










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.28 no.1 Bogotá June 2006
MORPHOLOGICAL CONSTRAINTS AND NECTAR ROBBING IN THREE ANDEAN BUMBLE BEE SPECIES (HYMENOPTERA, APIDAE, BOMBINI)
Limitantes morfológicos y robo de néctar de tres especies de abejorros andinos (Hymenoptera, Apidae, Bombini)
ANDRE J. RIVEROS
Center for Insect Science and Division of Neurobiology, University of Arizona. 611 Gould-Simpson Science Building, 1040 East Fourth Street. P.O. Box 210077, Tucson, AZ 85721, USA. ajosafat@email.arizona.edu
EDGAR J. HERNÁNDEZ
Department of Biology, University of Missouri-St. Louis, One University Boulevard, St. Louis, MO 63121, USA. ejh983@umsl.edu
GUIOMAR NATES-PARRA
Laboratorio de Investigación en Abejas (LABUN), Departamento de Biología, Universidad Nacional de Colombia, Apartado 14490, Bogotá D. C., Colombia. mgnatesp@unal.edu.co
ABSTRACT
We report differences in foraging behavior of three Andean bumblebee species on flowers of Digitalis purpurea (Scrophulariaceae). Bombus atratus was a potential pollinator while B. hortulanus and B. rubicundus collected nectar by robbing through holes. We attribute behavioral differences to physical constraints. B. atratus has a longer glossa and a larger body size and is able to reach the nectaries, whereas B. hortulanus and B. rubicundus have shorter glossae and smaller bodies and probably must rob nectar through holes at the base of flowers.
Key words. Apidae, Bombus, Digitalis purpurea, nectar robbing.
RESUMEN
Se reportan diferencias en el comportamiento de forrajeo de tres especies de abejorros en flores de Digitalis purpurea (Scrophulariaceae). Bombus atratus fue un polinizador potencial, mientras que B. hortulanus y B. rubicundus colectaron néctar robándolo a través de agujeros. Nosotros atribuimos las diferencias conductuales a limitantes físicos; mientras que B. atratus puede alcanzar fácilmente los nectarios por su glosa larga y un cuerpo más pequeño, B. hortulanus y B. rubicundus deben utilizar agujeros en la base de las flores para obtener el recurso por sus glosas más cortas y un cuerpo más grande.
Palabras clave. Apidae, Bombus, Digitalis purpurea, robo de néctar.
INTRODUCTION
The foxglove, Digitalis purpurea L. (Scrophulariaceae), is a perennial herb of 50–180 cm tall, which has a spike of flowers for the first half of the year. This plant is originally from Europe, but now is widely distributed in many areas of the world as a garden plant. The flowers are campanulate, with long (35–50 mm) corolla tubes; each flower has a very conspicuous constriction near the base of the corolla. Digitalis flowers are visited by several hummingbirds and bumblebee species in temperate zones; the latter are commonly referred to as the usual visitors (e.g. Percival & Morgan 1965, Percival et al. 1968, Best & Bierzychudek 1982).
MATERIALS AND METHODS
In the Eastern Andes of Colombia (La Calera, Cundinamarca) at 2800 m altitude, we observed the foraging behavior of three bumble bee species (Bombus atratus Franklin, 1913, B. hortulanus Friese, 1904 and B. rubicundus Smith, 1854) on two patches of D. purpurea. Some individuals took nectar through holes made in the base of flowers (nectar robbers), while others took nectar from inside the corolla and sometimes carried pollen (legitimate visitors). We hypothesize that nectar-robbing behavior is related to the bee body size and floral morphology, as these elements allow or prevent access to the nectaries when legitimate visits occurs.
Behavioral observations were made over a period of 70 hours from January to June 2000, usually from 8:00 to 14:00 hrs. The body size of Bombus was estimated by measuring the length of the extended glossa and the forewing (WL) (Morse 1977). We also collected and measured the following variables on 26 flowers of D. purpurea: distance between the hole (used for taking nectar) and the nectaries (DHN), and the distance between the nectaries and the maximum distance reached by a legitimate visitor, which is determined by a constriction close to the base of the flower and the length of the ovary (DCN) (Figure 1).
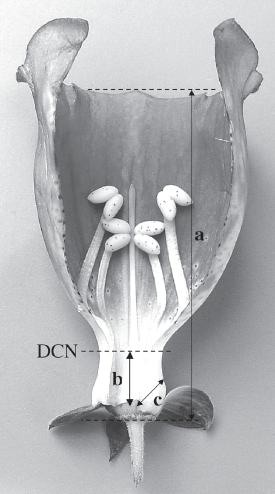
Figure 1. Floral measurements taken from Digitalis purpurea. a = Total length, b = Distance constriction-nectaries (DCN), c = Distance hole-nectaries (DHN).
RESULTS AND DISCUSSION
A total of 22 individuals of the three species of bumble bees were observed. Workers of Bombus atratus were the most frequent legitimate visitors, while B. hortulanus (except for three of 11 individuals with glossa long enough to reach nectaries) and B. rubicundus always behaved as nectar robbers. Robbers took nectar through holes located in the base of the corolla, at a distance sufficiently short as to reach the nectaries with their tongues (DHN: X =4.38 mm, s2 = 0.95, N = 26). A considerable number of flowers had perforations in the base of the corolla (77 %, N = 200), suggesting a high frequency of nectar robbery in this plant population.
On average, workers of Bombus atratus foraging on D. purpurea had a longer glossa (t- test: P < 0.001, t = 6.1, N1 = 11, N2 = 11) and smaller body size (t-student test for wing length: P < 0.001, t = 9.46, N1 = 11, N2 =11) than workers of B. hortulanus and B. rubicundus [since sample size for B. rubicundus was too small (N = 2), we considered two groups for analyzing: legitimate visitor (B. atratus) and “robbers” (B. hortulanus plus B. rubicundus), differences found were not affected by grouping]. Although we never observed bees perforating the corolla, opening holes in flowers for nectar robbery is commonly reported in bumble bees (Morse 1982 and references therein, reviewed by Maloof & Inoue 2000). Instead, hummingbirds frequently exhibited this behavior, suggesting that at least in some cases Bombus could be a secondary nectar robber. As shown, for example, by birds using holes made by robbing Trigona bees (Roubik et al., 1985)
Our observations suggest a strong effect of morphological constraints in bumble bee nectar robbing. A longer glossa and smaller body size may enable Bombus atratus workers to easily reach nectaries from the flower constriction (DCN: X = 7.5 mm, s2 = 2.1, N = 26), whereas a larger body size (WL, X =14.6 mm, s2 = 3.0) and shorter glossa (X = 6.9 mm, s2 = 0.4) prevent B. hortulanus and B. rubicundus from getting nectar legally; thus, forcing individuals to reach nectar through perforated flowers [but see Percival et al. 1968 for examples where inaccessibility to nectaries produces changes in plant preference].
Nevertheless, the presence of few individuals of Bombus hortulanus with a long glossa but exhibiting nectar robbing on Digitalis may suggest a behavioral attribute that would not correspond strictly with the floral-bee. First, robbing behavior could be the result of a successful early visit, which can induce specialization in foraging tactics. An early visit to flowers with narrow corollas and deep nectaries should produce exploratory behavior on the flower, by those unsuccessful bees unable to take nectar legally, and probably a tendency to perforate and rob (Morse 1982, Maloof & Inoue 2000, see Brian 1957 for a comparison between instinctive and learned hypothesis of robbing in bumble bees). Alternatively, forager bees could be attracted to the base of the corolla after observing other bees exploring the corolla base, a form of local enhancement. Transferring of information through this kind of social learning has been recently demonstrated in bumble bee foragers (Leadbeater & Chittka, 2005; Worden, B. D. personal communication). If a mechanism like local enhancement is involved in robbing behavior has not been previously tested. If present, bees can be attracted to the corolla base and then, as consequence of exploration, exhibit secondary robbing behavior when holes are found and nectar is obtained.
ACKNOWLEDGMENTS
We thank William T. Wcislo, Robert B. Srygley, Victor H. Gonzalez and two anonymous referees for their critics and help to improve this manuscript.
LITERATURE CITED
1. Best, L. S. & P. Bierzychudek. 1982. Pollinator foraging on foxglove (Digitalis purpurea): A test of a new model. Evolution 36: 70-79. [ Links ]
2. Brian, A. 1957. Differences in the flowers visited by four species of bumble-bees and their causes. Journal of Animal Ecology 26: 71-98. [ Links ]
3. Leadbeater, E. & L. Chittka. 2005. A new mode of information transfer in foraging bumblebees? Current Biology 15: R447-R448. [ Links ]
4. Maloof, J. & D. Inouye. 2000. Are Nectar Robbers Cheaters or Mutualists? Ecology 81: 2651-2661. [ Links ]
5. Morse, D. H. 1977. Estimating proboscis length from wing length in bumblebees (Bombus spp.). Annals of the Entomological Society of America 70: 311-315. [ Links ]
6. Morse, D. H. 1982. Behavior and ecology of bumblebees. Pp. 245-322. In: H. R. Hermann (ed.), Social Insects, Vol. III, Academic Press, New York. [ Links ]
7. Percival, M. S. & P. Morgan. 1965. Observations on the floral biology of Digitalis species. New Phytologist 64: 1-22. [ Links ]
8. Percival, M. S., P. Morgan, P. & D. E. Lewis. 1968. Behaviour of Bombus on Digitalis spp. New Phytologist 67: 759-769. [ Links ]
9. Roubik, D.W., N.M. Holbrook & G. Parra. 1985. Roles of nectar robbers in reproduction of the tropical treelet Quassia amara (Simaroubaceae). Oecologia 66: 161–167.
10. Wilson P. 1995. Selection for pollination success and the mechanical of Impatiens flowers around bumblebee bodies. Biological Journal of the Linnean Society 55: 355-383. [ Links ]
Recibido: 22/10/2005
Aceptado: 17/04/2006

