










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.28 no.1 Bogotá June 2006
OSTEOLOGÍA DE ELEUTHERODACTYLUS BOGOTENSIS (AMPHIBIA, ANURA, LEPTODACTYLIDAE) DEL PARQUE NACIONAL NATURAL CHINGAZA (CUNDINAMARCA, COLOMBIA)
Osteology of Eleutherodactylus bogotensis (Amphibia, Anura, Leptodactylidae) from the Parque Nacional Natural Chingaza (Cundinamarca, Colombia)
PAOLA AVILÁN
JULIO MARIO HOYOS
Pontificia Universidad Javeriana, Departamento de Biología, Unidad de Ecología y Sistemática (UNESIS), A.A. 56710, Bogotá, Colombia, jmhoyos@javeriana.edu.co
RESUMEN
Se presenta la descripción de las estructuras óseas de la rana Eleutherodactylus bogotensis (Leptodactylidae, Anura). Para este fin se estudiaron cuatro individuos (dos hembras y dos machos) colectados en el Parque Nacional Natural Chingaza (Cundinamarca, Colombia), en el Sector de Piedras Gordas a 3250 m.s.n.m. Se confirmó la pertenencia al grupo unistrigatus (sensu Lynch) con base en la osteología craneal. Se encontró variación en la disposición de algunas estructuras óseas a nivel del cráneo, variación interespecífica en el pubis y el esfenetmoide y la presencia de elementos sesamoideos. Se discuten las diferencias que se encontraron entre las estructuras de esta especie y lo encontrado por otros autores en otras especies de anuros.
Palabras clave. Amphibia, Anura, Eleutherodactylus bogotensis, osteología, Parque Nacional Natural Chingaza.
ABSTRACT
Osteology description of Eleutherodactylus bogotensis (Leptodactylidae, Anura) is presented. Four speciemens (two males and two females) collected at the Parque Nacional Natural Chingaza (Cundinamarca, Colombia), Piedras Gordas area, were examined. It was confirmed that this species belongs to the unistrigatus group (sensu Lynch,) base upon the skull. Several osteological variations at the level of the skull, the pubis, and presence of sesamoidal elements were discovered. Differences between osteological characters in E. bogotensis and several anuran species described by other authors are discussed.
Key words. Amphibia, Anura, Eleutherodactylus bogotensis, osteology, Chingaza Natural National Park.
INTRODUCCIÓN
La familia Leptodactylidae es la más diversa dentro de los anuros con 52 géneros y más de 800 especies agrupadas en cuatro subfamilias (Zug et al. 2001), siendo Telmatobiinae a la cual pertenece el género Eleutherodactylus, distribuida en su mayoría en Suramérica, el sur de Norteamérica y el Caribe, además de estar en Chile, Brasil, Argentina y México y dos especies de Eleutherodactylus introducidos en la Florida (EE.UU.) (Duellman & Trueb 1994).
El género Eleutherodactylus es el más grande y más homogéneo dentro de los vertebrados, distribuyéndose principalmente en toda Colombia y en el mundo en Sinaloa y Tamaulipas, México; en el sur y este de América Central hasta el norte de Argentina y sur de Brasil, el oeste de la India y dos especies introducidas en la Florida (Lynch 1971). Dentro de estos, el grupo unistrigatus contiene aproximadamente100 especies distribuidas principalmente en Sur América, dentro de las cuales está ubicada Eleutherodactylus bogotensis Peters, 1863 (Lynch & Duellman 1997).
En Colombia, E.bogotensis se distribuye principalmente en el área de Bogotá y sus alrededores, entre los 2600 y 3400 m.s.n.m. Los ejemplares estudiados fueron colectados en el Parque Nacional Natural Chingaza (Cundinamarca, Colombia) en el sector de Piedras Gordas (3250 m.s.n.m.), donde se puede encontrar en ecosistemas de páramo y subpáramo (Hoyos 1991).
Dentro de la descripción y la diagnosis de esta especie se ha tenido siempre en cuenta tan sólo caracteres externos del cuerpo, miembros, rostro, patrón de coloración, diferentes medidas del cuerpo, forma, entre otros aspectos, sin considerar características de huesos y músculos (Hoyos 1991). De ahí que nuestro objetivo sea el hacer una descripción de la mayor parte de los huesos de E. bogotensis. Para lograr esto se estudiaron algunos de los ejemplares pertenecientes a la colección de la Pontificia Universidad Javeriana. Estos individuos fueron tratados básicamente con la utilización del método de transparentación enzimático y coloración diferencial para la observación y descripción de los huesos.
El estudio detallado de la osteología de los anuros y en este caso de E.bogotensis tiene gran importancia ya que las características que se presentan a este nivel van a contribuir al conocimiento de la especie; además que con los resultados obtenidos de este trabajo se pueden hacer comparaciones y utilizar algunos de estos caracteres anatómicos en la reconstrucción de filogenias y poder aplicar el uso de estas características en otros grupos de anuros. También es importante este estudio para tener mejor conocimiento de los aspectos funcionales inherentes a dichas estructuras. Debido a la importancia de las comparaciones con otros trabajos, consideramos fundamental elaborar una síntesis de los estudios más relevantes acerca de la osteología de los anuros.
El primero y más importante trabajo que se ha hecho acerca de la anatomía de anuros corresponde al realizado por Ecker (1971) donde estudió la anatomía de Rana esculenta utilizando nombres derivados de estudios de anatomía humana aplicado a los anuros, y convirtiéndose en el pionero y más amplio a nivel de toda la osteología de los anuros. Aún hoy, algunos autores se fundamentan en este trabajo para sus estudios, como es el caso de Duellman & Trueb (1994) donde presentan una recopilación de las generalidades de los anfibios anuros, en cuanto a su sistema esquelético-muscular.
Ritland (1955) hizo un estudio de la morfología poscraneal de Ascaphus truei; Trueb (1973) realizó uno de los trabajos más significativos acerca de los huesos de las ranas y su evolución, describiendo las generalidades de las características óseas y especificando las particularidades de algunas familias y géneros, determinando estados, patrones y tendencias evolutivas de varios huesos. Clarke (1981) estudió las relaciones en ránidos africanos, basado en caracteres osteológicos, presentando diferentes estados de carácter y Clarke (1987) hizo la descripción de la morfología esquelética de Barbourula (Discoglossidae).
Laurent & Fabrezi (1989-1990), estudiaron las estructuras carpianas en ránidos describiendo diferencias en estructuras. De Sá & Trueb (1991) investigaron la osteología de Hamptophryne boliviana (Microhylidae), y describieron forma y características propias de cada hueso y los estados de desarrollo de los mismos. Fabrezi (1992) hizo una comparación de los carpos de los anuros entre diferentes familias exponiendo diferentes morfologías y ontogenias entre ellas; Fabrezi (1993) comparó el tarso de los anuros, también entre diferentes familias corroborando los estudios anteriores. Fabrezi & Alberch (1996) estudiaron los elementos carpianos de los anuros entre muchas familias. Recientemente, Sheil (1999) realizó un estudio sobre la osteología y el desarrollo del esqueleto de Pyxicephalus adspersus (Ranidae).
Acerca de la familia Leptodactylidae y especialmente del género Eleutherodactylus, se han realizado varios trabajos basados generalmente en osteología, aunque también hay algunos aportes acerca de la miología. Davies & Burton (1982) describieron características básicas de la osteología y miología de Rheobatrachus silus y Wild (1997) hizo una completa descripción del esqueleto adulto y desarrollo osteológico de Ceratophrys cornuta. En cuanto a estudios del género, es principalmente Lynch (1971, 1976, 1978, 1993) quien viene trabajando desde hace ya varios años en este grupo: en 1971 hizo un estudio sobre las relaciones de leptodactílidos teniendo en cuenta sus características óseas; en 1976, dividió el género Eleutherodactylus en diez grupos de especies distribuidas en Sur América, diagnosticando cada uno con caracteres osteológicos craneales; en 1978 realizó un estudio de leptodactílidos de la Patagonia haciendo un detallado estudio descriptivo de las estructuras óseas de todo el cuerpo y las diferencias entre las especies de esta área y en 1993 realizó hipótesis filogeneticas de Eleutherodactylus basados en el músculo depressor mandibulae.
Más adelante, Lynch & Ruiz-Carranza (1985) realizaron una sinopsis del género Eleutherodactylus de la Sierra Nevada de Santa Marta donde se describe la musculatura mandibular y la osteología craneal y con base en esto y algunos caracteres externos, plantean y discuten algunas relaciones con otras especies del género.
De nuevo Lynch (1986) estudió las relaciones de Eleutherodactylus basado en la musculatura mandibular y en 1989 estudió las relaciones especies del género de tierras medias, bajo este mismo fundamento. En el año 1990 describe una nueva especie en Colombia incluyendo características de su osteología craneal y en 1996 estudia las relaciones del subgénero Pelorius (Eleutherodactylus) basado en osteología craneal.
El único estudio encontrado de otros autores exponiendo algunas características osteológicas y miológicas, es el realizado por Rivero & Serna (1987) en la descripción de tres nuevas especies de Eleutherodactylus.
MATERIALES Y MÉTODOS
Se disecaron cuatro especímenes adultos (dos hembras y dos machos) colectados en el Sector Piedras Gordas, en el Parque Nacional Natural Chingaza, pertenecientes a la colección del Museo de la Pontificia Universidad Javeriana, Bogotá, Colombia, con los números de colección MUJ 478, MUJ 466 (hembras) y MUJ 093, MUJ 062 (machos). Este número de especímenes es suficiente para establecer el dimorfismo sexual y la variabilidad intraespecífica tal como ha sido demostrado en estudios miológicos por Raikow et al. (1990) y osteológicos por Hoyos (1999).
La disección se llevó a cabo por medio de la utilización de un estereoscopio con 16X y 40X de magnificación. Las comparaciones se hicieron con base en descripciones de especies de diferentes familias y géneros. Para la visualización del esqueleto se utilizó el procedimiento de tinción diferencial propuesto por Dingerkus & Uhler (1977). Los huesos se dibujaron con ayuda de una cámara clara por el plano donde mejor se observaron las características; en los huesos de la cabeza y la columna se dibujaron los huesos dorsales y ventrales y en la cabeza, también laterales.
La nomenclatura utilizada está dividida por partes: para la osteología apendicular se basó en las descripciones de Fabrezi (1992 y 1993) y los demás elementos según Duellman & Trueb (1994) y Hoyos (1999). Los nombres latinos de los huesos siguen la nomenclatura propuesta por el Consejo Internacional de Nomenclatura Anatómica Veterinaria (Anónimo 1973).
La discusión se hizo con base en la comparación con especies pertenecientes y no a la familia Leptodactylidae, teniendo en cuenta la amplitud y precisión de las descripciones que se han hecho para especies de otros grupos, por ejemplo Ranidae e Hylidae. Debido a que no son muchas las monografías osteológicas por especie, se tuvo en cuenta sobre todo aquellas en que la descripción era el centro del trabajo con el que se compara y no las breves anotaciones que se hacen en las descripciones de nuevas especies.
RESULTADOS
Skeleton axiale
Ossa cranii (Fig. 1 (A, B y C) y Fig. 2 (A y B))
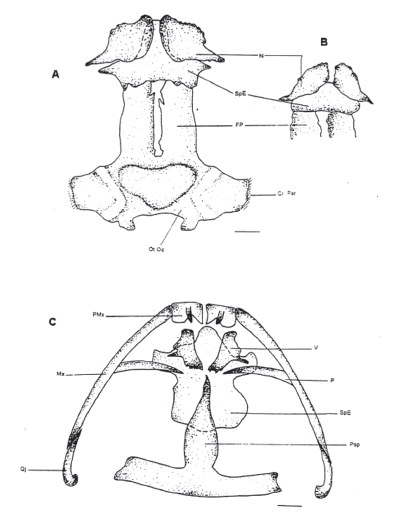
Figura 1. Vista dorsal del cráneo. A. Patrón general. MUJ 478. B. Variación en el esfenetmoide. MUJ 093. Escala 1 mm. C Vista ventral del cráneo. MUJ 478. Escala 1 mm. Ver convenciones en la tabla 1.
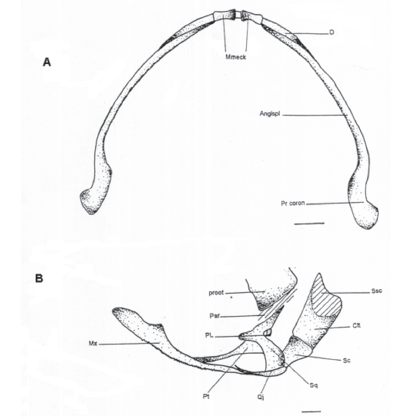
Figura 2. A. Vista ventral de la mandíbula. MUJ 478. Escala: 1 mm. B. Vista lateral del cráneo. MUJ 478. Escala: 1 mm
Os frontoparietale (FP)
Hueso par, ancho y largo de bordes internos irregulares separados uno del otro por un espacio relativamente grande. Anteriormente recubre parcialmente la parte posterior del esfenetmoide y posteriormente recubre el proótico y limita con el exoccipital. En el ejemplar MUJ 466 (hembra) se superpone parcialmente lateral y posteriormente con el proótico.
Tabla 1. Abreviaturas utilizadas

Os nasale (N)
Hueso par, posterior a la premaxila que recubre la cápsula nasal, separados entre si en la línea media. Se articula hacia cada lado de la cápsula nasal con los cartílagos de la misma y a los huesos palatinos por medio de los procesos posterolaterales. Posteriormente cubre al esfenetmoide cubriendo su parte anterior; hay variabilidad en el grado de recubrimiento.
Os otoccipitale (OtOc)
Es la fusión de dos huesos, el proótico y el exoccipital. El proótico posee dorsalmente la cresta parótica (crista parotica) cartilaginosa, sobre la cual se articula la rama otica del escamoso. El exoccipital se ubica posterior, del cual salen los cóndilos occipitales, uno a cada lado para articular el cráneo con la primera vértebra. En el ejemplar MUJ 478 los exoccipitales no están separados y el proótico no está totalmente fusionado al exoccipital.
Os sphenethmoidale (SpE)
Formado por una pequeña parte visible dorsalmente que se articula anteriormente con los nasales y posterolateralmente al frontoparietal de cada lado, de donde salen unas láminas verticales (laterales) que llegan a la parte ventral del cráneo agrandando su longitud para articularse posteriormente con la tercera parte del proceso cultriforme del paraesfenoides, hasta el extremo del mismo. Lateralmente tiene un proceso muy corto que se articula a los palatinos y anteriormente a los vómeres. En su extremo más anterior está el septum nasi. El proceso anterior es largo en la mayoría de los casos, excepto en el ejemplar MUJ 466 en el cual es muy corto, casi aplanado.
Ventrales
Premaxilla (PMx)
Hueso par separado uno del otro en el medio. Son los elementos más anteriores de la región maxilar y comprende tres partes: la pars dentalis, en la cual están los dientes; el processus alaris que está inclinado en posición anteroposterior en un ángulo menor a 90º, y la pars palatina en donde están los processus palatina que se presentan dispuestos horizontalmente. El proceso lateral es romo distalmente y el medial es puntiagudo, sin tocarse entre ellos.
Maxilla (Mx)
Elemento par que posee dientes. Comienza articulando por contacto anteriormente a la pars palatina de la PMx. Posee tres partes: la pars facialis que comprende la mitad anterior de la misma más gruesa que la parte posterior, dentro de la cual está el processus orbitalis ubicado anterodorsalmente, más corto con respecto a la pars facialis; se articula con el palatino formando la parte dorsal anterior de la orbita ocular sin articularse con los procesos posterolaterales de los nasales, y la pars palatina que es la parte ventral de la maxila. Distalmente se articula por superposición al cuadradoyugal.
Os quadratojugale (Qj)
Se origina inmediatamente posterior a la maxila, articulándose a esta por superposición; en su parte posterior se engrosa hasta llegar a articularse sobre la rama ventral del escamoso en su extremo distal.
Vomer (V)
Elementos pares ubicados en la parte anterior ventral del cráneo. Están formados por una placa palatal pequeña de donde salen dos procesos: el postcoanal o medial, casi tan largo como el diámetro de la coana y muy delgado que se encuentra libre y el precoanal que es el más anterior que se articula con los nasales. Los odontóforos no existen como tales, sin embargo hay un pequeño extremo terminal irregular de forma aserrada recubriendo parcialmente al esfenetmoide.
Os parasphenoidale (PSp)
Hueso ventral con forma de “T” invertida que recubre parcialmente al esfenetmoide. Está formado por dos procesos: su porción anterior es el processus cultriforme que se articula con el esfenetmoide en su extremo anterior el cual es agudo, los bordes laterales son biconvexos y su parte media no alcanza a llegar hasta el foramen magnum. Posterolateralmente, están los processus alaris de forma rectangular irregular que llegan hasta el extremo lateral del proótico recubriendo las cápsulas auditivas.
Os palatinum (P)
Elementos pares que no se juntan entre sí, ubicados lateralmente con respecto al solum nasi. Se originan proximalmente sobre los procesos laterales del esfenetmoide a nivel posterolateral del vómer formando el margen anteroventral de la órbita; se articulan sobre la pars facialis de la maxila y se extienden lateralmente sobre el planum antorbitale. Entre los extremos proximales de los dos palatinos se encuentra ubicado el proceso cultriforme del paraesfenoide.
Os mandibulae (M)
Compuesto por tres pares de elementos: el mentomekeliano que forma su extremo anterior articulándose entre sí por una conexión sindesmótica (cartilaginosa) y lateralmente se articulan al dentario; tienen forma bicóncava. El dentario que se articula anteriormente con el mentomekeliano, ocupa la mitad externa de la mandíbula articulándose en su porción media posterior con el angulosplenio; este último es el más largo y ocupa la mitad interna de la misma, teniendo un proceso coronoides en su parte posterior, y articulándose al cuadradoyugal. Estos tres elementos se encuentran recubriendo el cartílago de Meckel que recorre toda la mandíbula desde el mentomekeliano hasta el cuadradoyugal.
Laterales
Apparatus plectrum (PL)
El plectrum (o columnela) es el elemento que forma el oído medio de los anuros. Está compuesto por dos partes: la proximal (partes media e interna plectri) ósea y más ensanchada que se articula a la fenestra ovalis por una placa basal, y la parte distal (pars externa plectri) que se articula al tímpano y es más delgada terminando en un proceso cartilaginoso.
Os pterygoideum (Pt)
Compuesto por tres ramas que forman una “Y”. La rama anterior es la más larga que se extiende sobre el proceso pterigoideo (cartilaginoso) y se inserta sobre la maxila; la rama media se articula a la cápsula auditiva y la rama posterior se articula sobre el cuadradoyugal.
Os squamosus (Sq)
Consta de tres ramas: la rama ótica o posterior que se articula al proótico ventralmente y al otoccipital dorsalmente; la rama ventral, que es la más larga y la única vertical o perpendicular que se articula al cuadradoyugal, y la rama zigomática o anterior que es la más corta de las tres y se encuentra libre anteriormente.
Aparatus hyolaringeus (Fig. 3)
Comprende: la placa hioidea cartilaginosa, medial y pequeña de la cual se derivan todos los procesos hioideos. No tiene membrana hioglosa. El hial o cuerno que sale de la placa hioidea hacia arriba y se extiende hasta llegar a articularse sobre la parte posterior de la cápsula ótica. Los procesos posterolaterales que salen de la placa hioidea, son cortos, de forma triangular. De allí salen los procesos posteromediales (tirohiales) hacia cada lado sin estar fusionados. Los procesos anteriores del hial, son prolongaciones a manera de cortos filamentos cartilaginosos. Los tirohiales o procesos posteromediales son los únicos óseos del aparato hioideo. Se originan justo debajo de los procesos posterolaterales sin fusionarse a ellos; son largos, gruesos y sus extremos son cartilaginosos pasando dorsal a la cintura pectoral.
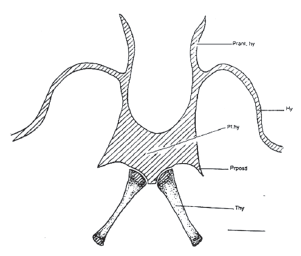
Figura 3. Aparato hioideo. MUJ 062. Escala: 1 mm
La laringe está compuesta por un par de cartílagos semicirculares llamados aritenoides y un anillo completo llamado cartílago cricoide. Posterolateralmente tiene un par de procesos bronquiales largos y delgados. Todo es cartilaginoso.
Skeleton thoracis
Omosternum (Omst) (Fig. 4A)
Único elemento prezonal de la cintura pectoral, es pequeño y cartilaginoso de forma alargada triangular con su base articulada al puente precoracoides.
Sternum (St) (Fig. 4A)
Elemento postzonal cartilaginoso con forma de puente cóncavo relativamente pequeño.
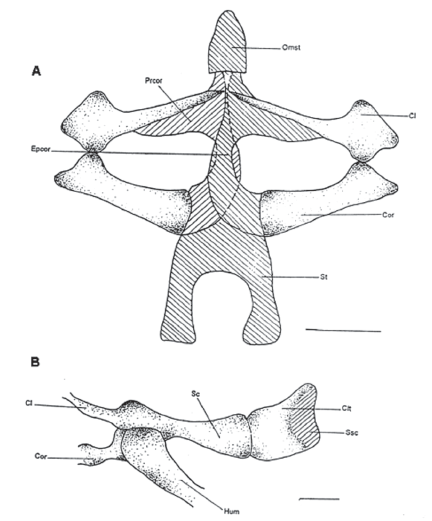
Figura 4. A. Vista ventral de la cintura pectoral. MUJ 478. B. Vista lateral de la cintura pectoral. MUJ 478. Escala: 1 mm
Columna vertebralis (Figs. 5)
Presenta ocho vértebras presacras. La primera vértebra presacra se articula al cráneo por medio de los cóndilos occipitales por una articulación de tipo I. Las siguientes siete vértebras son procélicas (igual que la primera). Estas siete vértebras tienen procesos transversos de diferentes formas y tamaños que terminan en cartílago; el proceso III es el más largo y ancho, los procesos V–VIII son delgados y más cortos. El individuo MUJ 062 sólo tiene siete vértebras presacras (hay dos fusionadas) y hay un proceso más en el lado izquierdo. Cada proceso tiene una inclinación: los procesos II y VIII están dirigidos posteroanteriormente, los procesos III y VII son horizontales y los procesos IV, V y VI están inclinados anteroposteriormente. Los cuerpos vertebrales están muy separados entres sí.

Figura 5. Columna vertebral. Izquierda: vista ventral Derecha: vista dorsal. MUJ 478. Escala: 1 mm
La diapófisis de la vértebra sacra (diapófisis sacra) es cilíndrica, más larga y ancha que las presacras, hacia cada lado distalmente se articula al ilium y a este nivel están los elementos sesamoideos que son cartilaginosos. La vértebra sacra se articula posteriormente de forma bicondilar al urostilo; hacia su parte más anterior tiene una cresta que va hasta su parte media y llega posteriormente hasta el ischium.
Skeleton appendiculare
Cingulum membri thoracici
Os coracoideus (Cor), procoracoideus (Prcor) y epicoracoideus (Epcor) (Fig. 4A)
Forman el área zonal de la cintura pectoral. El coracoideus que está separado simétricamente por el epicoracoides que en algunos casos alcanza a ser óseo pero en otros es cartilaginoso. El coracoides en su parte distal se articula con la escápula formando la fosa glenoidea. Los epicoracoides en E.bogotensis están superpuestos medialmente (cintura arciferal), posterior a la fusión del procoracoides con el extremo anteromedial del arco pectoral. Los procoracoideus están fusionados lateralmente y cubiertos anteriormente por la clavícula.
Clavicula (Cl) (Fig.4A)
Es una estructura par que se articula lateralmente con la escápula, posterolateralmente con el coracoides y cubre en su parte posterior al procoracoides. Tiene forma arqueada medialmente, delgada en su parte proximal y engrosada y con dos cabezas distalmente.
Scapula (Sc) (Fig.4A)
Componente par anterolateral de la cintura pectoral. Se articula a la clavícula y al coracoides formando la fosa glenoidea
Suprascapula (SSc) (Fig. 4B)
Se ubica dorsolateralmente en la cintura pectoral articulada a la escápula; tiene una parte cartilaginosa que está ubicada dorsalmente y otra ósea lateralmente que recibe el nombre de cleithrum. Dorsalmente la supraescápula está libre sosteniendo la musculatura dorsal.
Skeleton brachii (Figs. 6A)
Humerus (Hum)
El humerus es el elemento proximal (o segmento propodial), del miembro anterior; este posee dos crestas: la cresta ventral o deltoides que está bastante bien desarrollada y la cresta lateral a lado y lado del cóndilo distal; no hay desarrollo de la cresta medial. Este hueso es igual en hembras y machos.
Skeleton antibrachii
Radioulna (RU) (fig. 6A)
El radioulna forma el zeugopodio (o el segmento epipodial); su nombre se deriva de la fusión del radio con la ulna. Tiene un olecranón y no tiene crestas; en su parte anterior presenta un canal de división entre el radio y la ulna que se ve tanto ventral como dorsalmente.
Skeleton manus (Fig. 6B)
Ossa carpi (d2-5), Ossa metacarpalia (Mtcp), Ossa digitorum manus
Tiene seis elementos individuales carpales: ulnare, radiale, elemento “y”, carpiano distal 2, carpiano distal 3-4-5, elemento proximal del prepollex de dos elementos. El elemento más distal del prepollex es alargado y en forma de triángulo. La fórmula falangeal es 2-2-3-3 y la última falange está engrosada en la base y la terminación está modificada en forma de “T”. Todas las falanges tienen cartílagos en sus extremos.
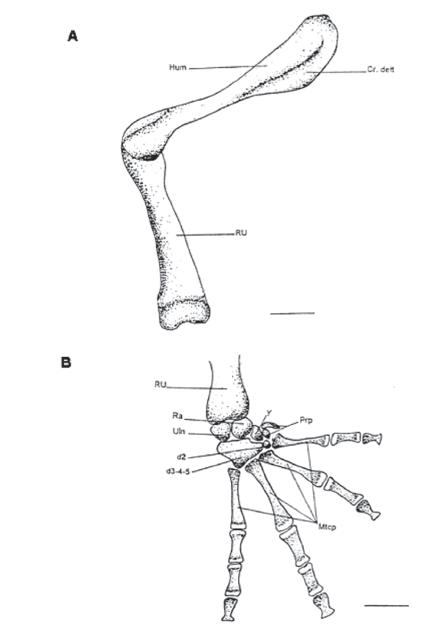
Figura 6. Vista ventral del miembro anterior. A. Húmero y radioulna. B. Mano. MUJ 478. Escala: 1 mm
Ossa memebri pelvini
Cingulum membri pelvini (Figs. 7A, B, C)
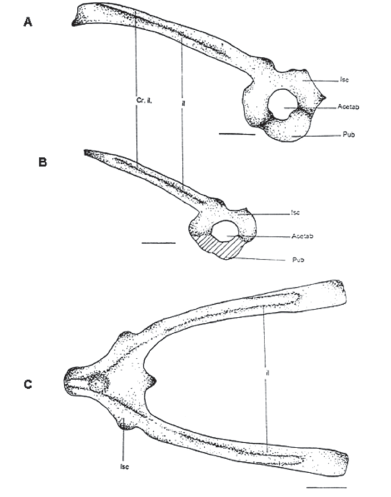
Figura 7. A. Pubis óseo. MUJ 478. B) Patrón general del pubis MUJ 466. C) Vista dorsal de la cintura pélvica. MUJ 478. Escalas: 1 mm
Os coxale
Consta de tres elementos unidos por una sínfisis media: el ilium, el ischium y el pubis.
Os ilium (il)
Elemento par. Es la parte más larga de la cintura pélvica y consta de una cresta ilial en su parte dorsal en donde tiene una protuberancia llamada protuberancia dorsal y una asta ilial ventral a ésta. La parte anterior está articulada a la diapófisis sacra en su parte ventral lateral a nivel de los elementos sesamoideos. El extremo posterior es más ensanchado en donde se articulan el pubis y el ischium para formar la parte anterior del acetábulo.
Os ischium (Isc)
Forma la parte posterior del acetábulo, es la parte más sobresaliente de la cintura pélvica. Dorsalmente tiene un pequeño proceso.
Os pubis (Pub)
Es la parte anteroventral que forma el acetábulo, en algunos casos es óseo (MUJ 478) pero en general es cartilaginoso.
Skeleton femoris
Os femoris (Fig. 8A)
Elemento proximal (o estilopodium) del miembro posterior, flexionado anteroposteriormente. Tiene un hueso sesamoideo detrás de la articulación con el tibiofíbula llamado fabella mediale (Fig 8A).
Skeleton cruris (Fig. 8A)
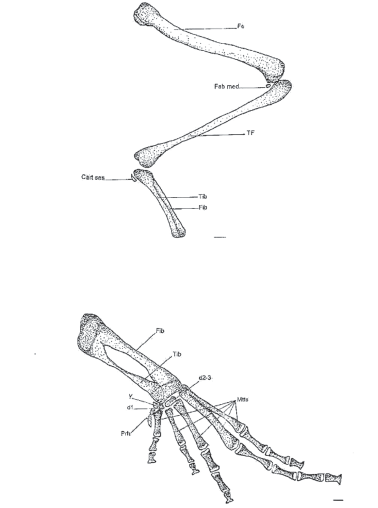
Figura 8. Vista ventral del miembro posterior. A. Muslo, pierna, tarsianos y elementos sesamoideos. B. Pie. MUJ 478. Escala: 1 mm
Tibiofibula (TF)
El tibiofíbula es el elemento medial (o zeugopodium) del miembro posterior. Está formado por la fusión de la tibia y la fibula. A nivel distal tiene un elemento sesamoideo detrás de la articulación con el tibial llamado cartílago sesamoideo.
Skeleton pedis (Fig. 8B)
Ossa tarsi (d1-5), Ossa metatarsalia (Mtts) Ossa digitorum pedis y Ossa sesamoidea (Ses)
Tiene cinco dedos. Su fórmula falangeal es 2-2-3-4-3. La última falange tiene forma de “T” igual que en las manos. Los metatarsianos IV y V están directamente articulados con el fibulare. Presenta tarsiano distal 2-3 (d2-3) fusionados que se asocian a los metatarsianos II y III. El tarsiano distal 1 (d1) se encuentra entre los metatarsianos I y II, más hacia el dedo I. Presenta elemento “y” y el prehallux está formado por tres elementos. El tibial y el fibular están fusionados distal y proximalmente pero separados en el medio. En el ejemplar MUJ 093 hay un elemento sesamoideo llamado cartílago plantaris (Fig. 9) entre el dedo V y el fibular ventralmente.
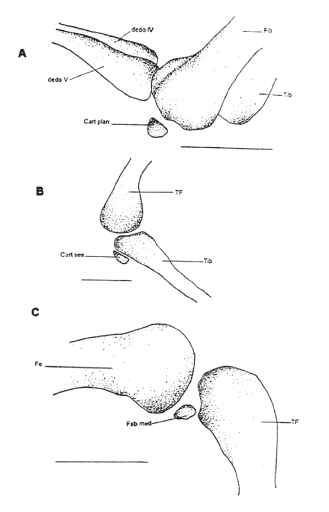
Figura 9. Elementos sesamoideos del miembro posterior. MUJ 093.
DISCUSIÓN
Huesos del cráneo. Como lo estableció Lynch (1976) en un estudio de las especies del género Eleutherodactylus de Suramérica, dentro del cual está incluida E. bogotensis, se puede corroborar que pertenece al grupo unistrigatus debido a su descripción y a los caracteres encontrados. En cuanto a la descripción ósea del grupo unistrigatus se encuentran los siguientes caracteres: frontoparietal no fusionado al proótico, rama medial de pterigoides usualmente superponiendo la pars alaris del paraesfenoide; huesos nasales largos en contacto medial o delgadamente separados. El único carácter que no está muy claro es la presencia de odontóforos desarrollados, que en la especie estudiada no alcanzan a serlo.
Lynch (1978) estableció la pérdida del cuadradoyugal en seis especies de Atelognathus, situación que no sucede en la especie estudiada y reporta que siete géneros más de leptodactílidos presentan también esta pérdida (Notaden, Crossodactylus, Pleurodema, Pseudopaludicola, Batrachyla, Hylorina e Insuetophrynus).
En el género Eleutherodactylus existen varios estudios filogenéticos y descriptivos, basados en la osteología craneal. Lynch & Ruiz-Carranza (1985) diferenciaron un grupo de Eleutherodactylus encontrando variaciones entre E. carmelitae, E. cristinae, E. delicatus, E. insignitus, E. megalops, E. ruthveni, E. sanctaemartae, E. tayrona y E. w-nigrum, destacando cambios significativos en el tamaño y forma de los nasales y frontoparietales. En todas las especies están separados estos dos pares de huesos y no hay fontanela entre los frontoparietales; los bordes de los frontoparietales tienen tres formas: transverso (E. tayrona), irregular medialmente (E. carmelitae, E. cristinae, E. insignitus, E. megalops, E. ruthvreni y E. sanctaemartae) y extendido medialmente e irregular sagitalmente (E. w-nigrum). Los palatinos están extendidos medialmente, excepto en E. tayrona.
Lynch (1989) dividió un grupo de Eleutherodactylus en tres grupos apoyado en la osteología craneal: en el primer grupo están E. dolops y E. babax donde hay una diferenciación con otros grupos, basada en el tamaño de los huesos nasales; en estas dos especies, los nasales son grandes, anchos, están en contacto medialmente y posteriormente, parcialmente con los frontoparietales los cuales se unen entre sí, pero no están fusionados al proótico; la parte palatina de la premaxila es relativamente angosta en comparación con los de E. nigrovittatus. Los vómeres son largos y relativamente angostos en su separación, los procesos alares y cultriforme del paraesfenoide son largos y delgados y perpendiculares entre sí; el extremo anterior del proceso cultriforme es puntudo y está ubicado entre los palatinos. Los cuadratojugales son muy delgados. Las ramas óticas y zigomáticas del escamoso son de igual longitud.
En el segundo grupo están E. cruralis y E. discoidalis donde los nasales están separados medialmente y el esfenetmoide está ubicado entre los nasales y los frontoparietales los cuales tienen una fontanela en E. discoidalis y no en E. cruralis donde hay fusión con los proóticos. Los processus alares de la premaxila en E. discoidalis esta dispuesto anterodorsalmente, mientras que en E. cruralis lo están posterodorsalmente, como en la mayoría de Eleutherodactylus incluyendo a E. bogotensis. Los procesos alares y cultriforme del paraesfenoide son menos delgados y cortos que los anteriores. Las demás características son similares a E. babax y E. dolops. El tercer grupo comprende E. elassodiscus, E. latens, E. mantipus y E. nigrovittatus donde difieren de los demás en el tamaño de los nasales, pues estos son más pequeños y separados medialmente igual que en el segundo grupo, excepto en E. latens. Los nasales están completamente separados de los frontoparietales en E. elassodiscus y E. mantipus; ligeramente separados en E. latens y limitándose en E. nigrovittatus. Los frontoparietales están fusionados al proótico. Los vómeres son grandes y separados medialmente. Hay diferencias con los otros grupos en la forma y disposición del proceso cultriforme del paraesfenoide porque es más grande y los palatinos son más largos. Un carácter importante es el largo proceso anterior del esfenetmoide que se extiende hasta el nivel de los nasales, al igual que en uno de los ejemplares estudiados para E. bogotensis. En E. cheiroplethus los nasales son pequeños, separados de los frontoparietales y los odontóforos de los vómeres son muy anchos (Lynch, 1990).
Lynch (1996), estudió el subgénero Pelorius, que comprende seis especies de Eleutherodactylus y analizó sus relaciones basado en osteología craneal encontrando básicamente una separación en dos grupos: el primero, donde se incluyen a E. chlorophenax, E. inoptatus y E. nortoni, con grandes y anchos nasales en contacto medial, y contacto con los frontoparietales que sostienen una cresta craneal en forma de U; vómeres separados posteromedialmente por el angosto proceso cultriforme del paraesfenoide, y procesos alares del paraesfenoide desviados a diferencia del segundo grupo dentro del cual están E. hypostenor, E. parapelates, y E. ruthae quienes presentan pequeños nasales que no están en contacto con los frontoparietles y perdieron la cresta cranial, los procesos alares y el cultriforme son perpendiculares y el cultriforme no separa los vómeres.
En contraste con otros géneros de leptodactílidos, se encuentran diferencias entre el estudio hecho por Davies (1983) en Rheobatrachus silus, lo descrito por Wild (1997) para Ceratophrys cornuta y lo encontrado en este trabajo en E. bogotensis. En primer lugar, se destaca que en Ceratophrys cornuta los frontoparietales limitan con los nasales (Wild 1997), mientras que en E. bogotensis tienen el esfenetmoide por medio al igual que en Rheobatrachus silus (Davies 1983). Sin embargo, la forma irregular de los márgenes de los frontoparietales es igual en las tres especies; otras diferencias importantes están a nivel de los nasales: mientras que en Rheobatrachus silus son muy pequeños y delgados, separados entre sí y con el esfenetmoide, en C. cornuta y E. bogotensis son anchos y gruesos, se articulan con los palatinos y cubren el esfenetmoide, aunque en C. cornuta están separados entre sí. Además, en el caso de Rheobatrachus silus (Davies 1983) el escamoso no se articula por su rama ótica a la cresta parótica; el proceso cultriforme del paraesfenoide es muy corto, no alcanza a llegar hasta el medio de los palatinos; la pars facialis de la maxila es muy delgada y sin processus orbitalis; la premaxila tiene un sólo proceso palatino; el processus alaris está dirigido posterolateralmente y los vómeres son muy pequeños, mientras que en Ceratophrys cornuta el escamoso se articula por su rama ótica al exoccipital; el proceso cultriforme del paraesfenoide llega hasta el medio de los palatinos; la pars facialis de la maxila es gruesa y con processus orbitalis; la premaxila tienen dos procesos palatinos, uno a cada lado y el processus alaris está dirigido anteroposteriormente y los vómeres son muy anchos y grandes (Wild 1997) igual a lo observado en E. bogotensis con la única diferencia que la rama ótica del escamoso se articula a la cresta parótica.
Lynch (1971) afirma que los procesos alares de la premaxila están muy bien desarrollados en todos los géneros de la familia, excepto en Myobatrachus en la cual son obsoletos; los nasales exhiben considerables variaciones en tamaño y posición con respecto a la premaxila, frontoparietal y esfenetmoide: Afirma que las especies del género Eleutherodactylus tienen grandes nasales en contacto medio, lo cual difiere con lo encontrado en la especie estudiada porque aunque son grandes no están en contacto, pero coincide en afirmar que se separan de los frontoparietales por el esfenetmoide. En general, los esqueletos de algunos géneros de leptodactílidos son variables dentro de los cuales está Eleutherodactylus.
Aparato hioideo. El aparato hioideo tiene básicamente las mismas formas y disposiciones entre los diferentes grupos de anuros, pero varía en la presencia o ausencia de alguno o varios de los procesos que se desprenden de la placa hioidea. En el caso de E. bogotensis no están presentes los procesos anterolaterales, pero sí los demás procesos variando de forma, lo cual concuerda con los estudios anteriores de la familia, como es el caso de Ceratophrys cornuta, (Wild 1997), con la diferencia de tener fusionados los proceso posteromediales a la placa hioidea y no tener procesos anteriores del hial.
Cintura pectoral y miembro anterior. De acuerdo con Duellman & Trueb (1994), la familia Leptodactylidae a la cual pertenece E. bogotensis tiene cintura pectoral arciferal, lo cual concuerda con lo observado; los cartílagos epicoracoides se superponen el uno con el otro claramente permitiendo el movimiento de toda la cintura.
Según Duellman & Trueb (1994), el húmero en muchas especies presenta una modificación en las crestas de los machos debido al dimorfismo sexual en la musculatura de los mismos. En E. bogotensis no hay dimorfismo sexual en el húmero; tanto machos como hembras poseen pequeñas crestas pero no se nota ninguna diferencia significativa entre ellas, pero se nota la asociación de estas crestas a la articulación del m. deltoideus. La mayoría de los anuros tienen cuatro dedos en la mano con una fórmula falangeal 2-2-3-3. De acuerdo con Duellman & Trueb (1994), algunos leptodactílidos tienen la última falange modificada en forma diferente a como se da en E. bogotensis que la presenta en forma de T. Los elementos individuales descritos corresponden en algunos casos a la fusión de uno o varios, como es el caso de E. bogotensis que sólo presenta seis elementos individuales por fusión de los elementos distales 3, 4 y 5. Los elementos del prepollex concuerdan con lo descrito por Duellman & Trueb (1994).
Fabrezi (1992), encontró diez morfologías diferentes (sin contar variaciones a nivel del prepollex), en los carpos de los anuros dentro del cual describió algunos Leptodactylus con cinco elementos carpianos individuales: ulnar, carpiano distal 5-4-3, elemento “y”, carpiano distal 2, radial y elemento proximal del prepollex; sin embargo, E. bogotensis se asemeja más a la morfología descrita para algunos bufónidos, dendrobátidos, artroléptidos, braquicefálidos, miobatráquidos, rinodermátidos, sooglósidos, ránidos y leptodactílidos (e. g. Eleutherodactylus discoidalis).
Esqueleto axial. El esqueleto axial de los anuros es el más modificado en comparación con los otros grupos de anfibios, lo cual ha llevado a hacer mayores trabajos de investigación (Duellman & Trueb, 1994). Los miembros de la familia Leptodactylidae son procélicos; tienen ocho vértebras presacras; la articulación entre el cráneo y la primera vértebra es de tipo I; las costillas están ausentes; el sacro tiene una articulación bicondilar con el urostilo que carece de procesos trasversos anteriormente según Duellman & Trueb, (1994), lo cual concuerda con lo observado en E. bogotensis. En la familia Leptodactylidae hay un carácter epecial en los géneros Adelotus, Heleioporus, Heleophryne, Kyarranus, Limnodynastes, Neobatrachus, Notadem y Philoria (Lynch 1971).
Un carácter importante es el tipo de articulaciones presentes: la primera vértebra presacra presenta articulación tipo I en E. bogotensis (Lynch 1969); por el contrario, las especies Atelognathus solitarius y Atelognathus somuncurensis presentan esta articulación de tipo II (Lynch 1978). En la especie Ceratophrys cornuta, perteneciente a la familia Leptodactylidae, se encontró articulación de tipo III (Wild 1997). La articulación ilio-sacro también es importante; tanto en E. bogotensis como en Ceratophrys cornuta es de tipo IIB (Emerson 1979).
En general, no hay muchas variaciones en las características de la columna vertebral, sin embargo, en el caso de Rheobatrachus silus (Leptodactylidae) se observan procesos trasversos en el urostilo; tal proceso no está presente consistentemente en cualquiera otra especie de leptodactílido australiano, aunque ocurren en la mayoría de las familias de ranas (Davies 1983). Davies (1983) sostiene que éste es un estado de carácter primitivo ya que persisten a pesar de la simplificación de los elementos pos-sacros.
Según Duellman & Trueb (1994) el urostilo representa la fusión de elementos pos-sacros. En E. bogotensis hay desarrollo de una pequeña cresta extendida sobre las dos terceras partes de su longitud y llega hasta tocar al ischium, a diferencia de otras familias donde sólo se extiende hasta la mitad o el 2/3 de la longitud del ilium.
Cintura pélvica y miembro posterior. Al igual que en la cintura pectoral, no hay diferencias muy significativas a nivel de la cintura o del miembro, sino a nivel de los pies. El carácter más significativo dentro de la cintura pélvica es la protuberancia dorsal del ilium presente en E. bogotensis, la cual también está presente en Ceratophrys cornuta (Leptodactylidae) (Wild 1997).
Un elemento importante es la existencia de huesos sesamoideos en los miembros posteriores: el primero, ubicado entre el fémur y la tibiofíbula; el segundo, entre la tibiofíbula y los tarsos, y los últimos, presentes en algunos casos en la superficie ventral del pie, lo que no estaba reportado para Eleutherodactylus. Al parecer el hecho de tener estos elementos heterotópicos no se toma como carácter muy importante (Duellman & Trueb 1994) ya que poco se describen la literatura.
Los huesos sesamoideos aún están siendo estudiados, pero se podría inferir que el hecho de la existencia de los sesamoideos tarsianos se deba a tres cosas 1) alargamiento del tendón de Aquiles; 2) ventaja en la mecánica y fuerza de traslación, y 3) formación del tendón para sostener el estrés (Nussbaum 1982).
Características particulares. En Eleutherodactylus bogotensis se encontraron algunas particularidades con respecto a la mayoría de las especies con las cuales se comparó. A continuación se hace un listado de las más destacables después de hacer un cuadro comparativo:
Maxila: con dientes.
Premaxila: processus palatina grandes con dos procesos: uno lateral romo y otro medial puntiagudo, sin tocarse entre ellos.
Frontoparietales: sin crestas, no fusionado al proótico, separado de los nasales por el esfenetmoide.
Nasales: grandes, separados medialmente, articulado a los palatinos.
Escamoso: bastante grande.
Vómeres: grandes, separados medialmente, sin odontóforos desarrollados como tales.
Paraesfenoides: procesos alares cubriendo de forma ractangular totalmente las cápsulas auditivas y proceso cultriforme largo con el extremo anterior agudo y bordes laterales biconvexos.
Palatinos: largos, articulados a la maxila y con el extremo proximal del proceso cultriforme entre ellos.
Esfenetmoide: variación en la longitud del proceso anterior. Puede llegar hasta el margen anterior de los nasales o solo hasta el posterior.
Aparato hioideo: presencia de procesos anteriores del hiale y ausencia de procesos anterolaterales.
Cintura pélvica: variación en el pubis; puede ser óseo o cartilaginoso. Presencia de cresta ilial y protuberancia dorsal.
Elementos sesamoideos: presencia de fabella medialis, cartílago sesamoideo y cartilago plantaris en el miembro posterior; y cartílagos sesamoideos en la articulación ilio-sacra.
CONCLUSIONES
Se reafirma la pertenencia de E. bogotensis al grupo unistrigatus basado en caracteres osteológicos craneales: frontoparietal no fusionado al proótico; rama medial de pterigoides usualmente superpuesta con la pars alaris del paraesfenoide; huesos nasales grandes en contacto medial o delgadamente separados. El único carácter que no corresponde es la presencia de odontóforos desarrollados, que en la especie estudiada no alcanzan a serlo. Existen variaciones interespecíficas sólo a nivel del esfenetmoide y del pubis. No hay caracteres únicos para definir la especie.
LITERATURA CITADA
1. Andersen, M. L. 1978. The comparative miology and osteology of the carpus and tarsus of selected anurans. Ph.D. Thesis. [ Links ]
2. Anónimo. 1973. Nomina Anatomica Veterinaria. Viena
3. Clarke, B. A. 1981. Comparative osteology and evolutionary relationship in the African Raninae (Anura Ranidae). Italian Journal of Zoology 14: 285-331. [ Links ]
4. Clarke, B. A. 1987. A description of the skeletal morphology of Barbourula (Anura: Discoglossidae) with comments on its relationships. Journal of Natural History 21: 879-891. [ Links ]
5. Davies, M. 1983. Skeleton.Págs. 58-68 en: M. J. Tyler (ed.).The gastric brooding frog. Croom Helm, Londres. [ Links ]
6. Davies, M & T. Burton. 1982. Osteology and myology of the gastric brooding frog Rheobatrachus silus Liem (Anura: Leptodactylidae). Australian Journal of Zoology 30: 503-521. [ Links ]
7. De Sá, R. & L. Trueb. 1991. Osteology, skeletal development, and chondocranial structure of Hamptophryne boliviana (Anura: Microhylidae). Journal of Morphology 209:311-330. [ Links ]
8. Dingerkus, G. & L.D. Uhler. 1977. Enzyme clearing of alcian blue stained whole small vertebrates for demonstration of cartilage. Stain technology. 52: 229-231. [ Links ]
9. Drewes, R. 1984. A phylogenetic analysis of the Hyperoliidae (Anura): Treefrogs of Africa, Madagascar, and the Seychelles Islands. California Academy of Sciences 1-70.
10. Duellman, W. & L. Trueb. 1994. Biology of Amphibians. University Press. Londres. [ Links ]
11. Ecker, A. 1971. The anatomy of the frog (Traducido por G. Haslam). A. Asher R. CO N. V. [ Links ]
12. Emerson, S. A. 1979. The ilio-sacral articulation in frogs form and function. Biological Journal of the Linnean Society 11: 153-168. [ Links ]
13. Fabrezi, M. 1992. El carpo de los anuros. Alytes. 10:1-29. [ Links ]
14. Fabrezi, M. 1993. The anuran tarsus. Alytes. 11:47-63. [ Links ]
15. Fabrezi, M. & P. Albrech. 1996. The carpal elements of anurans. Herpetologica. 52: 188-204. [ Links ]
16. Hoyos, J.M. 1991. Aspectos taxonómicos y patrones de diseño de Eleutherodactylus bogotensis del Parque Nacional Natural Chingaza. Cuadernos Divulgativos 18: 1-12. [ Links ]
17. Hoyos, J.M. 1999. Phylogenie des Ranidae (Amphibia, Anura). Museum National D´Historie Naturelle, Laboratorie des Reptiles el Amphibiens. Thèse pour obtenir le grade de Docteur és Sciences, discipline: Systématique. [ Links ]
18. Laurent, R. & M. Fabrezi. 1989-90. Further data on carpal structure in ranoid frogs. Alytes 8: 41-50. [ Links ]
19. Lynch, J.D. 1969. Evolutionary relationships, osteology, and zoogeography of leptodactylid frogs. Ph.D. Thesis. [ Links ]
20. Lynch, J. D. 1971. Evolutionary relationships, osteology, and zoogeography of Leptodactylid frog. Museum of Natural History. University of Kansas. Lawrence, Kansas 53:1-238. [ Links ]
21. Lynch, J.D. 1976. The species groups of the South American frogs of the genus Eleutherodactylus (Leptodactylidae). Museum of Natural History, University of Kansas. Lawrence, Kansas 61:1-24. [ Links ]
22. Lynch, J.D. 1978. A re-assessment of the Telmatobiine Leptodactylid frog of Patagonia. Museum of Natural History, University of Kansas, Lawrence, Kansas 72: 1-57. [ Links ]
23. Lynch, J.D. 1993. The value of the m. depressor mandibulae in phylogenetic hypotheses for Eleutherodactylus and its allies (Amphibia: Leptodactylidae). Herpetologica. 49: 32-41. [ Links ]
24. Lynch, J.D. & P.M. Ruiz-Carranza. 1985. A synopsis of the frogs of the genus Eleutherodactylus from the Sierra Nevada de Santa Marta, Colombia. Museum of Zoology, University of Michigan 711: 1-59.
25. Lynch, J. D. & W. Duellman. 1997. Frogs of the genus Eleutherodactylus in Western Ecuador: Systematics, Ecology and Biodiversity. Natural History Museum, Dyche Hall, Lawrence, Kansas. [ Links ]
26. Nussbaum, R. 1982. Heterothopic bones in the hindlimbs of frogs of the families Pipidae, Ranidae and Sooglossidae. Herpetologica 38: 312-320. [ Links ]
27. Raikow, R., A. Bledsoe, B. Myers & C. Welsh. 1990. Individual variation in avian muscles and its significance for the reconstruction of phylogeny. Systematic Zoology 39(4): 362-370. [ Links ]
28. Ritland, R. 1955. Studies on the postcranial morphology of Ascaphus truei. Journal of Morphology 97:119-178. [ Links ]
29. Rivero, J. & M. Serna. 1987. Tres nuevas especies de Eleutherodactylus (Amphibia, Leptodactylidae) de Antioquia, Colombia. Journal of Science 23: 386-399. [ Links ]
30. Sheil, C. 1999. Osteology and skeletal development of Pyxicephalus adspersus (Anura: Ranidae: Raninae). Journal of Morphology. 240: 49-75. [ Links ]
31. Wild, E. 1997. Description of the adult skeleton and developmental osteology of the Hyperossified Horned frog, Ceratophrys cornuta (Anura: Leptodactylidae). Journal of Morphology 232: 169-206. [ Links ]
32. Zug, G.R, L.J. Vitt, & J.P. Caldwell. 2001. Herpetology. An Introductory Biology Of Amphibians and Reptiles. Academic Press, Second Edition. USA. 630 pp. [ Links ]
Recibido: 18/10/2004
Aceptado: 01/07/2005

