pálea inferior ca. 0.2 mm de longitud, membranácea, hialina a pajiza; antecio superior tan largo como la lema inferior o ca. 0.2 mm más corto que ésta, lanceolado, estriado, cartilagíneo, glabro, pajizo a castaño muy claro o grisáceo, la lema superior con las márgenes membranáceas, hialinas a blanquecinas, plegadas sobre la pálea superior y cubriendo ca. 2/3-4/5 de ésta; lodículas ca. 0.3 mm de longitud, cordiformes. Cariopsis ca. 1.7 mm de longitud, comprimida dorsiventralmente; escudete embrional ca. ½ de la longitud de la cariopsis; hilo aovado.
Distribución geográfica y ecológica. Digitaria setigera es una especie nativa de Asia tropical y ha sido recientemente introducida en el trópico americano, siendo una maleza característica de bordes de caminos (Veldkamp 1973, Pohl 1980, Webster 1987, Webster & Hatch 1981, Pohl & Davidse 1994). Esta especie crece entre el nivel del mar y los 1500 m.
Observaciones. Digitaria setigera es fácilmente reconocible, pues carece de gluma inferior y además, la gluma superior es muy corta con relación a la longitud de la espiguilla. Estas características permiten separarla fácilmente de las demás especies de Digitaria sect. Digitaria. Véanse las observaciones dadas para D. ciliaris.
Ejemplares examinados
COLOMBIA. Antioquia: Mun. Dabeiba, 27 km después por la carretera al mar, 250 m, 28 sep 1986, J. Betancur et al. 257 (COL, HUA, MO). Mun. Chigorodó, hacienda Montecristo, 25 km después de Apartadó, en cultivo de “chontaduro”, 40 m, 2 oct 1986, J. Betancur et al. 347 (COL, HUA, MO), 3 oct 1986, J. Betancur et al. 355 (HUA: 35388, MO). Mun. Mutatá, carretera a Pavarandogrande, 2 km después de Mutatá, 150 m, 3 oct 1986, J. Betancur et al. 359 (HUA, MO). Mun. Dabeiba, 32 km SE de Mutatá, en la vía Mutatá-Dabeiba, río Chever, sitio Chever, 280 m, 5 ago 1987, R. Callejas et al. 5077 (COL, HUA, MO). Mun. Mutatá, carretera Mutatá-Pavarandó, potreros de la hacienda El Darién, 150 m, 29 abr 1987, R. Fonnegra et al. 1941 (HUA, MO), 4 may 1987, R. Fonnegra et al. 2152 (HUA, MO). Mun. Andes, 12.5 km de Andes hacia el corregimiento de Tapartó, vereda California, apiario California, 24 ago 1988, R. Fonnegra et al. 2476 (HUA, MO, SI). Quindío: vía Armenia-Barcelona-Caicedonia, 1250 m, 3 may 2000, C. Agudelo et al. 4159 (COL). San Andrés, Providencia y Santa Catalina: west side between airport and The Cove, 0-5 m, 29 ene 1985, J. Wood 4723 (COL).
Digitaria sect. Ischaemum Ohwi
9. Digitaria fuscescens (J. Presl) Henrard, Meded. Rijks-Herb. 61: 8. 1930. Paspalum fuscescens J. Presl, Reliq. Haenk. 1 (4-5): 213. 1830. TIPO. “Hab. in regione montana Peruviae”, Haenke s.n. (holotipo PR, no visto; isotipos MO!, L, US, W, no vistos). Fig. 15.
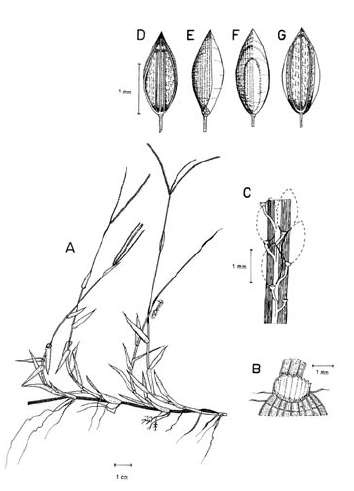
Figura 15. Digitaria fuscescens (J. Presl) Henrard: A. Hábito; B. Detalle de la región ligular; C. Porción de un racimo; D. Espiguilla, vista desde la lema inferior; E. Antecio superior, visto desde la pálea superior; F. Antecio superior, visto desde la lema superior; G. Espiguilla, vista desde la gluma superior. Todo de Davidse 5126 (COL).
Plantas anuales (?), gráciles, estoloníferas, los estolones hasta de 40 cm de longitud, originándose en éstos numerosas cañas floríferas; follaje laxo. Cañas floríferas erectas, simples, hasta de 23 cm de altura; entrenudos 3-5, pajizos, vináceos a verde claros, cilíndricos a colapsados, estriados longitudinalmente; nudos glabros, castaño oscuros. Vainas hasta de 6 cm de longitud, siendo más largas las vainas distales, glabras. Lígula membranácea a escariosa, glabra, redondeada con la porción distal levemente irregular, 1.2-1.6 mm de longitud. Láminas hasta de 3.5 cm de longitud x 2-4 mm de ancho, lanceoladas, planas, glabras, en ocasiones escasamente pilosas en la región ligular, entonces los pelos en un número ingerior a 8, éstos hialinos, de base tuberculada, hasta de 1.5 mm de longitud, ápice acuminado. Inflorescencias terminales, erectas, exertas; pedúnculos capilares, hasta de 13 cm de longitud; eje principal 0-0.3 cm de longitud; panojas constituidas por 2-3 racimos, ligeramente distanciados entre si a más comúnmente digitados, erectos a curvos, ascendentes, hasta de 6 cm de longitud; raquis alado, glabro, escabriúsculo marginalmente, ca. 0.7 mm de ancho; espiguillas dispuestas en tríades o excepcionalmente binadas, con pedicelos de diferente longitud, éstos subcilíndricos, capilares, escabriúsculos, los más largos hasta de 1.2 mm de longitud, los más cortos hasta de 0.4 mm de longitud; articulación pedicelo-espiguilla cupuliforme. Espiguillas 1.3-1.4 (-1.5) mm de longitud x ca. 0.6 mm de ancho, elípticas, agudas, glabras a glabrescentes, los pelos verrugosos (vistos a gran aumento); gluma inferior ausente o reducida a una minúscula membrana, hialina, glabra, enervia, ca. 0.1 mm de longitud; gluma superior de igual longitud que la lema inferior, 3-nervia; lema inferior de 1.3-1.4 (-1.5) mm de longitud, 7-nervia, los nervios en ocasiones anastomosados hacia su porción distal; pálea inferior ca. 0.1 mm de longitud; antecio superior tan largo como la lema inferior o ca. 0.15 mm más largo que ésta, elíptico, agudo a acuminado o ligeramente apiculado, estriado, cartilágíneo, glabro, pajizo, raramente con la porción distal vinácea, brillante, las márgenes de la lema superior membranáceas, hialinas, plegadas sobre la pálea superior y cubriéndola casi completamente; lodículas ca. 0.2 mm de longitud, escariosas, hialinas a pajizas, truncadas, con el borde superior ligeramente irregular. Cariopsis no vista.
Distribución geográfica y ecológica. Digitaria fuscescens es una especie nativa de África tropical y ha sido introducida en Mesoamérica y Sudamérica tropical (Pohl & Davidse 1994). Esta especie crece en sabanas, junto con especies del género Trachypogon (Poaceae: Andropogoneae). De Colombia sólo se poseen registros de Amazonas, Meta y Vichada, entre los 100 y 400 m.
Observaciones. En vista de su raquis alado, de la semejanza de sus inflorescencias y del tamaño de sus espiguillas, D. fuscescens podría confundirse con D. longiflora y D. violascens. No obstante, D. fuscescens posee espiguillas glabras a glabrescentes, mientras que en las otras dos especies las espiguillas son pilosas. Por otra parte, el antecio superior de D. fuscescens y de D. longiflora es pajizo, mientras que el de D. violascens es castaño oscuro a negruzco, lo que combinado con el indumento de las espiguillas, ayuda eficazmente a separar estas especies.
Ejemplares examinados
COLOMBIA. Amazonas: río Caquetá, sabanas de arenisca en contornos del aeropuerto, en pista de aterrizaje, 200-250 m, 18 jul 1977, A. Fernández-Pérez 20063 (COL). Meta: Mun. Puerto Gaitán, C.I. Carimagua, “Ensayo Culticore”, 150 m, 22 may 1996, R. Serna Isaza 738 (COAH, tres ejemplares). Vichada: ca. 8 km E of Las Gaviotas, along the unimproved dirt road to Santa Rita, completely open Trachypogon savanna with forest along spring fed stream, 220 m, 25 dic 1973, G. Davidse & F. Llanos 5126 (COL).
10. Digitaria longiflora (Retz.) Pers., Syn. Pl. 85. 1805. Paspalum longiflorum Retz., Observ. Bot. 4: 15. 1786. TIPO. India, König s.n. (holotipo LD; isotipos BM, K, no vistos). Fig. 16.
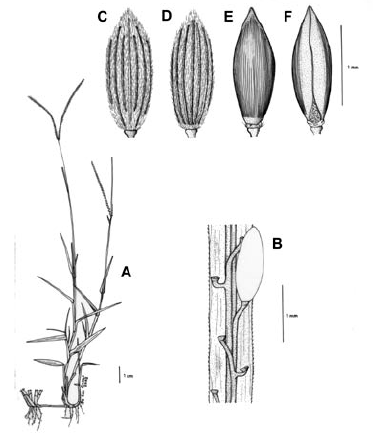
Figura 16. Digitaria longiflora (Retz.) Pers.: A. Hábito; B. Porción de un racimo; C. Espiguilla, vista desde la gluma superior; D. Espiguilla, vista desde la lema inferior; E. Antecio superior, visto desde la lema superior; F. Antecio superior, visto desde la pálea superior. Todo de Wood 4702 (COL).
Plantas anuales, gráciles, densamente cespitosas; follaje denso. Cañas erguidas a decumbentes, ramificadas, hasta de 20 cm de altura; entrenudos 4-7, pajizos, cilíndricos a colapsados, ligeramente estriados longitudinalmente, capilares, hasta de 3 cm de longitud, siendo los entrenudos basales más cortos que los distales; nudos glabros, castaño claros a oscuros. Vainas glabras, hasta de 4 cm de longitud, aunque normalmente no sobrepasan los 2.5 cm de longitud, siendo las vainas distales más largas. Lígula membranácea a escariosa, glabra, con la porción distal entera a ligeramente irregular, ca. 1 mm de longitud. Láminas hasta de 2.5 cm de longitud x 1.5-4 mm de ancho, glabras, planas, lineares a más comúnmente lanceoladas, ápice acuminado. Inflorescencias terminales, erectas, exertas; pedúnculos capilares, hasta de 10 cm de longitud; eje principal nulo, pues los racimos son conjugados; panojas constituidas por 2 racimos conjugados, erectos a curvos, hasta de 4.5 cm de longitud; raquis alado, glabro, marginalmente escabriúsculo, ca. 0.75 mm de ancho; espiguillas dispuestas en tríades, con pedicelos de diferente longitud, éstos capilares, flexuosos, lisos, glabros, los más largos hasta de 1.8 mm, los más cortos hasta de 0.4 mm de longitud; articulación pedicelo-espiguilla cupuliforme. Espiguillas ca.1.5 mm de longitud x ca. 0.6 mm de ancho, elípticas a oblongo-elípticas, agudas, mediana y cortamente pilosas, raras veces densamente pilosas, los pelos verrugosos, agudos, blanquecinos, adpresos a levemente divergentes, ca. 0.2 mm de longitud; gluma inferior ausente; gluma superior de igual longitud que la lema inferior, 3-nervia; lema inferior ca. 1.5 mm de longitud, 7-nervia; pálea inferior ca. 0.2 mm de longitud, carnosas; antecio superior tan largo como la lema inferior, elíptico a oblongo-elíptico, agudo a acuminado, estriado, cartilagíneo, glabro, pajizo, brillante, las márgenes de la lema superior membranáceas, hialinas, plegadas sobre la pálea superior y cubriendo cerca de 1/3 de ésta; lodículas ca. 0.25 mm de longitud. Cariopsis no vista.
Distribución geográfica y ecológica. Digitaria longiflora es una especie nativa del Viejo Mundo y ha sido introducida en México, Costa Rica, Perú, Brasil (Pohl 1980, Pohl & Davidse 1994) y Guayanas (Judziewicz 1990) y crece en sabanas húmedas y de suelos arenosos. De Colombia sólo se conocen dos colecciones de áreas bajas del departamento del Cesar.
Observaciones. Véanse las observaciones dadas para D. fuscescens.
Ejemplares examinados
COLOMBIA. Cesar: Hacienda Curucucu, near Chiriguaná, 150 m, 31 dic 1984, J. Wood 4702 (COL). Playones de Chiriguaná, in flooded land on sandy-silty soil, 50 m, 16 jul 1985, J. Wood 4988 (COL).
11. Digitaria violascens Link, Hort. Berol. 1: 229. 1827. TIPO. Cultivado en el Hort. Bot. Berolin. a partir de material recolectado en Brasil, Herb. Link 93 (holotipo B; isotipo BAA, no vistos). Fig. 17.
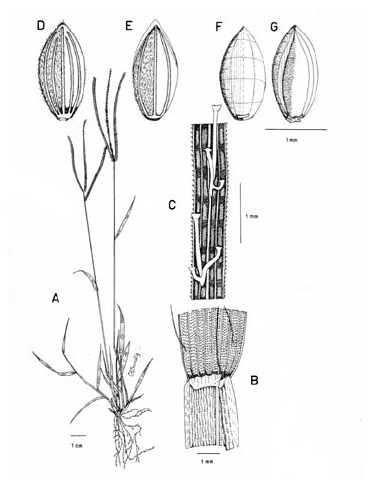
Figura 17. Digitaria violascens Link: A. Hábito; B. Región ligular; C. Porción de un racimo; D. Espiguilla desde el lado de la lema inferior; E. Espiguilla desde el lado de la gluma superior; F. Antecio superior desde el lado de la lema superior; G. Antecio superior desde el lado de la pálea superior. Todo de Giraldo-Cañas 3433 (COL).
Plantas anuales, gráciles, cespitosas; follaje laxo a medianamente denso, principalmente basal. Cañas erguidas, simples a más comúnmente ramificadas desde la base, hasta de 70 cm de altura, aunque normalmente la altura no supera los 30 cm; entrenudos 3-6, pajizos, cilíndricos a colapsados, canaliculados a más frecuentemente estriados longitudinalmente, capilares, hasta de 7 cm de longitud, siendo los entrenudos inferiores más cortos que los distales; nudos glabros, castaño oscuros a verde oscuros o violáceos. Vainas hasta de 8 cm de longitud, glabras. Lígula membranácea a escariosa, glabra, con la porción distal lisa o irregular a brevemente laciniada, 0.6-1.5 mm de longitud. Láminas hasta de 20 cm de longitud x 1-4 mm de ancho, aunque normalmente su longitud no sobrepasa los 7 cm, glabras, planas, lanceoladas, ápice acuminado. Inflorescencias terminales, erectas, exertas a largamente exertas; pedúnculos capilares, hasta de 29 cm de longitud; eje principal de 0-1.5 cm de longitud; panojas constituidas por 2-7 racimos, erectos, ascendentes, hasta de 13 cm de longitud, aunque normalmente no sobrepasan los 11 cm, digitados, verticilados a cortamente alternos o menos comúnmente distribuidos en dos verticilos; raquis alado, glabrescente, escabriúsculo marginalmente, 0.6-0.8 mm de ancho; espiguillas dispuestas en tríades o en tétradas, con pedicelos de diferente longitud, éstos capilares, escabriúsculos, flexuosos, los más largos hasta de 1.5 mm de longitud, los más cortos hasta de 0.5 mm de longitud; articulación pedicelo-espiguilla cupuliforme. Espiguillas 1.5-1.6 mm de longitud x ca. 0.7 mm de ancho, ovadas, escasa y cortamente pilosas, los pelos agudos y verrugosos, ca. 0.15 mm de longitud, adpresos a levemente divergentes, hialinos a blanquecinos; gluma inferior ausente a más comúnmente presente, aunque reducida a una minúscula membrana, hialina, glabra, enervia, ca. 0.15 mm de longitud; gluma superior igual que la lema inferior o apenas ca. 0.1 (-0.2) mm más corta que ésta, de ápice agudo a acuminado, 3-nervia, los nervios en ocasiones anastomosados hacia su porción distal; lema inferior de 1.5-1.6 mm de longitud, de ápice agudo a acuminado, (5-) 7-nervia, los nervios en ocasiones anastomosados hacia su porción distal; pálea inferior ca. 0.2 mm de longitud; antecio superior tan largo como la lema inferior o ligeramente más largo que ésta, ovado a elipsoide, agudo a acuminado, estriado, cartilagíneo, glabro, castaño oscuro a negruzco, brillante, las márgenes de la lema superior membranáceas, blanquecinas a hialinas, plegadas sobre la pálea superior y cubriéndola casi completamente; lodículas ca. 0.25 mm de longitud, escariosas, hialinas, truncadas. Cariopsis ca. 1 mm de longitud, pajiza a hialina; escudete embrional ca. 0.3 mm de longitud; hilo aovado.
Distribución geográfica y ecológica. Digitaria violascens se distribuye en las regiones tropicales y subtropicales de Asia, Australia y América (Rúgolo de Agrasar 1994). En Colombia sólo se conoce de los departamentos de Antioquia, Boyacá, Cauca, Cundinamarca, Huila, Meta, Quindío y Tolima, y crece en áreas degradadas, bordes de camino y zonas de cultivo de la región andina y en áreas de arenas blancas del piedemonte llanero en el departamento del Meta, entre los 400 y 2200 m.
Observaciones. En vista de la naturaleza de los pelos de las espiguillas, los cuales son verrugosos, y de su raquis alado, D. violascens debe ser ubicada en Digitaria sect. Ischaemum, tal como había sido considerado por Veldkamp (1973) y Webster (1983), a diferencia de la ubicación dada por Rúgolo de Agrasar (1974), quien la ubicó en Digitaria sect. Ternatae. Sin embargo, el color de su antecio superior (castaño oscuro a negruzco) la acerca a los miembros de Digitaria sect. Ternatae.
Los ejemplares de la población de Piedras Gordas (departamento de Antioquia, Giraldo-Cañas et al. 3299) presentan dimensiones más grandes, correspondiendo a los extremos registrados para la especie en Mesoamérica (véase Pohl & Davidse 1994). Véanse las observaciones dadas para D. fuscescens y D. filiformis.
Ejemplares examinados
COLOMBIA. Antioquia: Mun. Carmen de Viboral, campus universitario, Universidad de Antioquia, sede Oriente; creciendo con Axonopus compressus, Eragrostis tenuifolia, Paspalum sp. y Sporobolus indicus, ca. 2200 m, 26 dic 2003, D. Giraldo-Cañas et al. 3617 (COL). Mun. Jardín, sitio La Truchería, carretera Jardín-Riosucio, km 5, 1900-2000 m, 5 ene 2003, D. Giraldo-Cañas et al. 3433 (COL). Mun. Santo Domingo, vereda Piedras Gordas, quebrada Piedras Gordas, carretera Barbosa-Cisneros, a la altura de Pescadito Nro. 1, en inmediaciones de la finca de Don Manuel Arango-Rozzo, 1400-1500 m, 4 ene 2002, D. Giraldo-Cañas et al. 3299 (COL, dos ejemplares). Mun. Rionegro, vereda La Mosca, finca de Margarita Zuleta, en huerto de Citrus, 2150 m, 30 ago 1986, R. Pohl 15451 (COL, HUA, MO). Mun. Urrao, carretera a La Encarnación, en lodos al lado de la carretera, 1650 m, 10 sep 1986, R. Pohl & J. Betancur 15480 (HUA, dos ejemplares). Mun. Andes, aeropuerto, en área abierta cerca de la pista, 1200 m, 17 sep 1986, R. Pohl & J. Betancur 15519 (HUA, dos ejemplares, MO), 15526 (HUA, dos ejemplares). Mun. Jardín, cerro de Morro Amarillo, 7 km de Jardín, en cultivo de “yuca”, 2000 m, 20 sep 1986, R. Pohl & M. Palacio 15577 (COL, HUA, MO). Boyacá: Mun. Santa María, sendero ecológico de Hyca Quye, carretera entre Santa María y Juntas, 1075 m, 27 mar 2001, J. Betancur et al. 9137 (COL). Cauca: Mun. Popayán, predios internos y jardineras de la Facultad de Educación de la Universidad del Cauca, 1750 m, 7-11 nov 2004, D. Giraldo-Cañas et al. 3815 (COL). Mun. Popayán, jardines externos del Banco de la República, 1750 m, 7-11 nov 2004, D. Giraldo-Cañas et al. 3817 (COL). Mun. Popayán, río Palacé, vía a Totoró km 2.5, alrededores del puente, 1850 m, 1 dic 2000, B. Ramírez 13737-B (COL). Cundinamarca: Ubalá B, Inspección de Policía Mámbita, campamento EMGESA, 880 m, 26 jun 1998, J. Fernández et al. 15920-B (COL). Provincia de Río Negro, Mun. Pacho, carretera Pacho-La Capilla, alrededores del estadio municipal de fútbol, ca. 1600 m, 13 ene 2004, D. Giraldo-Cañas et al. 3728 (COL). Huila: a few km SW of Gallego, on a disturbed open roadside on stony soil, 1400 m, 23 oct 1983, J. Wood 4052 (COL). Meta: Mun. Villavicencio, carretera Villavicencio-Aeropuerto, piedemonte de la cordillera Oriental, sitio La Arenera, ca. 2 km del puente sobre el río Guatiquía, en cantera abandonada, área rocosa y de arenas blancas, dominada por gramíneas, cyperáceas, xyridáceas y eriocauláceas, ca. 400 m, 10 nov 2002, D. Giraldo-Cañas et al. 3350 (COAH, COL, HUA). Quindío: Mun. Armenia, jardineras de la Avenida del Museo Quimbaya, 1500 m, 20-25 mar 2005, D. Giraldo-Cañas & J. García-Ulloa 3872 (COL). Mun. Salento, camino principal hacia El Mirador, 1500 m, 20-25 mar 2005, D. Giraldo-Cañas & J. García-Ulloa 3895 (COL). Tolima: Mun. Ibagué, jardineras de la Plaza de Bolívar, ca. 1200 m, 11-12 jun 2005, D. Giraldo-Cañas et al. 3907, 3911 (COL).
Digitaria sect. Ternatae Hack.
12. Digitaria andicola Giraldo-Cañas, Caldasia 25 (2): 211. 2003. TIPO. Colombia. Cundinamarca: Mun. Nemocón, carretera principal de la vereda Susatá que comunica hacia la cabecera municipal de Nemocón, inmediaciones de la hacienda Susatá, en matorrales degradados de Dodonaea viscosa, Opuntia sp. y Solanum sp., 2700 m, 4 nov 2002, D. Giraldo-Cañas et al. 3319 (holotipo COL!; isotipos COL!). Fig. 18.
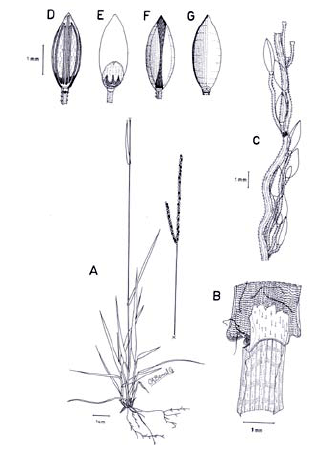
Figura 18. Digitaria andicola Giraldo-Cañas: A. Hábito; B. Región ligular; C. Porción de un racimo; D. Espiguilla desde el lado de la lema inferior; E. Espiguilla desde el lado de la gluma superior; F. Antecio superior desde el lado de la pálea superior; G. Antecio superior desde el lado de la lema superior. Todo del holotipo.
Plantas perennes, cespitosas; follaje laxo a medianamente denso, principalmente basal. Cañas erguidas a levemente decumbentes, simples, muy ocasionalmente ramificadas en los nudos medios, hasta de 35 cm de alto; entrenudos regularmente 3, raramente hasta 5, pajizos, estriados longitudinalmente, cilíndricos a levemente colapsados, hasta de 8 cm de longitud, siendo más cortos los entrenudos inferiores; nudos glabros, castaño-oscuros. Vainas hasta de 9 cm de longitud, aunque normalmente de menos de 5 cm, glabras a glabrescentes, sólo corta y laxamente pilosas hacia su porción distal, los pelos unicelulares, hialinos, rectos a levemente flexuosos, perpendiculares a la vaina, de base tuberculada, hasta de 2 mm de longitud. Lígula membranácea a escariosa, erosa, glabra o cortamente ciliada a pestañosa, 0.5-1.3 (--2) mm de longitud. Láminas hasta de 17 cm de longitud, aunque normalmente no superan los 10 cm, 1.5-4 (-5) mm de ancho, glabras o escasamente pilosas (principalmente hacia la región ligular), los pelos de la misma naturaleza de los de las vainas, aunque más largos, hasta 4 mm de longitud, planas, lineares, ápice acuminado. Inflorescencias terminales, erectas, largamente exertas; pedúnculos capilares, hasta de 26 cm de longitud; eje principal de 0-4.5 cm de longitud; panojas constituidas por 1-4 racimos erectos, ascendentes, hasta de 11 cm de longitud, próximos entre si; raquis triquetro, glabrescente, escabriúsculo marginalmente, 0.4-0.6 mm de ancho; espiguillas dispuestas en tríades o en tétradas, con pedicelos de diferente longitud, éstos filiformes a capilares, subcilíndricos a planos, flexuosos, glabrescentes a corta y laxamente pilosos, escabriúsculos, los más largos hasta de 4 mm de longitud, los más cortos hasta de 1 mm de longitud. Espiguillas 2-2.5 mm de longitud, ca. 0.8 mm de ancho, ovadas a elipsoides; gluma inferior ausente o reducida a una minúscula membrana, glabra, escariosa, entera o erosa, enervia, de menos de 0.2 mm de longitud; gluma superior de 0.8-1.5 (-1.8) mm de longitud (0.5-1.4 mm más corta que el antecio superior), escariosa, de ápice redondeado, emarginado o eroso, 3-nervia, los nervios gruesos, glabrescente a escasamente pilosa, los pelos conspicuamente estipitados, claviformes, hialinos, adpresos a levemente divergentes, ca. 0.2 mm de longitud; lema inferior de 2-2.5 mm de longitud, membranácea, acuminada, (5--) 7-nervia, los nervios gruesos, glabrescente a ligeramente pilosa, los pelos como los de la gluma superior aunque ocasionalmente un poco más abundantes; pálea inferior 0.3-0.5 mm de longitud, de márgenes irregulares, carnosa, verruculosa, pajiza a verde muy clara; antecio superior tan largo como la lema inferior, elipsoide, acuminado a levemente apiculado, estriado, cartilagíneo, glabro, castaño oscuro, brillante, las márgenes de la lema superior membranáceas, blanquecinas a hialinas, plegadas sobre la pálea superior y cubriéndola casi completamente; lodículas ca. 0.25 mm de longitud, hialinas a pajizas, truncadas, de ápice levemente irregular. Cariopsis no vista.
Distribución geográfica y ecológica. Hasta el momento, esta especie sólo se conoce de algunas áreas andinas de Colombia (departamentos de Boyacá, Cundinamarca, Nariño y Valle del Cauca). Digitaria andicola es poco frecuente y crece en hábitats abiertos de áreas rocosas y en matorrales de áreas secas de regiones montañosas entre los 2000 y 2800 m de altitud. Estos ambientes están dominados por Dodonaea viscosa (Sapindaceae), Opuntia spp. (Cactaceae) y Solanum sp. (Solanaceae), y en menor medida por Salvia spp. (Lamiaceae), entre el material arbustivo, mientras que el componente herbáceo está caracterizado por varias gramíneas (Agrostis, Andropogon, Axonopus, Bothriochloa, Chloris, Eragrostis, Heteropogon, Lycurus, Melinis, Microchloa, Muhlenbergia, Nassella, Paspalum, Piptochaetium, Schizachyrium, Setaria, Sporobolus, Stipa y Vulpia) y diversas especies de Cuphea (Lythraceae), Echeveria (Crassulaceae), Epidendrum (Orchidaceae), Evolvulus (Convolvulaceae), Plantago (Plantaginaceae) y Polygala (Polygalaceae) (observaciones personales). También crece en los sotobosques abiertos de plantaciones de Acacia decurrens (Mimosaceae) y Eucalyptus globulus (Myrtaceae), establecidas en áreas secas entre los 2400 y 2700 m de la cordillera Oriental (observaciones personales). Esta especie es la que presenta el mayor rango altitudinal entre las especies colombianas de Digitaria, llegando hasta los 2800 m.
Observaciones. En vista de su hábito, de las dimensiones de su gluma superior, de la forma de la espiguilla, de la forma, el ornamento y el color del antecio superior, D. andicola es afín y similar a D. curtigluma Hitchc. [ejemplar examinado: México, M. González 600 (MO)] y D. curvinervis (Hack.) Fernald [ejemplar examinado: Panamá, J. McCorkle C-264 (MO)]. No obstante, éstas pueden diferenciarse fácilmente por la siguiente clave:
1. Espiguillas de 1.4-1.5 mm de longitud; gluma superior aguda a obtusa, 0.5-0.7 mm más corta que el antecio superior; lema inferior 0.1-0.3 mm más corta que el antecio superior; lema superior cubriendo casi completamente o ca. 4/5 partes de la pálea superior; pálea inferior de 0.2 mm de longitud; plantas anuales.
Digitaria curvinervis
1’. Espiguillas de 2-2.5 mm de longitud; gluma superior de ápice redondeado o emarginado a eroso; lema inferior tan larga como el antecio superior; pálea inferior de 0.3-0.5 mm de longitud; plantas perennes.
2. Gluma superior 0.5-1.4 mm más corta que el antecio superior, nervios conspicuos; las márgenes de la lema superior cubriendo casi completamente la pálea superior; espiguillas de 2-2.5 mm de longitud, glabrescentes a escasamente pilosas. Digitaria andicola
2’. Gluma superior 1.9-2.1 mm más corta que el antecio superior, nervios tenues; las márgenes de la lema superior cubriendo 1/2-1/3 parte de la pálea superior; espiguillas de 2.3-2.5 mm de longitud, conspicuamente pilosas. Digitaria curtigluma
Por otra parte, D. andicola junto con D. cardenasiana son las especies de Digitaria sect. Ternatae presentes en Colombia que poseen las espiguillas más grandes (D. andicola: 2-2.5 mm de longitud, D. cardenasiana: 1.8-2.4 mm), mientras que en las restantes dos especies de la sección, la longitud de sus espiguillas oscila entre 1.2 y 1.7 mm. Estas dimensiones ayudan a separar las dos primeras especies de las restantes presentes en Colombia. Ahora bien, D. andicola se puede separar fácil y principalmente de D. cardenasiana por la naturaleza de los pedúnculos (glabros en D. andicola vs. glabrescentes o pilosos a hirsutos en D. cardenasiana), por la densidad del indumento de las espiguillas (glabrescentes a ligeramente pilosas en D. andicola vs. densamente pilosas en D. cardenasiana), por la naturaleza del indumento de las espiguillas (pelos conspicuamente estipitados en D. andicola vs. cortamente estipitados en D. cardenasiana) y por la longitud de la gluma superior (0.5-1.4 mm más corta que la lema inferior en D. andicola vs. 0-0.25 mm más corta en D. cardenasiana).
La población de D. andicola de Chachagüí (2000 m, departamento de Nariño, sur de Colombia) presenta los extremos de variación en sus estructuras vegetativas (tamaño general de las plantas, dimensiones de las vainas, lígulas y láminas foliares), siendo éstas más grandes que las que presentan los individuos de las poblaciones del altiplano cundiboyacense (departamentos de Boyacá y Cundinamarca, en el centro-este de Colombia).
Ejemplares examinados
COLOMBIA. Boyacá: Mun. Villa de Leyva, camino del cañón de Las Clusias, a la cumbre de Morro Negro, en arbustales con presencia de Clusia sp., Dodonaea viscosa, Befaria spp., Hypericum spp., Asteraceae y Melastomataceae, estrato herbáceo con abundacia de Puya y Espeletiopsis, 2700 m, 9 jun 2001, O. Chols et al. 109 (COL). Mun. Villa de Leyva, camino entre la hacienda “Torcoroma de Arriba” y el cañón de Las Clusias; matorrales degradados de áreas secas y rocosas, dominados principalmente por Dodonaea viscosa, y en segunda medida por varias especies de gramíneas (Agrostis sp., Andropogon spp., Axonopus compressus, Bothriochloa sp., Eragrostis spp., Melinis minutiflora, Paspalum spp., Piptochaetium sp., Setaria sp., Sporobolus spp. y Stipa sp.), ca. 2500 m, 18 jul 2003, D. Giraldo-Cañas et al. 3536 (COL). Mun. Villa de Leyva, matorrales saliendo por el camino que conduce a Iguaque, en las afueras del casco urbano de Villa de Leyva, 2200 m, 18 dic 2004, D. Giraldo-Cañas et al. 3833 (COL). Mun. Iza, en inmediaciones de Los Termales, en cantera abandonada, ca. 2500 m, 22 jun 2003, D. Giraldo-Cañas et al. 3524 (COL). Mun. Sáchica, ca. 2300 m, ago 1964, C. Saravia 4299 (COL), 4365-B (COL); sin localidad, 2700 m, 15 abr 1964, C. Saravia 3949 (COL) ; sin localidad, 2700 m, 15 abr 1964, C. Saravia 3960-B (COL). Cundinamarca: Mun. Suesca-Nemocón, hacienda Susatá, en bosques de Acacia y en pajonales 2650 m, 23 ago 2000, J. Fernández et al. 19183 (COL), 19188 (COL), 11 nov 1999, J. Groenendijk & N. Rietman 1218 (COL), 2 ago 2000, J. Groenendijk 1537 (COL). Mun. Mosquera, vereda Santa Helena, S of Mosquera, in sparse stony grassland on exposed treelers eroded stony hills, 2700 m, año 1983, J. Wood 3726 (COL). Nariño: Mun. Pasto, corregimiento de Chachagüí, 2000 m, 1 jun 1989, B. Ramírez 1564 (COL, PSO). Valle del Cauca: Cordillera Central, vertiente occidental, hoya del río Bugalagrande, Loma de Barragán, desde La Parrilla a La Machuca, 2660-2750 m, 13-14 abr 1946, J. Cuatrecasas 20661 (VALLE).
13. Digitaria cardenasiana Giraldo-Cañas, Caldasia 24 (2): 369. 2002. TIPO. Colombia. Meta: Mun. Puerto López, finca Los Laureles, formando sociedades casi puras, 300 m, 3 jun 1980, O. Rangel & J. Espina 1418 (holotipo COL!). Fig. 19.
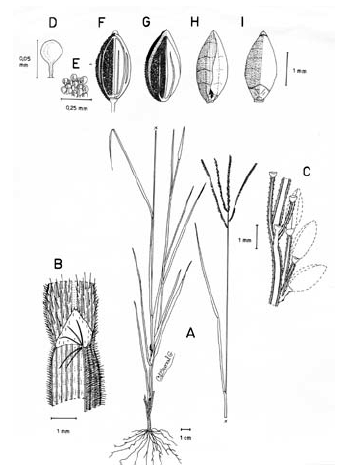
Figura 19. Digitaria cardenasiana Giraldo-Cañas: A. Hábito; B. Región ligular; C. Porción de un racimo; D y E. Detalle de la naturaleza de los pelos de la espiguilla; F. Espiguilla desde el lado de la lema inferior; G. Espiguilla desde el lado de la gluma superior; H. Antecio superior desde el lado de la pálea superior; I. Antecio superior desde el lado de la lema superior. Todo del holotipo.
Plantas perennes, cespitosas; follaje medianamente denso a denso, principalmente basal. Cañas erguidas, simples, hasta de 60 cm de altura; entrenudos 3-4, pajizos, levemente estriados a más comúnmente lisos, cilíndricos, hasta de 16 cm de longitud, siendo más cortos los entrenudos inferiores; nudos glabros, pajizos a castaño-claros. Vainas hasta de 12 cm de longitud, lanosas (raramente sólo ciliadas), los pelos unicelulares, blanquecinos a hialinos, rectos a sinuosos, adpresos o perpendiculares a la vaina, de base tuberculada, hasta de 8 mm de longitud. Lígula membranácea, cortamente ciliada, de ápice irregular, ca. 0.8 mm de longitud. Láminas hasta de 20 cm de longitud x 3-6 mm de ancho, lanosas (raramente glabrescentes), los pelos de la misma naturaleza de los de las vainas, planas, lineares, ápice largamente acuminado. Inflorescencias terminales, erectas, exertas a largamente exertas; pedúnculos ligeramente robustos, hasta de 50 cm de longitud, glabrescentes a lanosos o hirsutos, principalmente hacia su porción distal, los pelos de la misma naturaleza de los de las vainas y hasta de 3 mm de longitud; eje principal de 0.5-4 cm de longitud, glabrescente a lanoso; panojas constituidas por 3-4 racimos erectos, ascendentes, hasta de 10 cm de longitud, levemente distanciados entre si; raquis triquetro, glabrescente, en ocasiones laxamente piloso, principalmente hacia su porción proximal, los pelos de la misma naturaleza de los del pedúnculo, escabriúsculo marginalmente, ca. 0.5 mm de ancho; espiguillas bifloras, dispuestas en tríades, con pedicelos de diferente longitud, éstos planos a subtrígonos, flexuosos, escabriúsculos a corta o mediana y laxamente pilosos, los más largos hasta de 3 mm de longitud, los más cortos hasta de 1 mm de longitud; articulación pedicelo-espiguilla cupuliforme a discoide. Espiguillas 1.8-2.2 (-2.4) mm de longitud x ca. 1 mm de ancho, elipsoides, densamente pilosas, los pelos muy cortos aunque estipitados y claviformes, hasta de 0.25 mm de longitud; gluma inferior reducida a una minúscula membrana de menos de 0.1 mm de longitud, glabra, enervia; gluma superior casi tan larga como la lema inferior (sólo ca. 0.25 mm más corta que ésta), acuminada, 3-nervia; lema inferior de 1.8-2.2 (-2.4) mm de longitud, acuminada, (5--) 7-nervia, los nervios muy prominentes, pilosa en todos los espacios internervales; pálea inferior ca. 0.3 mm de longitud, membranácea, hialina; antecio superior tan largo o levemente más corto que la lema inferior (ca. 0.2 mm más corto), elipsoide, acuminado a ligeramente apiculado, estriado, cartilagíneo, glabro, castaño oscuro, brillante, las márgenes de la lema superior membranáceas, hialinas a blanquecinas, plegadas sobre la pálea superior y cubriéndola totalmente; flor perfecta; lodículas ca. 0.35 mm de longitud, hialinas, truncadas, de ápice levemente irregular. Cariopsis no vista.
Nombre vulgar. Paja de sabana (Meta).
Distribución geográfica y ecológica. Digitaria cardenasiana es endémica de Colombia y hasta ahora sólo se conocen registros de los departamentos de Boyacá, Meta, Santander y Vichada, entre los 100 y 2300 m y crece en sabanas orinocenses dominadas por Trachypogon (Poaceae: Andropogoneae) y en ambientes abiertos andinos de áreas secas y rocosas de la cordillera Oriental, correspondientes a matorrales con baja cobertura vegetal, dominados principalmente por Dodonaea viscosa, y en segunda medida por varias especies de gramíneas (Andropogon spp., Bothriochloa sp., Eragrostis spp., Sporobolus spp. y Stipa sp.).
Observaciones. Digitaria cardenasiana es afín y similar a D. killeenii A. S. Vega & Rúgolo, una especie endémica de Bolivia. No obstante, éstas pueden reconocerse básicamente por el menor porte de D. cardenasiana (hasta 0.6 m de altura vs 1.8-2 m en D. killeenii), por el menor tamaño de sus espiguillas (1.8-2.4 mm long. vs 2.8-3.2 mm en D. killeenii) y por la pálea inferior (glabra en D. cardenasiana vs pilosa en D. killeenii). Esta especie también es afín a D. andicola (véanse las observaciones dadas para D. andicola, en donde se mencionan las características más importantes para separar estas dos especies).
Las poblaciones de Boyacá (Giraldo-Cañas et al. 3556, 2300 m de altitud; Méndez & Becerra 262, 2220 m de altitud) presentan vainas y láminas glabrescentes y/o vainas sólo ciliadas, mientras que los demás ejemplares de tierras bajas (Meta y Vichada) presentan vainas y láminas lanosas (se excluye de esta comparación la población de Santander, pues el único espécimen de este departamento carece de hojas). Esto podría estar relacionado con las condiciones ecológicas de las sabanas orinocenses, sujetas a estrés hídrico y altas temperaturas, ya que la pubescencia en las estructuras vegetativas y/o reproductivas podría modificar el balance energético de la planta, reduciría el intercambio de calor y la difusión del vapor de agua al reducir la velocidad de flujo del aire deshidratante (véase Cutler 1987: 99), tal como ha sido destacado para otra especie de la tribu Paniceae, Axonopus aureus P. Beauv. (Giraldo-Cañas 2001b), la cual crece tanto en sabanas orinocenses como en ambientes andinos. De la misma manera, D. cardenasiana presenta pedúnculos glabrescentes a lanosos o hirsutos, principalmente hacia su porción distal, siendo los pelos unicelulares y de base tuberculada. Así, esta especie es la única entre las especies colombianas de Digitaria que puede presentar cañas floríferas lanosas a hirsutas.
Ejemplares examinados
COLOMBIA. Boyacá: Mun. Ráquira, cerca de los hornos de los artesanos, en inmediaciones del camino que conduce al desierto de La Candelaria; matorrales de áreas secas y rocosas, con baja cobertura vegetal, dominados principalmente por Dodonaea viscosa, y en segunda medida por varias especies de gramíneas (Andropogon spp., Bothriochloa sp., Digitaria cardenasiana, Eragrostis spp., Sporobolus spp. y Stipa sp.), ca. 2300 m, 19 jul 2003, D. Giraldo-Cañas et al. 3556 (COL). Mun. Ráquira, alrededores del pueblo, en suelos erosionados, 2220 m, 6 feb 1989, A. Méndez & C. Becerra 262 (COL). Meta: Hato Horizontes, sabana de Trachypogon vestitus, altillanura plana, 6 jun 1963, Blydenstein 955 (COL). Carimagua, 3 abr 1972, J. Hilton 9 (COL). Santander: Mun. Curití, ca. 1000 m, 17 ago 2004, O. Rangel s. n. (COL: 505913). Vichada: ca. 8 km E of Las Gaviotas along the unimproved dirt road to Santa Rita, completely open Trachypogon savanna with forest along spring fed stream, 220 m, 25 dic 1973, G. Davidse & F. Llanos 5145 (COL). 31 km W of Las Gaviotas along road to Puerto Gaitán, open treeless Trachypogon vestitus savanna, 210 m, 30 dic 1973, G. Davidse & F. Llanos 5372 (COL).
14. Digitaria filiformis (L.) Koeler, Descr. Gram. 26. 1802. Panicum filiforme L., Sp. Pl. 1: 57. 1753. LECTOTIPO. Norteamérica. Kalm s.n. (LINN, no visto, designado por Hitchcock, Contr. U.S. Natl. Herb. 12 (3): 117. 1908). Fig. 20.
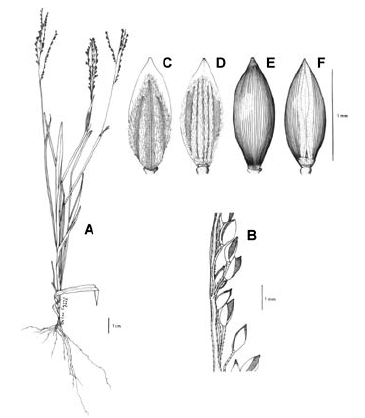
Figura 20. Digitaria filiformis (L.) Koeler: A. Hábito; B. Porción de un racimo; C. Espiguilla, vista desde la gluma superior; D. Espiguilla, vista desde la lema inferior; E. Antecio superior, visto desde la lema superior; F. Antecio superior, visto desde la pálea superior. Todo de Stevenson 653 (COL).
Plantas anuales, cespitosas; follaje laxo. Cañas erguidas, simples o ramificándose en los nudos basales, hasta de 70 cm de alto; entrenudos regularmente 3-6, pajizos, estriados longitudinalmente, cilíndricos a levemente colapsados, hasta de 10 cm de longitud, siendo más cortos los entrenudos inferiores; nudos glabros, castaño-oscuros. Vainas hasta de 10 cm de longitud, levemente pilosas, los pelos unicelulares, hialinos, rectos, adpresos, ascendentes, de base tuberculada, hasta de 1.3 mm de longitud. Lígula membranácea a escariosa, entera o levemente erosa, glabra, 0.7-2 mm de longitud. Láminas hasta de 13 cm de longitud x 2-6 mm de ancho, escasamente pilosas sólo en la base, los pelos similares a los de las vainas, planas, lineares, ápice acuminado. Inflorescencias erectas, largamente exertas; pedúnculos capilares, hasta de 22 cm de longitud; eje principal 0.5-5 cm de longitud; panojas constituidas por 2-18 (-20) racimos erectos, ascendentes, hasta de 10 cm de longitud, próximos entre si a distanciados; raquis triquetro, glabrescente, escabriúsculo marginalmente, ca. 0.25 mm de ancho; espiguillas dispuestas en tríades, con pedicelos de diferente longitud, éstos filiformes, subcilíndricos a planos, flexuosos, glabros, escabriúsculos, los más largos hasta de 3.2 mm de longitud, los más cortos hasta de 1.5 mm de longitud; articulación pedicelo-espiguilla discoide. Espiguillas 1.2-1.7 mm de longitud, 0.5-0.7 mm de ancho, elipsoides; gluma inferior reducida a una minúscula membrana, glabra, entera a emarginada, enervia, ca. 0.1 mm de longitud; gluma superior ca. 0.2-0.3 mm más corta que la lema inferior, lanceolada, acuminada a redondeada, 3-nervia, moderadamente pilosa, los pelos claviformes, hialinos, adpresos o ligeramente divergentes, ca. 0.25 mm de longitud en promedio, aunque los distales hasta ca. 0.5 mm de longitud; lema inferior de 1.2-1.7 mm de longitud, lanceolada, acuminada, 7-nervia, moderadamente pilosa, los pelos como los de la gluma superior; pálea inferior ca. 0.2 mm de longitud, escariosa, hialina a pajiza; lodículas reducidas, apenas perceptibles, menores que la pálea inferior; antecio superior tan largo como la lema inferior o apenas ca. 0.15 mm más largo que ésta, elipsoide, levemente apiculado, estriado, cartilagíneo, glabro, castaño oscuro, brillante, las márgenes de la lema superior membranáceas, blanquecinas, plegadas sobre la pálea superior y cubriendo cerca de 1/3 de ésta, lodículas ca. 0.2 mm de longitud, truncadas. Cariopsis no vista.
Distribución geográfica y ecológica. Digitaria filiformis crece en el continente americano en los Estados Unidos de América, México, Bolivia, Brasil, Paraguay y Argentina (Hitchcock 1935, Rúgolo de Agrasar 1974, 1994, Beetle 1987, Vega & Rúgolo de Agrasar 2002a, 2002b). De Colombia sólo se conocen tres colecciones, una del departamento del Magdalena, otra del departamento del Meta y una tercera sin localidad. D. filiformis es una especie poco frecuente y crece en lugares abiertos y húmedos de tierras bajas.
Observaciones. En vista de su condición anual y de su aspecto grácil, D. filiformis podría ser confundida con D. violascens. Sin embargo, ambas especies pueden reconocerse fácilmente por la naturaleza del raquis (triquetro y ca. 0.25 mm de ancho en D. filiformis vs. alado y 0.6-0.8 mm de ancho en D. violascens), por la articulación pedicelo-espiguilla (discoide en D. filiformis vs. cupuliforme en D. violascens) y por el tipo de pelos de las espiguillas (claviformes en D. filiformis vs. verrugosos en D. violascens).
Ejemplares examinados
COLOMBIA. Magdalena: Santa Marta, ca. 30 m, años 1898-1901, H. Smith 180-B (COL-253681; la recolección H. Smith 180 corresponde a un isotipo de D. adscendens var. rhachiseta Henrard, pero está mezclado con una planta de D. filiformis, por lo que se procedió a asignarle a esta última el número 180-B). Meta: cercanías del río Melúa, por la carretera de Los Japoneses, finca La Florida, en sabana, ca. 500 m, nov 1993, P. Stevenson 653 (COL). Sin departamento ni localidad: años 1783-1808, J. Mutis 6090 (COL, dos ejemplares).
15. Digitaria lehmanniana Henrard, Blumea 1 (1): 107. 1934. TIPO. Colombia. Cauca: forests of highlands of Popayán, 1500-2000 m, año 1886, Lehmann BT 632 (holotipo L; isotipos BAA, K, no vistos). Fig. 21.
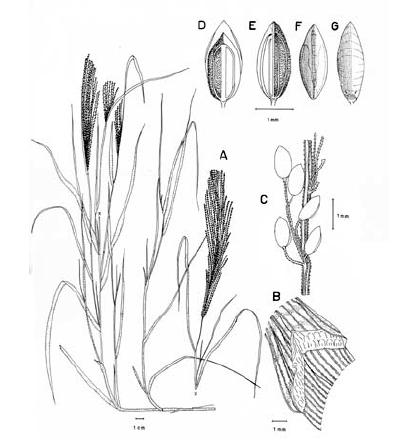
Figura 21. Digitaria lehmanniana Henrard: A. Hábito; B. Región ligular; C. Porción de un racimo; D. Espiguilla desde el lado de la gluma superior; E. Espiguilla desde el lado de la lema inferior; F. Antecio superior desde el lado de la pálea superior; G. Antecio superior desde el lado de la lema superior. Todo de Fuentes & Amaya 764 (COL).
Plantas anuales (?) (ejemplares estudiados sin rizomas ni raíces), follaje laxo a medianamente denso, caulinar. Cañas decumbentes en la base y con raíces en los nudos inferiores, con las porciones distales erectas, conspicuamente ramificadas, hasta de 70 cm de longitud; entrenudos numerosos, hasta 9, pajizos a verde-claros, levemente estriados a más comúnmente lisos, cilíndricos a colapsados, hasta de 6 cm de longitud, siendo los entrenudos basales similares en longitud a los distales; nudos glabros, castaño-oscuros. Vainas hasta de 12 cm de longitud, glabras. Lígula membranácea, glabra, con la porción distal levemente sinuosa, ca. 2 mm de longitud. Láminas hasta de 18 cm de longitud x 3-6 mm de ancho, glabras a glabrescentes, ocasionalmente sólo pilosas cerca de la región ligular, entonces los pelos escasos, hialinos, flexuosos, de base tuberculada, hasta de 5 mm de longitud, planas, lineares, ápice acuminado. Inflorescencias terminales, erectas, exertas; pedúnculos capilares, hasta de 6 cm de longitud; eje principal de 7-16 cm de longitud; panojas muy densas, constituidas por numerosos racimos, hasta 40 racimos por panoja, éstos erectos, ascendentes, hasta de 11 cm de longitud, próximos entre si; raquis capilar-filiforme, glabrescente, escabriúsculo marginalmente, ca. 0.25 mm de ancho; espiguillas dispuestas en tríades, en tétradas o en grupos de 5 por nudo, con pedicelos de diferente longitud, éstos capilares a filiformes, flexuosos, glabrescentes, escabriúsculos, los más largos hasta de 3 mm de longitud, los más cortos hasta 0.8 mm de longitud. Espiguillas ca. 1.2 mm de longitud x ca. 0.5 mm de ancho, ovadas a elipsoides, agudas, pilosas, los pelos estipitados, claviformes, adpresos a levemente divergentes, hialinos, hasta de 0.25 mm de longitud; gluma inferior ausente o reducida a una minúscula membrana, glabra, escariosa, enervia, de menos de 0.1 mm de longitud; gluma superior de 0.9-1 (-1.2) mm de longitud, membranácea, aguda, 3-nervia, los nervios en ocasiones anastomosados hacia su porción distal; lema inferior de 1.2 mm de longitud, membranácea, aguda, 7-nervia, los nervios en ocasiones anastomosados hacia su porción distal; pálea inferior ca. 0.2 mm de longitud; antecio superior tan largo como la lema inferior, ovado a elipsoide, agudo a acuminado, estriado, cartilagíneo, glabro, castaño claro, brillante, las márgenes de la lema superior membranáceas, blanquecinas a hialinas, plegadas sobre la pálea superior y cubriéndola completamente; lodículas ca. 0.2 mm de longitud, membranáceas a escariosas, hialinas, truncadas. Cariopsis no vista.
Distribución geográfica y ecológica. Digitaria lehmanniana es una especie propia de Sudamérica [Colombia, Venezuela, Ecuador, Brasil, Perú, Bolivia, Paraguay y Argentina (Vega & Rúgolo de Agrasar 2002a, Zuloaga et al. 2003)]. De Colombia sólo se conoce de los departamentos de Cauca y Meta y crece en áreas montanas, en sabanas y en cultivos, entre los 400 y 1800 m.
Observaciones. Digitaria lehmanniana es única entre las especies colombianas de Digitaria sect. Ternatae, pues presenta panojas ricamente ramificadas hasta con 40 racimos, mientras que en las demás especies de la sección las panojas no superan en ninguno de los casos los 20 racimos. Por otra parte, D. lehmanniana presenta las espiguillas más pequeñas de todas las especies colombianas del género y su antecio superior es castaño claro. Así, estas características permiten reconocer fácilmente esta especie.
Ejemplares examinados
COLOMBIA. Meta: Mun. Granada, ca. 400 m, asociada con cultivos de arroz, 10 jul 1999, C. Fuentes & J. Amaya 764 (COL, tres ejemplares).
Digitaria sect. Trichachne (Stapf) Henrard
16. Digitaria californica (Benth.) Henrard, Blumea 1 (1): 99. 1934. Panicum californicum Benth., Bot. Voy. Sulphur: 55. 1840. TIPO. México. Baja California Sur, año 1841, Hinds s.n. [holotipo K; isotipo US, no vistos). Fig. 22.
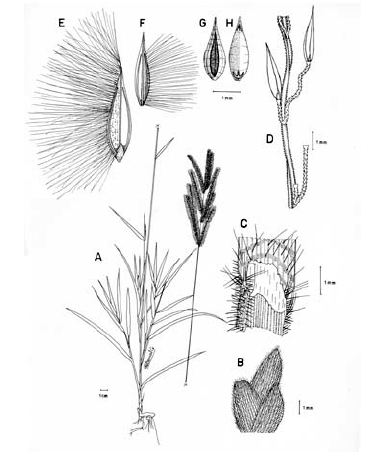
Figura 22. Digitaria californica (Benth.) Henrard: A. Hábito; B. Detalle de los catafilos; C. Región ligular; D. Porción de un racimo; E. Espiguilla desde el lado de la lema inferior; F. Espiguilla desde el lado de la gluma superior; G. Antecio superior desde el lado de la pálea superior; H. Antecio superior desde el lado de la lema superior. Todo de García-Barriga 14185 (COL).
Plantas perennes, cespitosas; rizomas cortos, engrosados, cubiertos de catafilos densa y cortamento pilosos; follaje medianamente denso a denso, caulinar. Cañas erguidas, conspicuamente ramificadas, hasta de 80 cm de altura; entrenudos 5-12, pajizos, cilíndricos, estriados longitudinalmente a canaliculados, hasta de 10 cm de longitud, siendo los entrenudos basales más cortos que los distales; nudos glabros, castaño claros. Vainas hasta de 23 cm de longitud, aunque normalmente no superan los 12 cm de longitud, pilosas, los pelos de base tuberculada, hialinos a blanquecinos, hasta de 4.5 mm de longitud. Lígula membranácea a escariosa, glabra, con la porción distal levemente irregular o denticulada a laciniada, 1.3-2.2 mm de longitud. Láminas hasta de 13 cm de longitud x 3-6 mm de ancho, planas, pilosas, con pelos de la misma naturaleza de los de las vainas, lanceoladas a lineares, ápice acuminado. Inflorescencias terminales, erectas, exertas; pedúnculos hasta de 44 cm de longitud; eje principal de 0.2-15 cm de longitud; panojas densas, constituidas por (2-) 6-16 racimos, éstos contraídos, ascendentes, muy próximos entre sí a más comúnmente alternos a lo largo del eje, hasta de 12 cm de longitud; raquis trígono, glabro, escabriúsculo a escabroso, 0.3-0.5 mm de ancho; espiguillas binadas, con pedicelos de diferente longitud, éstos subtrígonos, flexuosos, escabriúsculos, los más largos hasta de 3 mm de longitud, los más cortos hasta de 1 mm de longitud; articulación pedicelo-espiguilla discoide a cupuliforme. Espiguillas 3-3.5 mm de longitud x ca. 0.9 mm de ancho, ovadas, ápice acuminado, densa y largamente pilosas, los pelos simples, lanosos, blanco-plateados, divergentes, hasta de 4.5 mm de longitud; gluma inferior triangular, glabra, 0.6-0.75 mm de longitud; gluma superior triangular, tan larga como la lema inferior o un poco más corta que ésta, 3-nervia; lema inferior de 3-3.5 mm de longitud, triangular, 5-7-nervia; pálea inferior ausente a reducida, ca. 0.2 mm de longitud; antecio superior 0.5-0.8 mm más corto que la lema inferior, ovado, acuminado a apiculado, estriado, cartilagíneo, glabro, pajizo a cobrizo, brillante, lema superior enervia, las márgenes de la lema superior membranáceas, hialinas, plegadas sobre la pálea superior y cubriendo cerca de 4/5 de ésta; lodículas ca. 0.25 mm de longitud, truncadas. Cariopsis no vista.
Nombre vulgar. Rabo de zorro (Norte de Santander).
Distribución geográfica y ecológica. Digitaria californica se distribuye desde Norteamérica hasta Bolivia, Paraguay y el norte de Argentina y las Antillas. Esta especie es propia de suelos arenosos, taludes, cárcavas, arbustales y matorrales, principalmente de áreas xéricas y bosques secos y en Colombia está ampliamente distribuida y crece entre el nivel del mar y los 1600 m.
Observaciones. Con base en observaciones de material de herbario, pude advertir que D. californica es confundida con alguna frecuencia con Paspalum saccharoides Nees ex Trin. No obstante, ambas especies pueden separarse fácilmente por las espiguillas (binadas y ovadas en D. californica vs. solitarias y linear-lanceoladas en P. saccharoides), gluma inferior (presente en D. californica vs. ausente en P. saccharoides), por la lema inferior (5-7-nervia en D. californica vs. 2-nervia en P. saccharoides), por el antecio superior (0.5-0.8 mm más corto que la lema inferior, cartilagíneo y estriado-papiloso en D. californica vs. 0.6-1.1 mm más corto que la lema inferior, cartáceo y liso en P. saccharoides) y por las márgenes de la lema superior (membranáceas y plegadas sobre la pálea superior en D. californica vs. cartáceas y enrolladas sobre las orillas de la pálea superior en P. saccharoides). Véanse las observaciones dadas para D. similis.
Ejemplares examinadosCOLOMBIA. Boyacá: Mun. Boavita, cañón del Chicamocha, 1550 m, 0 may 1991, A. Etter & L. Baptiste 691 (COL). Cundinamarca: Mun. Nilo, carretera Nilo-Melgar, en coluvios de los cortes de la carretera, ca. 200 m, 19 oct 2004, D. Giraldo-Cañas et al. 3795 (COL). 3 km de Girardot, entre Girardot y Tocaima, 330 m, 3 dic 1965, E. Forero & J. Garzón 238 (COL). Tocaima, carretera Tocaima-Pubenza, 350-400 m, 14 abr 1952, H. García 14185 (COL). Mun. Nariño, 500 m, jul 1930, E. Pérez-Arbeláez 360 (COL, dos ejemplares). Entre Viotá y Girardot, 320-560 m, ago 1964, C. Saravia 4581 (COL). In the valley of the Río Sumapaz, between Pandi y Boquerón, 600 m, 25 mayo 1985, J. Wood 4859 (COL). Huila: Mun. Villavieja, desierto La Tatacoa, El Cuzco, alrededores del Observatorio Astronómico, 15 abr 2003, Y. Figueroa & W. Calderón 95 (COL), 5 ene 2004, Y. Figueroa & N. García 493 (COL). Cordillera Oriental, La Bodega, ca. 750 m, 22 nov 1944, E. Little Jr. 8973 (COL). La Guajira: Mun. Barrancas, in xeric location, near Roche, El Cerrejón, 13 mar 1981, P. Bunch et al. 460 (HUA). Norte de Santander: Cúcuta y El Rosario, en área árida, feb 1941, Carvajalino & Díaz 40 (COL). Santander: Mun. Soatá, cañón del Chicamocha, Puente Pinzón, 1550 m, 16 oct 1992, A. Etter & L. Villa 612 (COL). Mun. Boavita, cañón del Chicamocha, 1550 m, 7 may 1991, A. Etter et al. 691 (COL). Bajada desde San Gil al cañón del Chicamocha, El Pescadero, 750-1000 m, 8 ago 1987, J. Fernández & L. Caballero 7214 (COL). Entre San Gil y el cañón del Chicamocha, 13 jun 1962, C. Saravia et al. 745 (COL). Entre Pescadero y Piedecuesta, 1005 m, 15 jun 1962, C. Saravia et al. 876 (COL). Entre Aratoca y la carretera Piedecuesta, ca. 1400 m, 16 jun 1962, C. Saravia et al. 922 (COL). Tolima: entre El Espinal y Girardot, 350 m, 26 mar 1939, A. Alston 7697 (COL). Valle del Cauca: Loboguerrero, zona subxerofítica, 940 m, mar 1981, I. Armbrecht s.n. (HUA: 56647), 12-14 nov 1962, C. Saravia 1580 (COL).
17. Digitaria insularis (L.) Fedde, Just’s Bot. Jahresber. 31: 778. 1904. Andropogon insularis L., Syst. Nat. (ed. 10): 1304. 1759. LECTOTIPO. Jamaica, P. Browne s.n. (LINN, no visto, designado por Hitchcock, Contr. U.S. Natl. Herb. 12: 126. 1908). Fig. 23.
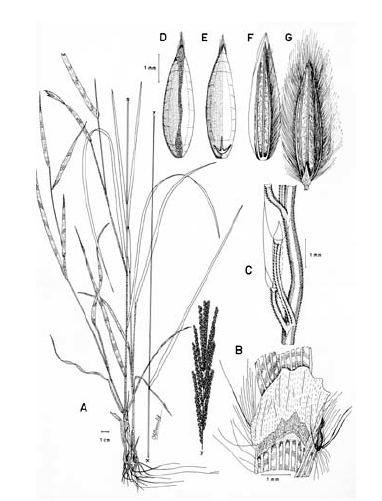
Figura 23. Digitaria insularis (L.) Fedde: A. Hábito; B. Región ligular; C. Porción de un racimo; D. Antecio superior desde el lado de la pálea superior; E. Antecio superior desde el lado de la lema superior; F. Espiguilla desde el lado de la gluma superior; G. Espiguilla desde el lado de la lema inferior. Todo de Giraldo-Cañas 3296 (COL).
Plantas perennes, densamente cespitosas; rizomas cortos, engrosados, cubiertos de catafilos corta y densamente pilosos; follaje medianamente denso a denso, caulinar. Cañas erguidas, ramificadas, hasta de 1.8 m de altura; entrenudos 3-10, pajizos, cilíndricos, estriados longitudinalmente a canaliculados, hasta de 12 cm de longitud, siendo los entrenudos basales más cortos que los distales; nudos glabros, castaño claros a oscuros. Vainas hasta de 30 cm de longitud, glabras o escasamente pilosas, los pelos de base tuberculada, hialinos a blanquecinos, hasta de 2.5 mm de longitud. Lígula membranácea a escariosa, glabra, con la porción distal levemente irregular, 2.2-3.5 mm de longitud. Láminas hasta de 30 cm de longitud x (2-) 8-15 mm de ancho, planas, glabras o sólo laxamente pilosas hacia su porción basal, con pelos de la misma naturaleza de los de las vainas, lineares a lanceoladas, ápice acuminado. Inflorescencias terminales, erectas, exertas; pedúnculos hasta de 80 cm de longitud; eje principal de 2-20 (-25) cm de longitud; panojas laxas a más comúnmente densas, constituidas por (3-) 10-50 racimos, éstos contraídos, ascendentes, alternos a lo largo del eje, hasta de 15 cm de longitud, los racimos inferiores en ocasiones ramificados; raquis trígono, glabro, escabriúsculo a escabroso, ca. 0.3 mm de ancho; espiguillas binadas, con pedicelos de diferente longitud, éstos subtrígonos, flexuosos, escabriúsculos a escabrosos, los más largos hasta de 5 mm de longitud, los más cortos hasta de 1.5 mm de longitud; articulación pedicelo-espiguilla discoide a cupuliforme. Espiguillas 3.7-4.5 (-4.8) mm de longitud x ca. 1 mm de ancho, fácilmente caducas, lanceoladas, ápice acuminado, densa y largamente pilosas, los pelos simples, lanosos, ocráceos, divergentes, hasta de 4 mm de longitud; gluma inferior triangular a ovada, glabra, 0.6-0.8 mm de longitud, enervia; gluma superior lanceolada, acuminada, un poco más corta que la lema inferior, ca. 0.3 mm más corta, 3-5-nervia; lema inferior de 3.7-4.5 (-4.8) mm de longitud, lanceolada, acuminada, 5-7-nervia; pálea inferior reducida, ca. 0.2 mm de longitud; antecio superior ca. 0.4 mm más corto que la lema inferior, lanceolado, acuminado a apiculado, estriado, cartilagíneo, glabro, pajizo a cobrizo, brillante, lema superior enervia, las márgenes de la lema superior membranáceas, hialinas, plegadas sobre la pálea superior y cubriendo cerca de 4/5 de ésta; lodículas ca. 0.4 mm de longitud, cordiformes. Cariopsis ca. 1.8 mm de longitud, comprimida dorsiventralmente; escudete embrional ca. ½ de la longitud de la cariopsis; hilo aovado.
Nombres vulgares. Cola de zorro (Cundinamarca), hierba de zorra (Magdalena), paja de burro (Huila), paja de zorro (La Guajira), rabo de zorro (Nariño, Tolima, Valle del Cauca), John Taylor grass (San Andrés, Providencia y Santa Catalina).
Distribución geográfica y ecológica. Digitaria insularis es nativa del continente americano y se distribuye desde el sur de los Estados Unidos de América hasta la Argentina y las Antillas, y ha sido introducida en Asia tropical y en algunas islas del Pacífico (Pohl & Davidse 1994). Esta especie crece en sabanas de areniscas, en afloramientos rocosos y arenas blancas del Escudo Guayanés, en chagras (chacras), en potreros, en márgenes de caminos y carrteras, en áreas secas, en cultivos de caña de azúcar, cacao y café. Menos frecuentemente se halla en lugares sujetos a humedad. En Colombia se encuentra ampliamente distribuida y crece entre el nivel del mar y los 2300 m.
Observaciones. Digitaria insularis es fácilmente reconocible por sus espiguillas con indumento ocráceo, las cuales son fácilmente caducas. Por otra parte, es la única especie colombiana que en ocasiones presenta inflorescencias con los racimos proximales ramificados en órdenes consecutivos, situación que Rua (2003) no había documentado para esta especie en su estudio morfológico de Digitaria sect. Trichachne.Las plantas de esta especie no son consumidas por el ganado [según información consignada en los ejemplares Echeverry 992 (COL) y Marulanda & Betancur 2156 (COL, HUA)] y al parecer son sospechosas de causar intoxicación en bovinos [Hernández s.n. (COL: 98274)]. No obstante, Subils (1995: 31) comenta que es una buena forrajera natural, tanto para equinos como para mulares y vacunos. Vega & Rúgolo de Agrasar (2002a) mencionan que en Bolivia se utiliza para calmar la tos: se machaca la planta entera en agua y se bebe el líquido resultante.
Ejemplares examinados
COLOMBIA. Amazonas: Mun. Leticia, corregimiento departamental Puerto Santander, región de Araracuara, trocha a La Chorrera, terrazas altas, 250-300 m, 22 jul 1996, D. Cárdenas & A. Matapí 7336 (COAH, COL). Corregimiento departamental La Pedrera, en el sitio denominado “Chorro de Córdoba”, zona de raudales del río Caquetá, en afloramiento rocoso y arenas blancas del Escudo Guayanés, 230, 19 jul 1997, D. Giraldo-Cañas et al. 2718 (COAH, COL, HUA, SI). Río Caquetá, región de Araracuara, 1 abr 1976, C. Sastre & H. Reichel 5085 (COL). Río Caquetá, La Pedrera and vicinity, 3 oct 1952, R. Schultes & I. Cabrera 17715 (COL). Amazonas-Caquetá: Araracuara, chagra Bartolomé, ca. 200 m, 30 jul 1986, J. Battjes 103 (COAH). Antioquia: Mun. Medellín, predios internos del jardín Botánico “Joaquín Antonio Uribe”, 1550 m, 2 ene 2002, D. Giraldo-Cañas et al. 3296 (CAUP, COL, HUA). Mun. San Jerónimo, 8 sep 1930, W. Archer 626 (COL). Mun. San Jerónimo, km 58 de las partidas de Sopetrán, 750 m, 24 sep 1986, R. Pohl 15589 (HUA). Bolívar: estación experimental El Carmen, 6.4 km al sur de El Carmen, 2 feb 1962, C. Saravia & D. Johnson 4 (COL). Caquetá: Mun. Solano, Medio Caquetá, chagra de Óscar Román, terraza baja bien drenada, 1 oct 1990, G. Vélez 308 (COAH). Cauca: río Patía, 1000 m, 11 mayo 1935, H. García 4489 (COL). Mun. Patía, a 2 km de Mojarras, granja de la Universidad de Nariño, 700 m, 23 abr 1988, B. Ramírez 1128 (COL, PSO). Alrededores de la población de Mojarras, 650 m, 20 may 1989, B. Ramírez 1496 (COL, PSO). Cundinamarca: Mun. Nilo, hacienda La Guaira, río Pagüey, márgenes del camino interno principal de la hacienda, 350 m, 16 oct 2004, D. Giraldo-Cañas et al. 3767 (COL). Girardot, finca Los Chircales, 326 m, 25 nov 1973, P. Dávila 28 (HFAB). La Esperanza, 1280 m, 20 may 1946, J. Duque-Jaramillo 3642 (COL). Poblado de Nariño, carretera Los Mangos, 340 m, 15 feb 1986, J. Fernández & R. Jaramillo 5249 (COL). Mun. Nariño, quebrada del Apauta, 400 m, 26/28 mar 1986, J. Fernández & R. Jaramillo 6273 (COL). 3 km de Girardot, entre Girardot y Tocaima, 330 m, 3 dic 1965, E. Forero & J. Garzón 238-A (COL). La Esperanza a San Javier, camino de herradura y línea del ferrocarril a Girardot, ca. 1150 m, 15 ene 1947, H. García 12183 (COL). Provincia de Río Negro, Mun. Pacho, carretera Pacho-La Capilla, alrededores del estadio municipal de fútbol, ca. 1600 m, 13 ene 2004, D. Giraldo-Cañas et al. 3729 (COL). Girardot, hacienda Potrerillo, 16 abr 1964, H. Hernández s.n. (COL: 98274). Tocaima, dic 1932, E. Pérez-Arbeláez 2430 (COL). Pandi, carretera Pandi-Sutatausa, finca La Aguablanca, 1024 m, 26 abr 1970, R. Tamayo 19 (HFAB). La Mesa, 1300 m, dic 1856, J. Triana s.n. (COL: 2585). On the valley of the Río Sumapaz, between Pandi and Boquerón, 600 m, 25 may 1985, J. Wood 4861 (COL). Huila: embalse de Betania, península entre brazos Yaguará y Magdalena, frente al campamento santa Elena, brazo Yaguará, 520 m, 14 jun 1987, C. Becerra et al. 208 (COL). La Bodega, cordillera Oriental, ca. 750 m, 22 nov 1944, E. Little Jr. 8974 (COL). Neiva, 472 m, 27 sep 1969, S. Mosquera 10 (HFAB). La Guajira: Mun. Fonseca, corregimiento Distracción, Doscaminos, finca El Carmen, 500 m, 1 sep 1990, O. Marulanda & J. Betancur 2156 (COL, HUA). Rumbo a San Juan del Cesar, 4 km de Distracción, 100-200 m, 18 ene 1963, C. Saravia 2103 (COL). Magdalena: valle del río Cesar, parte occidental, al oeste de Los Venados, 60 m, 31 sep 1961, A. Dugand 5826 (COL). Región de Santa Marta, en Ciénaga, inmediaciones del río Córdoba, 25 feb 1950, R. Romero 2035 (COL). Rumbo a Codazzi, 20 km al sur de La Paz, 3 nov 1963, C. Saravia 2950 (COL, UPTC). Santa Marta, ca. 5 m, años 1898-1901, H. Smith 193 (COL). Santa Marta, paque nacional Tairona, carretera troncal del Caribe, 20 m, nov 1991, P. Torrijos & J. Martínez 198 (COL). Meta: Mun. Guamal, vereda El Encanto, 470 m, 9 mar 1989, R. Sánchez 1114 (COL). Nariño: El Manzano, ca. 1100 m, 1 abr 1966, H. Martínez & C. Martínez 22 (COL). Mun. Pasto, corregimiento de Chachagüí, km 38, túnel al aeropuerto, 1800 m, 7 nov 1987, B. Ramírez 1065 (COL, PSO). Mun. Pasto, corregimiento de Chachagüí, carretera Panamericana, puente de Juanambú, 950 m, 23 abr 1988, B. Ramírez 1111 (CAUP, COL, PSO). Carretera Pasto-Túquerres, ca. 2300 m, 30 nov 1962, C. Saravia & R. Jaramillo 1858 (COL). San Andrés, Providencia y Santa Catalina: isla de Providencia, camino de la montaña al aeropuerto, 22 jul 1990, P. Lowy & F. González 197 (COL). Isla de San Andrés, inmediaciones del Poblado, 363 m, 17 jul 1966, J. Torres et al. 192 (COL). Isla de Providencia, Smooth Water Bay, South Hill, Archibolds Point, Rocky Point Manchioneal Bay, Alligator Point, 19/25 jul 1966, J. Torres et al. 296 (COL). Isla de San Andrés, cerca de la fábrica de grasa de coco “El Cove”, 26/31 jul 1966, J. Torres et al. 425 (COL). Santander: Between San Gil and Barichara, 1500 m, año 1983, J. Wood 3877 (COL, dos ejemplares). Chicamocha Valley, 4 km N of Pescadero towards Bucaramanga, 700 m, 15 sep 1985, J. Wood 5070 (COL). Tolima: El Espinal-Girardot, 350 m, 26 mar 1939, A. Alston 7694 (COL). Armero, carretera Armero-Mariquita, 421 m, 24 oct 1976, A. Ángel 12 (HFAB). Valle del río Magdalena, Armero, 400 m, 6 oct 1940, J. Cuatrecasas 10497 (COL). Mun. El Espinal, Molino San Martín, 400-450 m, 24 may 1964, R. Echeverry 992 (COL). Mun. Armero, antiguo San Lorenzo, 380 m, 13/15 may 1939, H. García 7526 (COL). El Espinal, 500 m, dic 1859, M. Lindig 1104-103 (COL). Valle del Cauca: cerro Tres Cumbres, en los alrededores de Cali, 1100 m, 1 dic 1947, L. Bermúdez & F. Barkley 17-C-823 (COL). Entre Gorgona y Cabuyal, márgenes del camino, 1000 m, 2 jun 1943, J. Cuatrecasas 14500 (COL). Sin localidad, 1000 m, jun 1938, J. Duque-Jaramillo 4409 (COL). Mun. Palmira, 1050 m, may 1935, H. García 3173 (COL). Entre Cerrito y Palmira, hacienda El Trejo, 1050 m, 28 dic 1938, H. García 6293 (COL). Palmira, Facultad de Agronomía, 1020 m, 11 abr 1962, J. Idrobo 4989 (COL). Buga, riverbank weeds along Río Guadalajara, 15 jun 1982, H. Murphy 581 (COL, dos ejemplares). Entre Dagua y Loboguerrero, 1050 m, 10/12 nov 1962, C. Saravia 1439 (COL). Yumbo, entre Pubenza y Mulato, 1100 m, 21 nov 1962, C. Saravia 1726 (COL). Vaupés: Río Kuduyarí, tributary of Río Vaupés, middle and lower course, ca. 250 m, 16 oct 1952, R. Schultes & I. Cabrera 17867 (COL). Sin departamento ni localidad: años 1783-1808, J. Mutis 5507 (COL).
18. Digitaria similis Beetle ex Gould, J. Arnold Arbor. 60 (2): 320. 1979. Basado en Trichachne affinis Swallen, Rhodora 65: 355. 1963. TIPO. República Dominicana, Prov. Santiago, Quinqua, Valle del Cibao, 21oct 1930, E. L. Ekman 16090 (holotipo US, no visto; isotipo SI!). [Digitaria similis Beetle, Phytologia 37 (4): 344. 1977; el binomio propuesto por Beetle es un nom. nov. y un nom. inval., ya que no presentó la referencia directa de la página y la fecha de publicación del sinónimo reemplazado (T. affinis Swallen)]. Fig. 24.
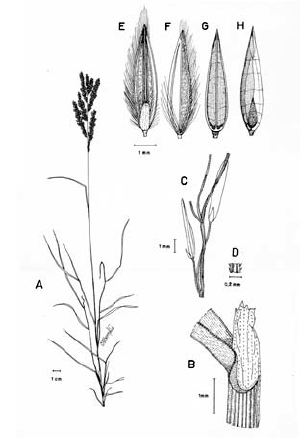
Figura 24. Digitaria similis Beetle ex Gould: A. Hábito; B. Región ligular; C. Porción de un racimo; D. Detalle del raquis; E. Espiguilla desde el lado de la lema inferior; F. Espiguilla desde el lado de la gluma superior; G. Antecio superior desde el lado de la lema superior; H. Antecio superior desde el lado de la pálea superior. Todo de Etter 610 (COL).
Plantas perennes (?) (ejemplares examinados sin rizomas ni raíces), gráciles; follaje laxo, caulinar. Cañas erguidas o levemente geniculadas en la base, ramificadas, hasta de 0.45 m de altura; entrenudos 3-6, pajizos, cilíndricos, estriados longitudinalmente a canaliculados, hasta de 7 cm de longitud, siendo los entrenudos basales más cortos que los distales; nudos glabros, castaños. Vainas hasta de 10 cm de longitud, glabrescentes. Lígula membranácea a escariosa, glabra, con la porción distal erosa a irregular, 2-3 mm de longitud. Láminas hasta de 11 cm de longitud x 1-3 mm de ancho, planas, glabrescentes, lineares, ápice acuminado. Inflorescencias terminales, erectas, exertas; pedúnculos hasta de 18 cm de longitud; eje principal de 8 cm de longitud; panojas medianamente densas, constituidas por 6-8 racimos, éstos contraídos, ascendentes, alternos a lo largo del eje, hasta de 6 cm de longitud; raquis trígono, glabro, escabriúsculo a escabroso, ca. 0.25 mm de ancho; espiguillas binadas, con pedicelos de diferente longitud, éstos subtrígonos, flexuosos, escabriúsculos a escabrosos, los más largos hasta de 6 mm de longitud, los más cortos hasta de 2.8 mm de longitud; articulación pedicelo-espiguilla discoide a cupuliforme. Espiguillas 4.1-5.1 mm de longitud x ca. 1 mm de ancho, lanceoladas, ápice acuminado, densa y largamente pilosas, los pelos simples, lanosos, blanquecinos, divergentes, hasta de 2 mm de longitud; gluma inferior lanceolada, ápice obtuso a truncado o levemente irregular, glabra, hialina, 0.5-1.4 mm de longitud; gluma superior lanceolada, ápice acuminado, ca. 1 mm más corta que la lema inferior, 3-5-nervia; lema inferior de 4.1-5.1 mm de longitud, lanceolada, ápice acuminado, 5-7-nervia; pálea inferior ausente; antecio superior tan largo como la espiguilla, lanceolado, acuminado, estriado, cartilagíneo, glabro, pajizo, brillante, lema superior 3-nervia, las márgenes de la lema superior membranáceas, hialinas, plegadas sobre la pálea superior y cubriéndola casi completamente; lódiculas ca. 0.4 mm de longitud, cordiformes. Cariopsis no vista.
Distribución geográfica y ecológica. Digitaria similis es nativa del continente americano y se distribuye en el Caribe (República Dominicana) y en Colombia, Brasil, Bolivia, Paraguay y Argentina (Zuloaga et al. 2003). De Colombia sólo se conocen dos ejemplares, uno del cañón del Chicamocha en la Cordillera Oriental (departamento de Boyacá), coleccionado en un bosque seco bajo ubicado a 1800 m de altitud, y el segundo de Santa Marta, coleccionado al nivel del mar (departamento del Magdalena).
Observaciones. Digitaria similis es fácilmente reconocible por sus grandes espiguillas (4.1-5.1 mm de longitud) y por ser la única entre las especies de Digitaria sect. Trichachne que presenta la lema superior 3-nervia. No obstante, por su indumento lanoso y blanquecino podría ser confundida con D. californica, de la que se diferencia por las espiguillas (lanceoladas y de 4.1-5.1 mm de longitud en D. similis vs. ovadas y de 3-3.5 mm en D. californica), por la gluma inferior (lanceolada en D. similis vs. triangular en D. californica) y por el antecio superior (lanceolado y tan largo como la lema inferior en D. similis vs. ovado y 0.5-0.8 mm más corto que la lema inferior en D. californica).
Ejemplares examinados
COLOMBIA. Boyacá: Mun. San Mateo, cañón del Chicamocha, Guicas-Buraga, 1800 m, en bosque seco bajo, 3 oct 1991, A. Etter et al. 610 (COL). Magdalena: Santa Marta, nivel del mar, años 1898-1901, H. Smith 200 (MO).
ESPECIES EXCLUIDAS
Digitaria argillacea (Hitchc. & Chase) Fernald: esta especie fue citada para Colombia por Zuloaga et al. (2003). No obstante, no se pudieron hallar ejemplares colombianos de la misma. Por otra parte, el espécimen E. Escobar 166 (VALLE!) de los Llanos Orientales del departamento del Meta estaba determinado como D. argillacea, pero éste realmente corresponde a la especie Axonopus purpusii (Mez) Chase. Quizás la cita de D. argillacea para Colombia esté basada en esta colección mal determinada.
Digitaria neesiana Henrard: esta especie fue citada para Colombia por Killeen (1990) y Escobar et al. (1993), pero no se detectaron ejemplares colombianos de ésta. Posteriormente, Killeen & Rúgolo de Agrasar (1992) consideran a esta especie como endémica del cerrado brasileño. Quizás la especie citada por Killeen (1990) y Escobar et al. (1993) corresponde a D. dioica. Véanse las observaciones dadas para esta última especie.
Digitaria fragilis (Steud.) Luces (= Paspalum fragile Steud.): esta especie fue citada para Colombia por Luces de Febres (1963) y por Renvoize (1998), pero no se detectaron ejemplares colombianos de ésta. La presencia de D. fragilis en territorio colombiano es dudosa ya que está basada en el ejemplar Funck & Schlim 724 (L) y éste es el holotipo de D. rhachitricha Henrard (= D. fragilis). Con relación a este ejemplar, Luces (1942: 82) hace una interesante anotación, la cual ayudaría a resolver el problema de la localidad de la recolección Funck & Schlim 724: “D. fragilis (Steud.) Luces, Journ. Wash. Acad. Sc. 32: 160. 1942. Descrita de Cumaná (Venezuela), Funck 724, tipo (D. rhachitricha Henr. Blumea 1: 95. 1934. Funck & Schlim 724, tipo). El espécimen en el Herbario de Leiden tiene por localidad Colombia. Funck 724, llamado Paspalum fragile, que se encuentra en París, tiene como localidad Cumaná, inscrita de la propia mano de Steudel. Otro especimen en París tiene la misma localidad. Los especímenes distribuidos por Funck & Schlim parecen ser de la misma recolección; a menudo no tienen localidad y algunos tienen como tal Colombia y otros Venezuela. Puede ser que Funck 724 y Funck & Schlim 724 sean diferentes colecciones, pero los especímenes son idénticos.” Por otra parte, en el reciente Catálogo de las gramíneas del Nuevo Mundo (Zuloaga et al. 2003: 202), esta especie no es considerada para Colombia. Asimismo, Vega & Rúgolo de Agrasar (2002a: 180) tampoco la citan para Colombia.
EJEMPLARES EXCLUIDOS
COLOMBIA. Vichada: Región Guayanesa, Mun. Puerto Carreño, afloramientos rocosos del tipo “lajas” entre Punta de Lajas y el cerro El Bita, ribera del río Orinoco, 40-100 m, 4-5 ene 2004, D. Giraldo-Cañas & C. Parra 3673, 3677 (COL).
Estos ejemplares son muy afines a D. horizontalis en virtud de las siguientes características: raquis laxamente piloso, siendo los pelos hialinos de base tuberculada; espiguillas binadas (aunque a menudo la espiguilla cortamente pedicelada aparece rudimentaria), de 2.1-2.5 mm de longitud, pilosas, siendo los pelos simples; gluma inferior apenas perceptible (ca. 0.1 mm de longitud); gluma superior ca. 1.5 mm de longitud, triangular y 3-nervia; lema inferior de 2.1-2.5 mm de longitud, 5-nervia; antecio superior tan largo como la lema inferior. No obstante, estos ejemplares poseen el antecio superior cobrizo oscuro [como el presentado por algunos ejemplares de D. californica y D. insularis], mientras que los típicos representantes de D. horizontalis lo presentan pajizo a castaño muy claro. Esta característica pone en duda la ubicación de los ejemplares Giraldo-Cañas & Parra 3673 y 3677 en dicha especie, pero tampoco se ajusta a ninguna de las especies conocidas del género. Por el momento, estos dos ejemplares quedan sin determinación taxonómica confiable.
AGRADECIMIENTOS
Al Instituto de Ciencias Naturales y a la Universidad Nacional de Colombia por todas las facilidades brindadas para la preparación de este trabajo. A C. A. Bernal (COL) y J. C. De Las Casas (COL) por la realización de las ilustraciones que acompañan esta contribución. A C. Parra (COL), E. Forero (COL), O. Rangel (COL) y O. Rivera (COL) por su valiosa colaboración. A J. L. Fernández (COL) y E. Linares (COL) por la valiosa evaluación de esta contribución. A F. Zuloaga (SI), O. Morrone (SI), S. Renvoize (K), S. Lægaard (AAU), Á. Idárraga (HUA), R. Ortiz-Gentry (MO), P. M. Peterson (US), J. F. Veldkamp (L), J. Palmer (CANB) y A. Van den Borre (CANB) por el obsequio de valiosa bibliografía. A los curadores de los herbarios CAUP, COAH, COL, HUA, JAUM, MEDEL, MO, PSO, RSA, SI y VALLE por los préstamos enviados o por su grata colaboración durante la visita a sus instalaciones. Esta contribución es derivada del proyecto “Estudios sistemáticos en gramíneas de Colombia: Partes I y II”, de la Dirección Nacional de Investigación de la Universidad Nacional de Colombia, sede Bogotá.
LITERATURA CITADA
1. ALISCIONI, S. & M. ARRIAGA. 1998. Estudio histofoliar comparado de las especies de los grupos Virgata y Quadrifaria del género Paspalum L. (Poaceae: Panicoideae: Paniceae). Candollea 53: 333-348. [ Links ]
2. ARRIAGA, M. 1987. Interpretación del ornamento del antecio de Eriochloa (Poaceae). Bol. Soc. Argent. Bot. 25: 131-141. [ Links ]
3. BEETLE, A. 1987. Las gramíneas de México, Tomo II. Secretaría de Agricultura y Recursos Hidráulicos-COTECOCA, México, D. F. [ Links ]
4. BOR, N. 1956. The genus “Digitaria” Heist. in India and Burma. Webbia 11: 301-367.
5. CÁMARA HERNÁNDEZ, J. 2001. Morfología de la inflorescencia de Digitaria sanguinalis (Poaceae). Bol. Soc. Argent. Bot. 36: 87-95. [ Links ]
6. CÁMARA HERNÁNDEZ, J. & G. RUA. 1991. The synflorescence of Poaceae. Beitr. Biol. Pflanzen 66: 297-311. [ Links ]
7. CHASE, A. 1911. Notes on genera of Paniceae, IV. [ Links ] Proc. Biol. Soc. Wash. 24: 103-160.
8. CHASE, A. & Z. LUCES DE FEBRES. 1972. Primer libro de las gramíneas. Instituto Interamericano de Ciencias Agrícolas de la OEA, Lima. [ Links ]
9. CIALDELLA, A. & A. VEGA. 1996. Estudios sobre la variación estructural de las espiguillas en géneros de la tribu Paniceae (Poaceae). Darwiniana 34: 173-182. [ Links ]
10. CIALDELLA, A., O. MORRONE & F. ZULOAGA. 1995. Revisión de las especies del género Paspalum (Poaceae: Panicoideae: Paniceae), grupo Bonplandiana. Darwiniana 33: 67-95. [ Links ]
11. CLARK, C. & F. GOULD. 1975. Some epidermal characteristics of paleas of Dichantelium, Panicum, and Echinochloa. Amer. J. Bot. 62: 743-748. [ Links ]
12. CLAYTON, W. & S. RENVOIZE. 1986. Genera Graminum: Grasses of the world. Kew Bull., Additional series 13: 1-389. [ Links ]
13. COCUCCI, A. 1980. Precisiones sobre la terminología sexológica aplicada a angiospermas. Bol. Soc. Argent. Bot. 19: 75-81. [ Links ]
14. COCUCCI, A. & A. ANTON. 1988. The grass flower: suggestions on its origin and evolution. Flora 181: 353-362. [ Links ]
15. CONNOR, H. 1981. Evolution of reproductive systems in the Gramineae. Ann. Missouri Bot. Gard. 68: 48-74. [ Links ]
16. CONNOR, H. 1987. Reproductive biology in the grasses. En: T. Soderstrom, K. Hilu, C. Campbell & M. Barkworth (eds.), Grass Systematics and Evolution: 117-132. Smithsonian Institution Press, Washington, D. C. [ Links ]
17. CONNOR, H., A. ANTON & M. ASTEGIANO. 2000. Dioecism in grasses in Argentina. En: S. Jacobs & J. Everet (eds.), Grasses: Systematics and Evolution: 287-293. CSIRO, Melbourne. [ Links ]
18. COSTEA, M. & F. TARDIF. 2002. Taxonomic of the most common weedy European Echinochloa species (Poaceae: Panicoideae) with special emphasis on characters of the lemma and caryopsis. Sida 20: 525-548. [ Links ]
19. CRINS, W. 1991. The genera of Paniceae (Gramineae: Panicoideae) in the southeastern United States. J. Arnold Arbor., suppl. ser. 1: 171-312. [ Links ]
20. CUTLER, D. 1987. Anatomía vegetal aplicada. Serie Biblioteca Mosaico, Librería Agropecuaria S. A., Buenos Aires. [ Links ]
21. DAVIDSE, G. & F. ZULOAGA. 1992. Two new species of Paspalum (Poaceae: Paniceae) from Venezuela and Colombia. Novon 2: 193-197. [ Links ]
22. DÁVILA, P. & L. CLARK. 1990. Scanning electron microscopy survey of leaf epidermis of Sorghastrum (Poaceae: Andropogoneae). Amer. J. Bot. 77: 499-511. [ Links ]
23. DÁVILA ARANDA, P. & E. MANRIQUE FORCECK. 1990. Glosario de términos agrostológicos. Cuadernos del Instituto de Biología 5, Universidad Nacional Autónoma de México UNAM, México, D. F. [ Links ]
24. DÁVILA ARANDA, P., J. SÁNCHEZ KEN & L. CABRERA MARTÍNEZ. 1993. Las gramíneas: características generales e importancia. Bol. Instituto de Botánica Universidad de Guadalajara (época 3) 1: 397-421. [ Links ]
25. DUVALL, M., J. NOLL & A. MINN. 2001. Phylogenetics of Paniceae (Poaceae). Amer. J. Bot. 88: 1988-1992. [ Links ]
26. EDWARDS, G. & C. BLACK. 1971. Isolation of mesophyll cells and bundle sheath cells from Digitaria sanguinalis (L.) Scop. Pl. Physiol. 47: 149-156.
27. ELLIS, R. 1979. A procedure for standardizing comparative leaf anatomy in the Poaceae. II. The epidermis as seen in surface view. Bothalia 12: 641-671.
28. ESCOBAR, E., J. BELALCÁZAR & G. RIPPSTEIN. 1993. Clave de las principales plantas de sabana de la altillanura de los llanos Orientales en Carimagua, Meta, Colombia. CIAT-Universidad Nacional de Colombia, Cali.
29. FILGUEIRAS, T., G. DAVIDSE & F. ZULOAGA. 1993a. Ophiochloa, a new endemic serpentine grass genus (Poaceae: Paniceae) from the Brazilian Cerrado vegetation. Novon 3: 360-366.
30. FILGUEIRAS, T., O. MORRONE & F. ZULOAGA. 1993b. A new species of Streptostachys (Poaceae: Paniceae) from Brazil. Novon 3: 252-257.
31. FILGUEIRAS, T., G. DAVIDSE, F. ZULOAGA & O. MORRONE. 2001. The establishment of the new genus Altoparadisium and a reevaluation of Arthropogon (Poaceae, Paniceae). Ann. Missouri Bot. Gard. 88: 351-372.
32. FONT QUER, P. 1993. Diccionario de Botánica. Ed. Labor, S. A., Barcelona.
33. GARCÍA-ULLOA, J., C. LASTRA & C. SALAS. 2005. Estudios en gramíneas (Poaceae) de Colombia: veinte novedades corológicas. Caldasia 27 (2): 131-145.
34. GIRALDO-CAÑAS, D. 1998. Nuevas especies de Axonopus (Poaceae: Panicoideae: Paniceae) de la Amazonia colombiana. Caldasia 20 (2): 87-92.
35. GIRALDO-CAÑAS, D. 1999a. Nuevas citas de Axonopus (Poaceae: Panicoideae: Paniceae) para Colombia. Hickenia 3 (14): 47-50.
36. GIRALDO-CAÑAS, D. 1999b. Una nueva especie de Axonopus (Poaceae: Panicoideae: Paniceae) de la Guayana de Colombia y Venezuela. Caldasia 21 (2): 132-140.
37. GIRALDO-CAÑAS, D. 2000a. Nuevos registros de bambúes herbáceos (Poaceae) para la flora de Colombia. Hickenia 3 (16): 55-56.
38. GIRALDO-CAÑAS, D. 2000b. Estudios sobre la variación estructural de las sinflorescencias del género Axonopus (Poaceae: Panicoideae: Paniceae): tipología y tendencias evolutivas. Darwiniana 38: 208-219.
39. GIRALDO-CAÑAS, D. 2000c. Una nueva sección del género Axonopus (Poaceae, Panicoideae, Paniceae). Revista Acad. Colomb. Cienc. 24 (91): 183-191.
40. GIRALDO-CAÑAS, D. 2001a. Las especies de la tribu Paniceae (Poaceae: Panicoideae) de Colombia. Biota Colombiana 2 (3): 249-264.
41. GIRALDO-CAÑAS, D. 2001b. Sinopsis de la sección Cabrera del género neotropical Axonopus (Poaceae, Panicoideae, Paniceae). Revista Acad. Colomb. Cienc. 25 (95): 207-223.
42. GIRALDO-CAÑAS, D. 2001c. Estudios micromorfológicos y anatómicos en el género neotropical Axonopus (Poaceae: Paniceae). I. Epidermis foliar. Caldasia 23 (1): 119-133.
43. GIRALDO-CAÑAS, D. 2002a. Una nueva especie de Digitaria sect. Clavipilae (Poaceae: Panicoideae: Paniceae) de Colombia Caldasia 24 (2): 367-377.
44. GIRALDO-CAÑAS, D. 2002b. Una nueva especie de Glyceria (Poaceae: Pooideae: Meliceae) de Colombia. Caldasia 24 (1): 9-13.
45. GIRALDO-CAÑAS, D. 2002c. Revisión de las especies colombianas del género neotropical Axonopus (Poaceae, Panicoideae, Paniceae). Monografía en evaluación, Flora de Colombia.
46. GIRALDO-CAÑAS, D. 2002d. Estudios micromorfológicos y anatómicos en el género neotropical Axonopus (Poaceae: Paniceae). II. Antecio superior. Caldasia 24 (2): 293-316.
47. GIRALDO-CAÑAS, D. 2002e. Las especies de la sect. Lappagopsis del género neotropical Axonopus (Poaceae, Panicoideae, Paniceae). Revista Acad. Colomb. Cienc. 26 (98): 13-23.
48. GIRALDO-CAÑAS, D. 2003a. Novedades taxonómicas y corológicas en la tribu Paniceae (Poaceae: Panicoideae) para Colombia: Digitaria y Panicum. Caldasia 25 (2): 209-227.
49. GIRALDO-CAÑAS, D. 2003b. Revisión de las especies del género neotropical Axonopus P. Beauv. (Poaceae: Panicoideae: Paniceae), serie Barbigeri G. A. Black. Tesis doctoral, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Buenos Aires (Argentina).
50. GIRALDO-CAÑAS, D. 2004a. Características micromorfológicas y anatómicas de la espiguilla y el antecio superior del género Digitaria (Poaceae: Panicoideae: Paniceae). Caldasia 26 (1): 1-35.
51. GIRALDO-CAÑAS, D. 2004b. Novedades agrostológicas (Poaceae) para Colombia, México y Venezuela. Caldasia 26 (1): 315-322.
52. GIRALDO-CAÑAS, D. 2004c. El género Polypogon (Poaceae: Pooideae) en Colombia. Caldasia 26 (2): 417-422.
53. GIRALDO-CAÑAS, D. & C. MAYORGA. 2001. Nuevos registros de gramíneas (Poaceae) para la flora de Colombia. Hickenia 3 (28): 99-103.
54. GRAM, K. 1961. The inflorescence of the grasses. Bot. Tidsskr. 56: 293-313.
55. GUISSANI, L., J. COTA-SÁNCHEZ, F. ZULOAGA & E. KELLOGG. 2001. A molecular phylogeny of the grass subfamily Panicoideae (Poaceae) shows multiple origins of C4 photosynthesis. Amer. J. Bot. 88: 1993-2012.
56. HATTERSLEY, P. 1987. Variations in photosynthetic pathway. En: T. Soderstrom, K. Hilu, C. Campbell & M. Barkworth (eds.), Grass Systematics and Evolution: 49-64. Smithsonian Institution Press, Washington, D. C.
57. HITCHCOCK, A. 1935. Manual of the grasses of the United States. Misc. Publication 200: 1-1040. United States Department of Agriculture, Washington, D. C.
58. HSU, C. 1965. The classification of Panicum (Gramineae) and its allies, with special reference to the characters of lodicule, style-base and lemma. J. Fac. Sci. Univ. Tokyo 9: 43-150.
59. JOHNSTON, C. & L. WATSON. 1981. Germination flaps in grass lemmas. Phytomorphology 31: 78-85.
60. JUDZIEWICZ, E. 1990. Poaceae, fasc. 8, family 187. A. Gorts-van Rijn (ed.), Flora of the Guianas. Koeltz Scientific Books, Alemania.
61. KELLOGG, E. & C. CAMPBELL. 1987. Phylogenetic analyses of the Gramineae. En: T. Soderstrom, K. Hilu, C. Campbell & M. Barkworth (eds.), Grass systematics and evolution: 310-322. Smithsonian Institution Press, Washington D. C.
62. KILLEEN, T. 1990. The grasses of Chiquitanía, Santa Cruz, Bolivia. Ann. Missouri Bot. Gard. 77: 125-201.
63. KILLEEN, T. & Z. RÚGOLO DE AGRASAR. 1992. Taxonomy and reproductive biology of Digitaria dioica and D. neesiana (Gramineae: Paniceae). Syst. Bot. 17: 594-606.
64. KRISHNAN, S. 2000. Phytoliths of Indian grasses and their potential use in identification. Bot. J. Linnean Soc. 132: 241-252.
65. LUCES DE FEBRES, Z. 1963. Las gramíneas del Distrito Federal. Instituto Botánico, Dirección de Recursos Naturales Renovables, Ministerio de Agricultura y Cría, Caracas.
66. LUCES, Z. 1942. Géneros de las gramíneas venezolanas. Ministerio de Agricultura y Cría, Boletín Técnico no. 4: 1-149. Tipografía Garrido, Caracas.
67. MAYNE, B., G. EDWARDS & C. BLACK. 1971. Spectral, physical, and electron transport activities in the photosynthetic apparatus of mesophyll cells and bundle-sheath cells of Digitaria sanguinalis (L.) Scop. Pl. Physiol. 47: 600-605.
68. MCCLURE, F. & T. SODERSTROM. 1972. The agrostological term Anthecium. Taxon 21: 153-154.
69. MEJÍA-SAULÉS, M. 1986. Gramíneas forrajeras en la región central del Estado de Veracruz. Cuadernos de divulgación INIREB, Nro. 22. Instituto Nacional de Investigaciones sobre Recursos Bióticos, Xalapa.
70. METCALFE, C. 1960. Anatomy of monocotyledons. I. Gramineae. Clarendon Press, Oxford.
71. MORRONE, O. & F. ZULOAGA. 1989. Sobre la posición sistemática de Brachiaria glabrinodis (Poaceae: Paniceae). Darwiniana 29: 261-268.
72. MORRONE, O. & F. ZULOAGA. 1991a. Revisión del género Streptostachys (Poaceae-Panicoideae), su posición sistemática dentro de la tribu Paniceae. Ann. Missouri Bot. Gard. 78: 359-376.
73. MORRONE, O. & F. ZULOAGA. 1991b. Estudios morfológicos en el subgénero Dichanthelium de Panicum (Poaceae: Panicoideae: Paniceae), con especial referencia a Panicum sabulorum. Ann. Missouri Bot. Gard. 78: 915-927.
74. MORRONE, O. & F. ZULOAGA. 1992. Revisión de las especies sudamericanas de los géneros Brachiaria y Urochloa (Poaceae: Panocoideae: Paniceae). Darwiniana 31: 43-109.
75. MORRONE, O., T. FILGUEIRAS, F. ZULOAGA & J. DUBCOVSKY. 1993. Revision of Anthaenantiopsis (Poaceae: Panicoideae: Paniceae). Syst. Bot. 18: 434-453.
76. MORRONE, O., A. VEGA & F. ZULOAGA. 1996. Revisión de las especies del género Paspalum (Poaceae: Panicoideae: Paniceae), grupo Dissecta (s. str.). Candollea 51: 103-138.
77. MORRONE, O., F. ZULOAGA, M. ARRIAGA, R. POZNER & S. ALISCIONI. 1998. Revisión sistemática y análisis cladístico del género Chaetium (Poaceae: Panicoideae: Paniceae). Ann. Missouri Bot. Gard. 85: 404-424.
78. NICORA, E. & Z. RÚGOLO DE AGRASAR. 1987. Los géneros de gramíneas de América austral. Ed. Hemisferio Sur, Buenos Aires.
79. PALACIOS, R. 1968. Variaciones en la estructura de las espiguillas en algunas especies argentinas de Panicum. Bol. Soc. Argent. Bot. 12: 38-43.
80. PEÑALOZA-JIMÉNEZ, G., P. PETERSON & D. GIRALDO-CAÑAS. 2002. Los géneros Eragrostis y Leptochloa (Poaceae: Cynodonteae) en Colombia. Hickenia 3 (35): 133-141.
81. PINTO-ESCOBAR, P. 2002. Las gramíneas en Colombia. Revista Acad. Colomb. Cienc. 26 (98): 59-68.
82. PINTO-NOLLA, J. 1997. Notas sobre la tribu Paniceae R. Brown (Gramineae) y clave para sus géneros en Colombia. Revista Acad. Colomb. Cienc. 21 (80): 217-223.
83. POHL, R. 1980. Gramineae. En: W. Burguer (ed.), Flora Costaricensis. Fieldiana, Bot., New Series 4: 1-608.
84. POHL, R. & G. DAVIDSE. 1994. Digitaria. En: G. Davidse, S. Sousa & A. Chater (eds.), Flora Mesoamericana 6: 365-371.
85. POILECOT, P. 1999. Les Poaceae du Niger: description, illustration, ecologie, utilisations. Boissiera 56: 5-766.
86. RENVOIZE, S. 1984. The grasses of Bahia. Royal Bot. Gard., Kew.
87. RENVOIZE, S. 1998. Gramíneas de Bolivia. Royal Bot. Gard., Kew.
88. RENVOIZE, S. & F. ZULOAGA. 1984. The genus Panicum group Lorea (Gramineae). Kew Bull. 39: 185-202.
89. RODRÍGUEZ, H., E. CAMACHO DE TORRES & L. GUEVARA. 2000. La sección Parviflora Rodríguez del género Paspalum L. (Gramineae) en Venezuela. Ernstia 10 (3): 117-143.
90. ROSENGURTT, B., B. ARILLAGA DE MAFFEI & P. IZAGUIRRE DE ARTUCIO. 1970. Gramíneas uruguayas. Ed. Depto. de Publicaciones, Universidad de la República, Montevideo.
91. ROST, T. & D. SIMPER. 1975. The germination lid: A characteristic of the lemma in the Paniceae. Madroño 23: 68-72.
92. ROTH, I. 1977. Fruits of Angiosperms.Gebrüder Borntraeger, Berlín-Stuttgart.
93. RUA, G. 1999. Inflorescencias, bases teóricas para su análisis. Sociedad Argentina de Botánica, Buenos Aires.
94. RUA, G. 2003. Growth forms, branching patterns, and inflorescence structure in Digitaria sect. Trichachne (Poaceae, Paniceae). Flora 198: 178-187.
95. RUA, G. & I. BOCCALONI. 1996. The inflorescence of Digitaria phaeotrix: morphological and developmental aspects. Flora 191: 117-19.
96. RUA, G. & C. GRÓTTOLA. 1997. Growth form models within genus Paspalum L. (Poaceae: Paniceae). Flora 192: 65-80.
97. RUA, G. & F. WEBERLING. 1995. Growth form and inflorescence structure of Paspalum L. (Poaceae: Paniceae): a comparative morphological approach. Beitr. Biol. Pflanzen 69: 363-431.
98. RÚGOLO DE AGRASAR, Z. 1974. Las especies del género Digitaria (Gramineae) de la Argentina. Darwiniana 19: 65-166.
99. RÚGOLO DE AGRASAR, Z. 1994. Digitaria. En: R. Spichiger & L. Ramella (eds.), Flora del Paraguay 23 (Gramineae V): 75-138.
100. RÚGOLO DE AGRASAR, Z. & E. NICORA. 1993. Paspalum verrucosum y Eriochloa grandiflora (Gramineae, Paniceae), nuevas citas para la Argentina. Hickenia 2 (22): 99-103.
101. RÚGOLO DE AGRASAR, Z. & E. SÁNCHEZ. 1989. Estudios sobre el género Digitaria. I. Anatomía de Digitaria phaeothrix y especies afines. Bol. Soc. Argent. Bot. 26: 107-118.
102. SANGSTER, A. 1977. Characteristics of silica deposition in Digitaria sanguinalis (L.) Scop. (Crabgrass). Ann. Bot. 41: 341-350.
103. SHAW, R. & F. SMEINS. 1979. Epidermal characteristics of the callus in Eriochloa (Poaceae). Amer. J. Bot. 66: 907-913.
104. SHAW, R. & F. SMEINS. 1981. Some anatomical and morphological characteristics of North American species of Eriochloa (Poaceae: Paniceae). Bot. Gazette 142: 534-544.
105. SHAW, R. & R. WEBSTER. 1983. Characteristics of the upper anthecia of Ichnanthus (Poaceae: Paniceae). Bot. Gazette 144: 363-370.
106. SNOW, N. 1996. The phylogenetic utility of lemmatal micromorphology in Leptochloa s.l. and related genera in subtribe Eleusininae (Poaceae, Chloridoideae, Eragrostideae). Ann. Missouri Bot. Gard. 83: 504-529.
107. SRIVASTAVA, A. 1978. Study of the leaf epidermis in the genus Digitaria (Gramineae). J. Indian Bot. Soc. 57: 155-160.
108. SUBILS, R. 1995. Digitaria. En: F. Zuloaga (ed.), Flora Fanerogámica Argentina 12: 21-36.
109. THOMPSON, R. & J. ESTES. 1986. Anthecial and foliar micromorphology and foliar anatomy of Brachiaria. Amer. J. Bot. 73: 398-408.
110. TOVAR, O. 1993. Las gramíneas (Poaceae) del Perú. Ruizia 13: 1-480.
111. TÜRPE, A. 1966. Histotaxonomía de las especies argentinas del género Paspalum. Lilloa 32: 1-272.
112. VEGA, A. 1996. Biología reproductiva de Panicum spathelosum (Poaceae: Panicoideae: Paniceae). Darwiniana 34: 199-211.
113. VEGA, A. & Z. RÚGOLO DE AGRASAR. 2001. Morphological interpretation of the spikelet in Digitaria atra (Poaceae: Panicoideae: Paniceae) and emended generic description. Amer. J. Bot. 88: 1670-1674.
114. VEGA, A. & Z. RÚGOLO DE AGRASAR. 2002a. Novedades taxonómicas y sinopsis del género Digitaria (Poaceae: Panicoideae: Paniceae) en Bolivia. Darwiniana 40: 171-190.
115. VEGA, A. & Z. RÚGOLO DE AGRASAR. 2002b. Digitaria killeenii (Poaceae: Panicoideae: Paniceae), a new species from Bolivia. Syst. Bot. 27: 252-256.
116. VEGETTI, A. & A. ANTON. 1995. Some evolution trends in the inflorescence of Poaceae. Flora 190: 225-228.
117. VEGETTI, A. & A. ANTON. 1996. The synflorescence concept in Poaceae. Flora 191: 231-234.
118. VEGETTI, A. & J. PENSIERO. 1993. Tipología de la inflorescencia de Panicum sección Panicum (Poaceae: Paniceae). Bol. Soc. Argent. Bot. 29: 7-10.
119. VEGETTI, A. & F. WEBERLING. 1996. The structure of the paracladial zone in Poaceae. Taxon 45: 453-460.
120. VELDKAMP, J. 1973. A revision of Digitaria Haller (Gramineae) in Malesia. Blumea 21: 1-80.
121. WATSON, L. & M. DALLWITZ. 1992. The grass genera of the world. CAB International, Wallingford.
122. WEBSTER, R. 1983. A revision of the genus Digitaria Haller (Paniceae: Poaceae) in Australia. Brunonia 6: 131-216.
123. WEBSTER, R. 1987. Taxonomy of Digitaria section Digitaria in North America (Poaceae: Paniceae). Sida 12: 209-222.
124. WEBSTER, R. 1988. Genera of the North American Paniceae (Poaceae: Panicoideae). Syst. Bot. 13: 576-609.
125. WEBSTER, R. 1992. Character significance and generic similarities in the Paniceae (Poaceae: Panicoideae). Sida 15: 185-213.
126. WEBSTER, R. & S. HATCH. 1981. Two Digitaria species (Poaceae) new to the New World. Sida 9: 87-88.
127. WEBSTER, R. & S. HATCH. 1990. Taxonomy of Digitaria section Aequiglumae (Poaceae: Paniceae). Sida 14: 145-167.
128. WIPFF, J. & S. HATCH. 1994. A systematic study of Digitaria sect. Pennatae (Poaceae: Paniceae) in the New World. Syst. Bot. 19: 613-627.
129. ZUCOL, A. 1996. Microfitolitos de las Poaceae argentinas: I. Microfitolitos foliares de algunas especies del género Stipa (Stipeae: Arundinoideae) de la provincia de Entre Ríos. Darwiniana 34: 151-172.
130. ZUCOL, A. 1998. Microfitolitos de las Poaceae argentinas: II. Microfitolitos foliares de algunas especies del género Panicum (Poaceae, Paniceae) de la provincia de Entre Ríos. Darwiniana 36: 29-50.
131. ZUCOL, A. 2000. Fitolitos de Poaceae de Argentina. III. Fitolitos foliares de especies del género Paspalum (Paniceae) en la provincia de Entre Ríos. Darwiniana 38: 11-32.
132. ZULOAGA, F. 1985. El género Panicum (L.) sección Parviglumia. Darwiniana 26: 353-369.
133. ZULOAGA, F. 1987a. Systematics of New World species of Panicum (Poaceae: Paniceae). En: T. Soderstrom, K. Hilu, C. Campbell & M. Barkworth (eds.), Grass systematics and evolution: 287-306. Smithsonian Institution Press, Washington D. C.
134. ZULOAGA, F. 1987b. A revision of Panicum subgenus Panicum section Rudgeana (Poaceae: Paniceae). Ann. Missouri Bot. Gard. 74: 463-478.
135. ZULOAGA, F., R. ELLIS & O. MORRONE. 1992. A revision of Panicum subgenus Phanopyrum sect. Laxa (Poaceae: Panicoideae: Paniceae). Ann. Missouri Bot. Gard. 79: 770-818.
136. ZULOAGA, F. & O. MORRONE. 1992. Panicum mystasipum (Poaceae: Panicoideae: Paniceae), una nueva especie para el Brasil. Iheringia, Sér. Bot. 42: 13-23.
137. ZULOAGA, F. & O. MORRONE. 1994. Mesosetum. En: R. Spichiger & L. Ramella (eds.), Flora del Paraguay 23 (Gramineae V): 200-202.
138. ZULOAGA, F. & O. MORRONE. 1996. Revisión de las especies americanas de Panicum sección Panicum (Poaceae: Panicoideae: Paniceae). Ann. Missouri Bot. Gard. 83: 200-280.
139. ZULOAGA, F., O. MORRONE, G. DAVIDSE, T. FILGUEIRAS, P. PETERSON, R. SORENG & E. JUDZIEWICZ. 2003. Catalogue of New World grasses (Poaceae): III. Subfamilies Panicoideae, Aristoideae, Arundinoideae, and Danthonioideae. Contr. U.S. Natl. Herb. 46: 1-662.
140. ZULOAGA, F., O. MORRONE & J. DUBCOVSKY. 1989. Exomorphological, anatomical, and cytological studies in Panicum validum (Poaceae: Panicoideae: Paniceae): Its systematic position within the genus. Syst. Bot. 14: 220-230.
141. ZULOAGA, F., O. MORRONE & T. KILLEEN. 1993. Gerritea, a new genus of Paniceae (Poaceae: Panicoideae) from South America. Novon 3: 213-219.
142. ZULOAGA, F., O. MORRONE, Z. RÚGOLO DE AGRASAR, A. ANTON, M. ARRIAGA & A. CIALDELLA. 1994. Gramineae V. En: R. Spichiger & L. Ramella (eds.), Flora del Paraguay 23: 1-327.
143. ZULOAGA, F., O. MORRONE & A. SÁENZ. 1987. Estudio exomorfológico e histofoliar de las especies americanas del género Acroceras (Poaceae: Paniceae). Darwiniana 28: 191-217.
144. ZULOAGA, F., O. MORRONE, A. VEGA & L. GIUSSANI. 1998. Revisión y análisis cladístico de Steinchisma (Poaceae: Panicoideae: Paniceae). Ann. Missouri Bot. Gard. 85: 631-656.
145. ZULOAGA, F., A. SÁENZ & O. MORRONE. 1986. El género Panicum (Poaceae: Paniceae) sect. Cordovensia. Darwiniana 27: 403-429.
146. ZULOAGA, F. & T. SENDULSKY. 1988. A revision of Panicum subgenus Phanopyrum section Stolonifera (Poaceae: Paniceae). Ann. Missouri Bot. Gard. 75: 420-455.
147. ZULOAGA, F. & T. SODERSTROM. 1985. Classification of the outline species of New World Panicum (Poaceae: Paniceae). Smithsonian Contr. Bot. 59: 1-63.
Recibido: 03/11/2003
Aceptado: 01/05/2004

