










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.28 no.2 Bogotá Dec. 2006
POTENCIAL DEL BANCO DE SEMILLAS EN LA REGENERACIÓN DE LA VEGETACIÓN DEL HUMEDAL JABOQUE, BOGOTÁ, COLOMBIA
Potential of the seed bank in the regeneration of plant communities at Jaboque wetland, Bogotá, Colombia
ALBA L. MONTENEGRO-S.
YUDY A. ÁVILA PARRA
HOOZ A. MENDIVELSO-CH.
ORLANDO VARGAS
Departamento de Biología, Universidad Nacional de Colombia, Bogotá, D.C., Colombia. almontenegros@unal.edu.co; yaavilap@unal.edu.co; hachaparrom@unal.edu.co; jovargasr@unal.edu.co
RESUMEN
Se estudió el banco de semillas germinable (BS) en seis parches de vegetación en la zona conservada del humedal Jaboque: 1) Rumex conglomeratus - Polygonum sp. 2) Juncus effusus - Polygonum sp. 3) Typha latifolia 4) Juncus effusus 5) Pennisetum clandestinum - Polygonum sp. 6) Pennisetum clandestinum – Holcus lanatus. En cada parche se evaluó la composición, la densidad de las especies presentes en el BS) el tipo de BS que poseen algunas de las especies nativas del humedal) la similitud entre la composición del BS y la vegetación. El BS del humedal Jaboque actualmente presenta un estado de degradación variable en todos los parches, revelado por la alta dominancia de unas pocas especies y la presencia de semillas de especies oportunistas. Por lo anterior, si el BS se expresa luego de un disturbio, su potencial para la regeneración de los parches es bastante limitado; bajo las condiciones generadas por el disturbio, las especies oportunistas pueden germinar y ser favorecidas conduciendo a la competencia, el reemplazamiento y/o la eliminación de las especies del humedal. Con base en la presencia de las especies propias del humedal en el BS y en los parches de vegetación se evidencian tres tipos de vulnerabilidad a la extinción local, 1) Especies muy vulnerables: son susceptibles a la extinción local, debido a que tienen baja frecuencia en la vegetación y están ausentes en el BS (Carex luridiformis y Ludwigia peruviana) o a que están ausentes en la vegetación y presentes en el BS (Juncus microcephalus y Eleocharis spp.), 2) Especies vulnerables: presentan bajas frecuencias en la vegetación y forman algún tipo de BS (Cyperus rufus) y 3) Especies no vulnerables: son poco susceptibles a la desaparición local por poseer mecanismos para asegurar su permanencia, principalmente mediante la regeneración clonal (Ludwigia peploides y Typha latifolia) o combinada con la reproducción sexual, además presentan la formación de BS grandes de tipo persistente, cuyas semillas germinan con facilidad (Juncus effusus, Polygonum sp. y Bidens laevis).
Palabras clave. Banco de semillas germinable, regeneración de la vegetación, especies oportunistas, humedales neotropicales.
ABSTRACT
The germinable seed bank (SB) was studied in six patches of vegetation in the preserved area of the Jaboque wetland, 1) Rumex conglomeratus - Polygonum sp. 2) Juncus effusus - Polygonum sp. 3) Typha latifolia 4) Juncus effusus 5) Pennisetum clandestinum - Polygonum sp. 6) Pennisetum clandestinum – Holcus lanatus. In each patch was evaluated) the composition and density of the SB species) the type of SB that possesses some of the native species of the wetland, and the similarity between composition of the SB and the vegetation. Currently, the SB of Jaboque wetland has a variable degradation in all patches, due to high dominance of few species and the presence of opportunity species seeds in all vegetation patches. Therefore, after a disturbance the SB has a low potential to reestablishment of wetland vegetation. Besides, if the generated conditions by the disturbance allow to germination and establishment of opportunist species, these can contribute to the replacement and elimination of native wetland species. Based on the presence of own species of the wetland in the SB and the vegetation patches, it is evident there are three types of vulnerability to the local extinction. 1) Highly vulnerable species: are susceptible species to the local extinction, since they have low frequency in the vegetation and they are absent in the SB (Carex luridiformis and Ludwigia peruviana) or because they are absent in the vegetation and they are present in the SB (Juncus microcephalus and Eleocharis spp.), 2) Vulnerable species: are species with low frequency in the vegetation and they have some type of SB (Cyperus rufus) and 3) Not vulnerable species: are species with low susceptibility to the local extinction because they have mechanisms to guarantee their survival, mainly by the clonal regeneration (Ludwigia peploides and Typha latifolia) or by the combination of this with the sexual reproduction, besides these species have a large SB of persistent type, whose seeds germinate easily (Juncus effusus, Polygonum sp., and Bidens laevis).
Palabras clave. Germinable seed bank, regeneration of vegetation, opportunist species, Neotropical wetland.
INTRODUCCIÓN
El banco de semillas (BS) es una agregación de semillas viables (Simpson et al. 1989) no germinadas presentes en el suelo, potencialmente capaces de reemplazar plantas adultas anuales o perennes (Harper 1977, Baker 1989). Constituye el principal medio para el reestablecimiento de la vegetación, particularmente de las especies cuya supervivencia de propágulos vegetativos durante periodos de sequías largos no es posible; sin embargo, la importancia del BS parece estar restringida a las comunidades de vegetación emergente ya que Boedeltje et al. (2003) concluyeron que el BS no juega un papel importante en la revegetalización de especies acuáticas sumergidas.
Las semillas de las especies que forman BS a menudo entran en un periodo de latencia una vez son liberadas de la planta parental; esta estrategia les permite tolerar la variación climática, las fluctuaciones en la disponibilidad de nutrientes y sobrevivir a periodos desfavorables prolongados (Bliss & Zedler 1998), es así como algunas especies de plantas sobreviven a las épocas secas debido a la formación de un gran BS durante periodos de disponibilidad de agua (Warwick & Brock 2003).
La germinación y el posterior reclutamiento de las plántulas emergidas a partir del BS son considerados los procesos más críticos que determinan la estructura final de la comunidad vegetal (Parker et al. 1989). Durante la sucesión secundaria, la composición de especies del BS puede ayudar a determinar la estructura de la comunidad de plantas luego de un disturbio, indicando cuáles especies pueden llegar a establecerse en un sitio, si las condiciones son adecuadas para su germinación (Cronk & Fennesy 2001). Por su parte, la respuesta del reclutamiento a la heterogeneidad medioambiental es la que finalmente influye en la diversidad de especies y en el patrón espacial observado en la vegetación (Peterson & Baldwin 2004, Leck & Simpson 1995, Galinato & Van der Valk 1986, Van der Valk 1981).
En cuanto al tipo de BS que presenta cada especie, éste puede ser transitorio o persistente. El primero se caracteriza por poseer una viabilidad de las semillas inferior a un año (Thompson & Grime 1979), presentar un único evento de germinación (Walck et al. 1996, Baskin & Baskin 2001) y encontrarse en la vegetación o en el nivel superior del suelo (Thompson et al. 1997). Por su parte, el BS tipo persistente presenta semillas con viabilidad por mucho tiempo e incluso siglos (Thompson & Grime 1979), más de un evento de germinación (Walck et al. 1996, Baskin & Baskin 2001) y se encuentra en los niveles intermedios del suelo -persistentes a corto plazo- o predominando en el nivel inferior del suelo -persistentes a largo plazo- (Thompson et al. 1997).
BS de tipo persistente es el más importante desde el punto de vista de la regeneración (Garwood 1989), es característico de comunidades que están sujetas a disturbios frecuentes pero impredecibles, como son cultivos, fuego o fluctuaciones del nivel de agua (Grime 1979, Pickett & McDonnell 1989, Fenner 1995). Representa sólo una parte de las especies de la comunidad y está compuesto generalmente por las especies ruderales (Garwood 1989, Louda 1989, Pickett & McDonnell 1989) y por las plantas dominantes (Louda 1989).
Los BS en los humedales son considerados como reservas naturales de semillas que se pueden usar para la restauración de la vegetación (Van der Valk & Pederson 1989), ya que permiten una rápida regeneración del conjunto de especies nativas (Galatowitsch & Van der Valk 1996), conservando en gran medida la diversidad y la estructura de la comunidad gracias a su alta densidad (Vavrek & Campbell 2005). En los proyectos de restauración el conocimiento del BS es particularmente útil, debido a que su composición provee una idea de las especies que colonizarán un lugar cuando la hidrología del humedal sea restaurada (Cronk & Fennessy 2001).
El humedal Jaboque al igual que todos los humedales de la Sabana de Bogotá ha estado sometido a presiones tanto naturales como antrópicas, siendo estas últimas las que están deteriorando aceleradamente los componentes bióticos y abióticos del sistema. Entre los agentes tensionates del humedal están: 1) asentamientos humanos ilegales que favorecen procesos de relleno en éste (EAAB & CI 2000), 2) construcción de avenidas y la canalización del río Bogotá (DAMA 2000, EAAB & CI 2000), 3) vertimiento de aguas servidas y basuras de los barrios e industrias (DAMA 2000), 4) utilización de las zonas de ronda para el pastoreo y cultivos, 5) uso del agua del humedal para riego 6) presencia de animales domésticos (p.e. perros) y 7) actividad de carboneras.
Lo anterior ha llevado a cambios en la dinámica de las comunidades vegetales permitiendo la expansión de especies emergentes y flotantes libres, reduciendo el espejo de agua y favoreciendo el ingreso de especies oportunistas terrestres (Pennisetum clandestinum, Trifolium repens, Gnaphalium americanum, Solanum americanum), conduciendo a la degradación de las comunidades. Con el fin de evaluar el potencial de regeneración por BS de diferentes parches de comunidades vegetales del Humedal Jaboque se plantearon los siguientes cuestionamientos ¿Cómo cambia la composición, la densidad y la diversidad de especies en los BS en los diferentes parches?, ¿Qué tipos de BS presentan las especies propias del humedal? y ¿La composición de especies del BS refleja la composición de la vegetación en cada uno de los parches?
MATERIALES Y MÉTODOS
Área de Estudio
El estudio se realizó en el humedal Jaboque, ubicado en la localidad de Engativá (Bogotá D.C.), en zonas topográficamente bajas, conformadas por una pequeña cuenca bordeada por el río Juan Amarillo y el área que ocupa el Aeropuerto El Dorado. El humedal ocupa un área total de 80 hectáreas (EAAB 2000). En el estudio “investigación aplicada para la restauración del humedal Jaboque (Convenio Universidad Nacional de Colombia – Empresa de Acueducto y Alcantarillado de Bogotá -ESP)” se estableció la sectorización del humedal en tres zonas según su estado de degradación: zona conservada (a pesar de su estado de degradación), zona con alteración intermedia y zona muy alterada. Este trabajo se realizó en la zona conservada.
Muestreo
Se tuvo en cuenta el mapa de comunidades vegetales del humedal Jaboque elaborado por Hernández et al. (2005) y observaciones de campo (Fig. 1). Con el fin de comparar la composición del BS con la vegetación en pie, se muestreo un parche de vegetación en cada una de las seis comunidades más grandes y accesibles de la zona conservada (Tabla 1). El BS fue evaluado mediante el método de emergencia de plántulas, para lo cual en cada parche se tomaron 20 núcleos de suelo al azar, los cuales fueron tratados como una muestra. Cada núcleo se tomó hacia el centro del parche para reducir el efecto de borde, empleando una sonda de extracción de suelo de 5,4 cm de diámetro y 10 cm de profundidad (volumen total de 4580 cm3).

Figura 1. Mapa del humedal Jaboque (diseño H. Arellano). Puntos de muestreo del banco de semillas en las comunidades de: 1. Juncus effusus-Polygonum sp., 2. Rumex conglomeratus-Polygonum sp. 3. Typha latifolia. 4. Pennisetum clandestinum-Holcus lanatus. 5. Juncus effusus. 6. Pennisetum clandestinum- Polygonum sp.
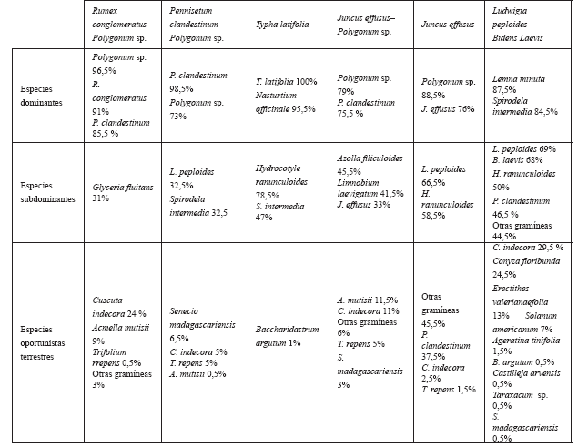
Tabla 1. Comunidades vegetales seleccionadas a partir de Hernández et al. (2005) y especies dominantes en la vegetación de cada parche estudiado.

Todas las muestras fueron empacadas en bolsas plásticas, etiquetadas, selladas y transportadas a un invernadero de la Universidad Nacional de Colombia, donde fueron extendidas en bandejas germinadoras (siguiendo la metodología propuesta por Montenegro 2000), con un espesor de 1 cm aproximadamente y se removieron cada mes para facilitar la incidencia de luz. Se permitió la expresión del BS durante ocho meses, en los primeros cuatro meses se dejaron a capacidad de campo y en los siguientes cuatro en sobresaturación, con el fin de permitir diferentes condiciones para la germinación. Para calcular la densidad de semillas, se utilizó el área ocupada por las muestras en las bandejas, expresándola como número de semillas germinadas por metro cuadrado (s/m2).
Para complementar la caracterización de la vegetación (Hernández et al., 2005) se realizaron levantamientos en los diferentes parches utilizando el método de línea-intercepto, de acuerdo a lo propuesto por Matteucci & Colma (1982). Se trazaron diez líneas cada una de 10 m, separadas entre sí aproximadamente 3 m, éstas fueron utilizadas como transectos, sobre los cuales se determinó la presencia o ausencia de las especies a intervalos de 0,5 m.
Análisis de Datos
Con el fin de determinar la diversidad del BS en los diferentes parches se utilizó el índice de Shannon. Para establecer el tipo de BS para las especies encontradas con mayor frecuencia en todos los parches (Bidens laevis, Cuscuta indecora, Cyperus rufus, Hydrocotyle ranunculoides, Juncus effusus, Polygonum sp. y Rorippa pinnata) se empleó el índice de persistencia propuesto por Hölzel & Otte (2004), AV/SBfrec = (SBf /(SBf + AVf)) * 100, donde AVf es la frecuencia de la especie en la vegetación y SBf la frecuencia de la especie en el BS. Este índice puede tener valores desde cero (especie no formadora de BS, presente sólo en vegetación), hasta 100 (especie con un BS altamente persistente).
Para determinar el grado de similitud entre la composición de la vegetación y el BS germinable se utilizó el índice de Sørensen: Q = 2c/(a+b), donde a es el número de especies en el BS, b el número de especies en la vegetación y c el número de especies compartidas.
RESULTADOS
Frecuencia de las especies presentes en la vegetación
La mayoría de los parches de vegetación presentan dos especies con frecuencia alta (>70%). En general son especies típicas del humedal con excepción de los parches de Pennisetum clandestinum –Polygonum sp., Juncus effusus – Polygonum sp. donde una de las especies dominantes es la invasora Pennisetum clandestinum con una frecuencia del 98,5 % y 75,5%, respectivamente. Por otra parte las especies oportunistas terrestres fueron comunes a todos los parches de vegetación estudiados excepto para el de Typha latifolia, que tuvo una sola especie (Baccharidastrum argutum) (Tabla 2).
Tabla 2. Composición de la vegetación actual de diferentes parches de vegetación del humedal Jaboque. Se indica la frecuencia de aparición de la especie en %.
Densidad y composición del banco de semillas del Humedal Jaboque
Existieron valores muy extremos en la densidad de semillas entre los diferentes parches de vegetación, el dominado por Juncus effusus presentó 40935 s/m2, mientras el de Ludwigia peploides- Bidens laevis presentó tan sólo 662 s/m2 (Fig. 2).
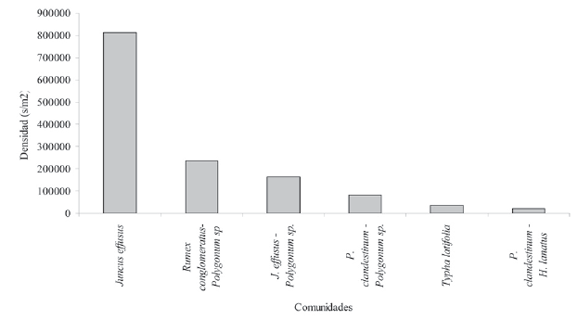
Figura 2. Densidad total de semillas para los diferentes parches de vegetación del humedal Jaboque.
La distribución de la abundancia (s/m2) de las especies en los diferentes parches genera tres patrones; el primero está dado por la alta dominancia de una única especie y la reducción abrupta en la densidad, debido a que la mayoría de las especies presentan densidades bajas (parche de Juncus effusus), el segundo patrón corresponde a la dominancia de varias especies y la reducción abrupta en la densidad, debido a que la mayoría presenta bajas densidades (Pennisetum clandestinum -Polygonum sp. y de Rumex conglomeratus-Polygonum sp.) y el tercero presenta varias especies dominantes y una reducción gradual en la densidad dada por especies con densidades intermedias (parches de Juncus effusus – Polygonum sp., Typha latifolia y Pennisetum clandestinum – Holcus lanatus, Fig. 3).
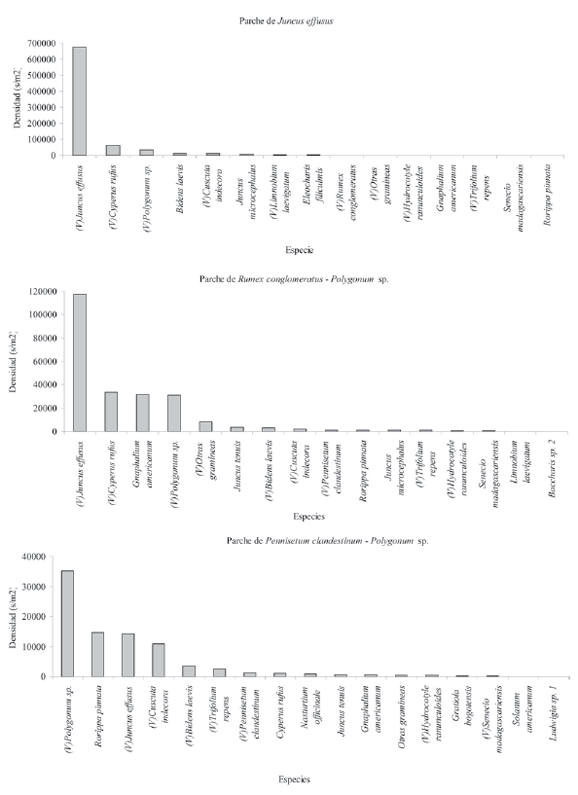
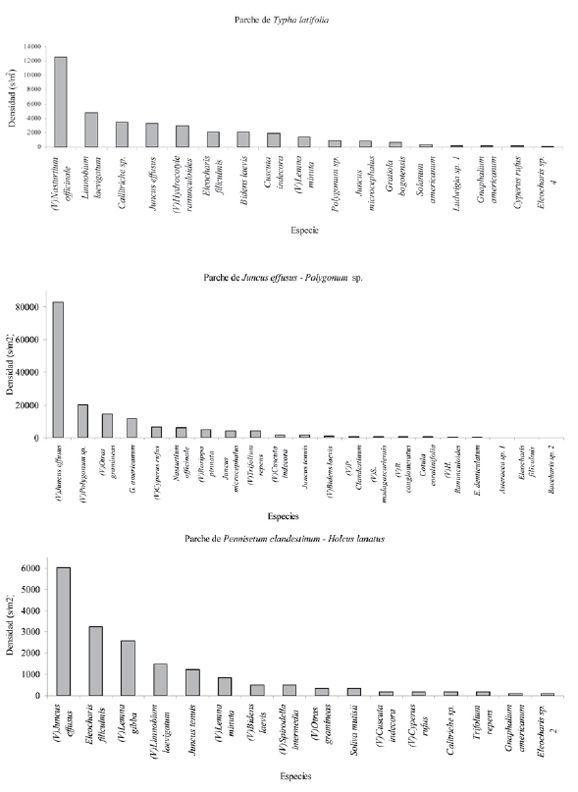
Figura 3. Densidad de semillas por especie para los parches de vegetación, (V) especies presentes en la vegetación. Patrones de abundancia.
En cuanto a la composición de especies, Juncus effusus es la más frecuente (presente en todas los parches de vegetación) y a la vez la más abundante. En el parche que lleva su nombre conforma el 83% (34031 s/m2) del BS. Otras especies que también están presentes en todos los parches son Polygonum sp. (entre 4 y 1879 s/m2), Bidens laevis (entre 22 y 537 s/m2) y Cyperus rufus (entre 8 y 3200 s/m2).
Es de resaltar que algunas especies dominantes en la vegetación presentan densidades muy bajas o nulas en el BS tal es el caso de Rumex conglomeratus y Typha latifolia, mientras que otras especies a pesar de no estar en la vegetación exhiben densidades relativamente altas en el BS, es el caso de Eleocharis filiculnis (106 s/m2), Juncus tenuis (78 s/m2) en el parche de vegetación de Ludwigia peploides – Bidens laevis y Juncus microcephalus (41 s/m2) en el parche de Juncus effusus – Polygonum sp. (Fig. 3).
La parásita Cuscuta indecora, a pesar de presentar una baja frecuencia de aparición en la vegetación de todos los parches (no superó 30%, Anexo 1) tiene densidades entre 8 y 561 s/m2, ésta última densidad corresponde al parche de Pennisetum clandestinum – Polygonum sp.
Anexo 1. Presencia de especies en la vegetación, en el banco de semillas y compartidas.
En el BS se presentan especies oportunistas terrestres (Gnaphallium americanum, Senecio madagascariensis, Trifolium repens, Solanum americanum, Pennisetum clandestinum y el conjunto de otras gramíneas), las cuales están en todos los parches. En conjunto las densidades más bajas corresponden al parche de Typha latifolia (25 s/m2) y las más altas al de Juncus effusus – Polygonum sp. (2231 s/m2), es de resaltar que en este último la especie Gnaphalium americanum presenta la mayor densidad (1582 s/m2) de este conjunto (Fig. 3). La especie Pennisetum clandestinum a pesar de presentarse con gran frecuencia en la vegetación, solamente está en el BS de tres parches con densidades bajas (Rumex conglomeratus – Polygonum sp., Juncus effusus - Polygonum sp. y Pennisetum clandestinum - Polygonum sp., con valores inferiores a 153 s/m2, Fig. 3)
Persistencia de las especies más comunes del BS
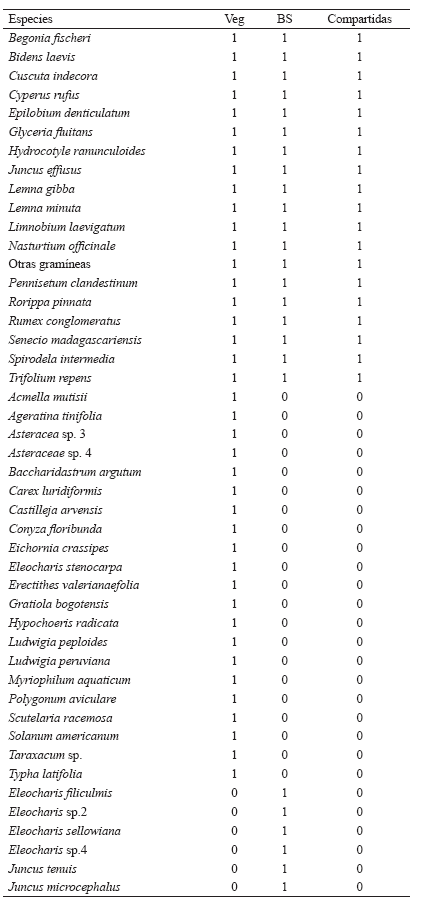
Juncus effusus presenta un BS persistente, sus semillas pueden formar parte del BS en algunos parches donde esta especie no está formando parte de la vegetación (parche de Rumex conglomeratus-Polygonum sp.), exhibiendo los valores más altos del índice de persistencia. También puede darse el caso en que la frecuencia de aparición de la especie en el BS y en la vegetación sea similar generando un valor del índice cercano a 50 (parche de Juncus effusus). Por su parte, Cyperus rufus muestra una tendencia persistente en el BS de todos los parches (Fig. 4). Rorippa pinnata tiene los valores más altos de persistencia (excepto en el de Juncus effusus – Polygonum sp.) indicando su escasa presencia en la vegetación. Cuscuta indecora tiene un BS tipo persistente aunque en algunas comunidades su frecuencia de aparición en la vegetación es baja (parche de Pennisetum clandestinum-Holcus lanatus, Fig. 4).
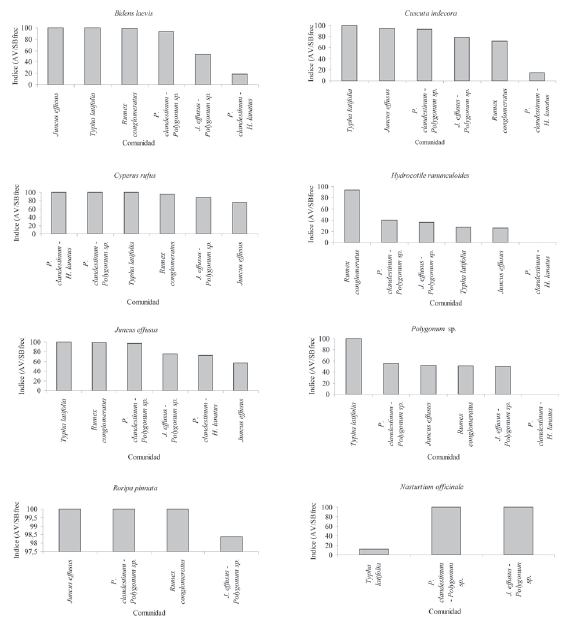
Figura 4. Persistencia de las especies más comunes del banco de semillas en el Humedal Jaboque.
Bidens laevis presenta una persistencia menor a las especies anteriormente mencionadas; por su parte Polygonum sp. e Hydrocotyle ranunculoides son las de menor persistencia en el BS, presentándose sólo en la vegetación de algunos parches (Pennisetum clandestinum-Holcus lanatus, Fig. 4).
Diversidad del BS en el humedal
La diversidad del BS para los diferentes parches, en general es baja (<2,5, Fig. 5). El valor más bajo lo presenta el parche de Juncus effusus (0,8), que exhibe la dominancia más marcada de una sola especie (Juncus effusus, Fig. 3).
Figura 5. Índices de diversidad de Shannon.
Comparación entre el Banco de Semillas y la Vegetación
Las especies compartidas entre el BS y la vegetación son muy pocas (Fig. 6), sobre todo en los parches de Pennisetum clandestinum-Holcus lanatus y Typha latifolia, en los demás parches la similitud no superó un 60% de especies en común.
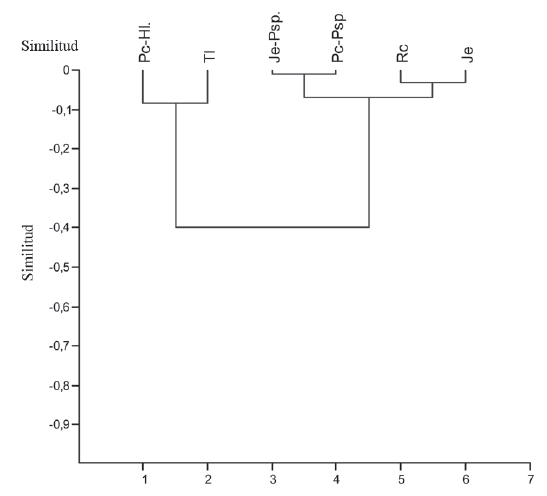
Figura 6. Similitud entre el Banco de semillas y la vegetación del humedal Jaboque (Índice de Sørensen).
Esta baja similitud entre el BS y la vegetación evidencia la existencia de tres patrones de presencia de las especies propias del humedal y oportunistas terrestres en estos parches (Anexo 2): el primero corresponde a especies exclusivas de la vegetación (Typha latifolia, Castilleja arvensis), el segundo a exclusivas del BS (Juncus tenuis y Gnaphalium americanum) y un último a especies presentes tanto en la vegetación como en el BS (Polygonum sp., Trifolium repens, Fig. 3).
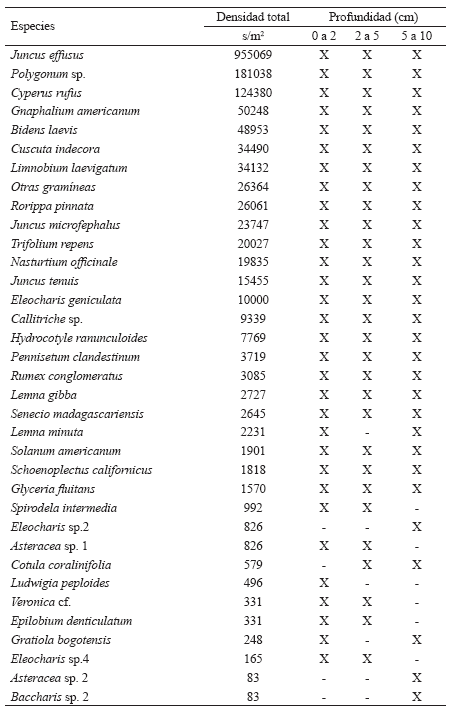
Anexo 2. Densidad de semillas en el Banco de semillas y su distribución a lo largo del perfil del suelo.
DISCUSIÓN
Estado actual del BS del humedal
En el humedal Jaboque no se ha estudiado el efecto de los tensionantes sobre la flora; se sabe para los humedales que el pastoreo intensivo elimina especies sensibles al pisoteo y al ramoneo y contribuye a la introducción de semillas de especies oportunistas terrestres (Reeves & Champion 2004); la alteración del régimen hidrológico conduce a una reducción de la diversidad y la riqueza de especies debido a la incapacidad de algunas para tolerar este cambio.
Otro tensionante es la descarga de aguas residuales domésticas e industriales (Otálora 2006) y el aporte de fertilizantes y plaguicidas, los cuales pueden conducir al enriquecimiento de nutrientes favoreciendo solamente a las especies capaces de captar con mayor eficacia dichos recursos; estos efectos sobre la composición de especies de un humedal sólo son observables a largo plazo (aproximadamente 20 años) y pueden conducir a la formación de monotipos de las especies tolerantes (Cooke et al.1990).
La composición actual de los BS del humedal Jaboque es el resultado del régimen histórico y actual de disturbios a que ha sido sometido, siendo el principal responsable de la diferencia en composición entre el BS y la vegetación actual. El régimen de disturbios, debió generar condiciones que favorecieron el ingreso de especies oportunistas e invasoras terrestres, tanto en la vegetación como en el BS, éstas a su vez eliminaron o desplazaron algunas especies propias del humedal. Esta serie de acontecimientos se reporta para otros humedales, donde los disturbios son reconocidos como generadores de cambios drásticos y/o moderados en la vegetación (Chang et al. 2001, Zedler & Kercher 2004) y para otros ecosistemas como el bosque alto andino (Montenegro 2000, Montenegro & Vargas 2005).
Actualmente el BS del humedal Jaboque presenta un estado de degradación variable a nivel de todos los parches de vegetación estudiados, revelado por la alta dominancia de unas pocas especies, la presencia de semillas de especies oportunistas en el BS de todos los parches y la baja similitud del mismo con la vegetación. Para éste último caso se sabe que en humedales conservados la similitud del BS y la vegetación tiende a ser alta (Blomqvist et al. 2003). Además, los parches de vegetación de la zona con alteración intermedia presentan una mayor degradación que se manifiesta en la reducción notable de los BS de las especies típicas del humedal. (Vargas et al. 2005).
Potencial del banco de semillas para la restauración
Algunas especies del humedal actualmente están ausentes de la vegetación y únicamente se encuentran en el BS, es el caso de Juncus tenuis, Juncus microcephalus, Eleocharis spp. También es el caso de algunas oportunistas terrestres como Gnaphalium americanum, que continúa haciendo un aporte abundante al BS, probablemente desde los alrededores del humedal, pues se encontraron sus semillas en el lodo durante el muestreo (obs. pers.).
Por otro lado, unas pocas especies poseen las estrategias para tolerar el disturbio y permanecer como elementos dominantes tanto en la vegetación como en el BS, donde otras especies han sido eliminadas; es el caso de Juncus effusus, Bidens laevis y Polygonum sp. Entre sus estrategias se encuentran la producción masiva de semillas, la formación de un BS persistente, la alta capacidad de expansión clonal; todas éstas son reconocidas para especies tolerantes al disturbio (Grime 1979). En el humedal La Conejera Bidens laevis y Polygonum sp., se reportan como especies altamente colonizadoras muy adaptables a condiciones extremas (Chisacá 2002).
Vulnerabilidad de las especies del humedal
Los bajos niveles de similitud entre la vegetación y el BS, se pueden relacionar no sólo con la historia de disturbios del humedal Jaboque sino también con la capacidad diferencial reproductiva, de dispersión y la viabilidad de las semillas, como lo mencionan Thompson & Grime (1979) para comunidades de especies herbáceas. La tolerancia diferencial de las especies al disturbio y a las condiciones generadas determina su presencia en la vegetación, en el BS o en ambos dependiendo de sus estrategias de regeneración.
Patrones de presencia de las especies
• Especies exclusivas de la vegetación: poseen estrategias como la regeneración principalmente por reproducción vegetativa, semillas que germinan casi inmediatamente luego de su dispersión, o tienen requerimientos especiales para formar BS y/o para germinar (Typha latifolia, Ludwigia peruviana y Carex luridiformis). Para el género Carex se reporta una baja viabilidad de semillas en el BS y de germinación (Van der Valk et al. 1999, Wheeler et al. 2002), sus semillas pierden viabilidad con la edad, se registra una germinación del 70% en semillas frescas, mientras a los seis meses es tan sólo del 25 al 40% (Bertilius & Boberg 2002a). Para Myriophyllum sp. la dispersión de semillas es limitada por la capacidad de flotación de las mismas; para este género es más importante la dispersión de propágulos vegetativos (Boedeltje et al. 2004).
• Especies exclusivas del BS: estas especies no se presentan en la vegetación de los parches estudiados posiblemente porque las condiciones actuales están limitando su establecimiento en los mismos (Juncus tenuis, Juncus microcephalus y Eleocharis spp.), las alteraciones del régimen hídrico pueden producir ambientes permanentemente inundados o secos, que reducen la expresión de las especies anfibias a partir del BS, debido a su adaptación a las fluctuaciones impredecibles del nivel del agua (Crosslé & Brock 2002).
• Especies presentes tanto en la vegetación como en el BS: dependen tanto del BS como de la propagación vegetativa para su mantenimiento. Se observó que Polygonum sp. logra colonizar un claro generado por la eliminación de la cobertura vegetal durante la estación seca, realizando una regeneración masiva a partir del BS y se mantiene utilizando estrategias de reproducción vegetativa. Esta combinación de estrategias parece ser la más exitosa al permitir a las especies establecerse en una gran variedad de comunidades vegetales y a la vez presentar las mayores abundancias en el BS y las mayores frecuencias de aparición (%) en la vegetación (Juncus effusus y Polygonum sp.).
La presencia de especies en el BS y en la vegetación evidencia una vulnerabilidad diferencial de las especies a la extinción local. En el humedal Jaboque se encontraron especies propias de estos ambientes con algún grado de vulnerabilidad dado por la baja frecuencia en la vegetación y la formación o no de un BS (Fig. 7).
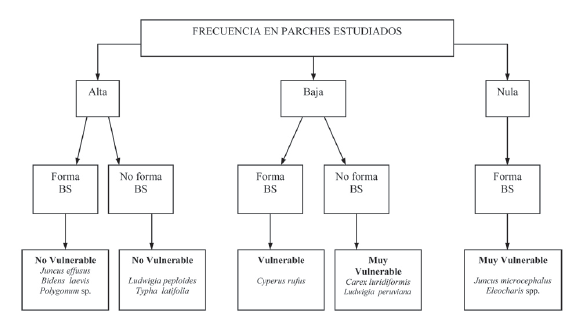
Figura 7. Vulnerabilidad de las especies propias del humedal Jaboque. BS: Banco de Semillas.
Especies muy vulnerables
Dada la importancia del BS como estrategia de regeneración de especies herbáceas en ambientes disturbados. Se consideraron como especies muy vulnerables aquellas ausentes en el BS y con baja frecuencia en la vegetación (Carex luridiformis, Ludwigia peruviana y Myriophyllum aquaticum), así como aquellas presentes en el BS, con frecuencias nulas en la vegetación (Juncus microcephalus y Eleocharis spp.) o cuya presencia esta condicionada por factores bióticos y/o abióticos temporales; en los dos casos son sensibles a la extinción local si la dinámica de la vegetación no permite su regeneración a partir del BS; por ejemplo en el caso de la especie amenazada Helenium virginicum, propia de humedales de los Estados Unidos de América, se demostró que la supervivencia de las semillas del BS es el elemento más importante para el crecimiento poblacional de ésta y que el BS actúa como amortiguador frente a la variabilidad ambiental (Adams et al. 2005).
Especies vulnerables
Se incluyen aquellas como Cyperus rufus, con baja frecuencia en la vegetación y formadoras de algún tipo de BS. A diferencia de la anterior categoría éstas poseen un potencial de regeneración a través del BS presente en diversos parches de vegetación del humedal.
Especies no vulnerables
Estas especies típicas del humedal no presentaron ningún grado de vulnerabilidad (Fig. 7) debido a que poseen mecanismos para asegurar su permanencia en el mismo, principalmente mediante la regeneración clonal (Ludwigia peploides, Typha latifolia) o combinada con la reproducción sexual, ésta ultima permite la formación de BS grandes de tipo persistente a corto o largo plazo, cuyas semillas germinan con facilidad y los individuos presentan tasas de crecimiento y biomasa altas (Ávila 2006), siendo menos susceptibles a la desaparición local (Juncus effusus, Polygonum sp. y Bidens laevis).
Las especies con altas tasas de crecimiento y reproducción tanto sexual como clonal pueden llegar a presentar características de especies invasoras, al ser las únicas capaces de sobrevivir en los hábitats severamente degradados (Nichols 1991, Newman et al. 1996, NRCS 2005a). Si bien, su crecimiento agresivo puede ser deseable para la restauración del hábitat por su capacidad de cubrir el suelo rápidamente, por ejemplo Juncus effusus (NRCS 2005b), otro estudio reporta que esta especie genera efectos fuertes en la estructura y la composición de las comunidades donde es dominante, disminuyendo la diversidad, debido a la sombra generada por su cobertura vegetal (Ervin & Wetzel 2002).
Typha latifolia, se puede considerar no vulnerable debido a sus altas tasas de crecimiento y biomasa bajo diferentes condiciones hídricas de sequía o inundación (Kercher & Zedler 2004, NRCS 2005a). Sin embargo, es una especie sensible al pastoreo por ser palatable y al disturbio, lo que restringe su hábitat a las zonas más conservadas (Bertilius & Boberg 2002b); presenta dispersión por viento (Bertilius & Boberg 2002a); aún así para otras especies de este género (Typha dominguensis) se afirma que la estrategia de regeneración consiste en dispersar sus semillas para permitir la colonización de nuevas áreas y luego expandir y mantener su población principalmente a través de reproducción vegetativa (Smith et al. 2002).
Por otra parte, las juncáceas forman BS persistentes a largo plazo; este tipo de BS a menudo pertenece a especies adaptadas a hábitats con disturbios impredecibles tales como diferentes regímenes de inundación (Bertilius & Boberg 2002a). Estudios realizados en Juncus effusus indican que aunque sus semillas germinan rápidamente en campo (7 a 10 días), sólo el 5% de las semillas dispersadas germinan antes de ser enterradas por la acumulación de sedimentos y materiales (Ervin & Wetzel 2001). Esta rápida velocidad de enterramiento puede contribuir a la formación de los BS de gran tamaño exhibidos por esta especie en el humedal Jaboque, unida seguramente a una elevada longevidad de sus semillas.
Por su parte, Polygonum punctatum crece y se reproduce rápidamente completando su ciclo de vida durante períodos cortos en los cuales el nivel del agua es bajo, confiriéndole el carácter de especie ruderal (Middleton 2002) y la dependencia de un BS persistente para su supervivencia, características que pueden conferirle a Polygonum sp. presente en el humedal Jaboque una alta frecuencia en la vegetación de la mayoría de los parches.
Tolerancia de las especies oportunistas terrestres
Los humedales, incluyendo el de Jaboque, por ser sumideros del paisaje acumulan materiales resultantes de los disturbios terrestres y del humedal (detritos, sedimentos, agua, nutrientes, sales, metales pesados y otros contaminantes), lo que facilita la invasión de especies cuando se crean claros en la vegetación y acelera el crecimiento de las plantas oportunistas (Zedler & Kercher 2004).
Al igual que para el humedal Jaboque, en otros humedales se reportan entre las invasoras pastos, graminoides, hierbas, arbustos e incluso árboles cuyo establecimiento depende de los micrositios disponibles y de la presencia de propágulos de estas invasoras (Zedler & Kercher 2004). La tolerancia de éstas y de otras especies oportunistas terrestres puede llegar a incrementar la vulnerabilidad de especies propias del humedal, si se ven favorecidas por las condiciones ambientales. Esta tolerancia permite al igual que en las especies propias de humedal la generación de tres patrones de presencia.
• Especies exclusivas de la vegetación, aunque su frecuencia de aparición fue muy baja, su presencia puede estar generada por tres factores:
1. La proximidad a una fuente de semillas, suministrada en las comunidades marginales por su cercanía al ambiente terrestre y en las comunidades ubicadas en el interior del humedal por la presencia de camellones en los cuales se encuentran estas especies (Baccharidastrum argutum).
2. El entramado de las partes vegetativas de algunas especies (Ludwigia peploides, Pennisetum clandestinum) funciona como una trampa de sedimentos generando micro-sitios para la germinación y establecimiento de especies terrestres (Castilleja arvensis y Conyza floribunda).
3. La alta incidencia de luz debido a la cobertura de poca densidad y/o altura de las especies presentes en algunas comunidades (Rumex conglomeratus – Polygonum sp.).
• Especies exclusivas del BS: estas especies actualmente no se encuentran en la vegetación posiblemente por su poca tolerancia al régimen hídrico presente en el humedal que inhibe la germinación de las semillas (Gnaphalium americanum y Cotula coralinifolia)
• Especies presentes tanto en la vegetación como en el BS: estas especies lograron adaptarse a las condiciones del humedal, tanto para establecerse en la vegetación como para presentar eventos reproductivos sexuales que les permiten aportar semillas al BS (Trifolium repens, Solanum americanum, Senecio madagascariensis, Pennisetum clandestinum y Holcus lanatus).
CONSIDERACIÓN FINAL
Teniendo en cuenta los resultados obtenidos en este estudio, si el BS del humedal Jaboque se expresa luego de un disturbio bien sea de origen natural o antrópico, su potencial para la regeneración de los parches es bastante limitado debido a su composición y al conjunto de características anteriormente descritas, principalmente porque de mantenerse las condiciones generadas por el disturbio, las especies oportunistas pueden germinar y verse favorecidas conduciendo a la competencia, el reemplazamiento y/o la eliminación de las especies de humedal como es característico en los procesos de invasión de especies (Ricklefs & Miller 2000).
AGRADECIMIENTOS
Al Profesor Orlando Rangel y a las personas que administraron el proyecto, a la Empresa de Acueducto y Alcantarillado de Bogotá (EAAB) por la financiación del mismo. A Francisco Mora por su contribución en la fase inicial de esta investigación. Al profesor Eduardo Flórez por el préstamo del invernadero del Instituto de Ciencias Naturales. A los auxiliares de campo Juan Villalba, Rafael Manjarres, Jorge Arroyo y Ricardo Beltrán, por su fundamental apoyo durante el muestreo y de manera muy especial a Ricardo Beltrán por la elaboración de la sonda para la extracción de suelo. A Ana M. Ávila por la colaboración en la fase de invernadero y a todos aquellos que de una u otra manera hicieron posible la culminación exitosa de este trabajo.
LITERATURA CITADA
1. Adams, V. M., D. M. Marsh & J. S. Knox. 2005. Importance of seed bank for viability and population monitoring in a threatened wetland herb. Biological Conservation 124: 425-436. [ Links ]
2. Ávila, Y. A. 2006. Banco de semillas germinable de ocho comunidades vegetales del Humedal “Jaboque”, Bogotá D. C. Tesis de Maestría. Universidad Nacional de Colombia. Bogotá. D.C.
3. Baker, H. G. 1989 The Ecology of Soil Seed Banks. Some Aspects of Natural History of Seed Banks. Academic Press, San Diego, California. [ Links ]
4. Baskin, C. & J. Baskin. 2001. Seeds. Ecology, Biogeography and Evolution of Dormancy and Germination. Academic Press, Nueva York. [ Links ]
5. Bertilius, K. & E. Boberg. 2002a. Wetland plants – life histories and distributional patterns Project course in Biology. Länsstyrelsen Kalmar Iän University.
6. Bertilius, K. & E. Boberg. 2002b. Adult and juvenile strategies in wetland species. A comparative study of lake Stormaren, Öland Sweden University of Kalmar, Departament of Biology and Environmental Sciences. 12-18. [ Links ]
7. Bliss, S. A. & P. H. Zedler. 1998. The germination process in vernal pools: sensitivity to environmental conditions and effects on community structure. Oecologia 113: 67-73. [ Links ]
8. Blomqvist, M. M., R. M. Bekker & P. Vos. 2003. Restoration of ditch bank plant species richness: the potential of the soil seed bank. Applied vegetation Science 6:179-188. [ Links ]
9. Boedeltje, G., J. P. Bakker, & G. N. J. Ter Heerdt. 2003. Potential role of seed banks in the development of aquatic vegetation in backwaters along navigation canals. Aquatic Botany 77: 53-69. [ Links ]
10. Boedeltje, G., J. P. Bakker, A. Ten Brinke, J. Van Groenendael y M. Soesbergen. 2004. Dispersal phenology of hydrochorous plants in relation to dischargeseed release time and buoyancy of seeds: the flood pulse concept supported. Journal of Ecology 92: 786-796. [ Links ]
11.Chang, E. R., R. L. Jefferies & T. J. Carleton. 2001. Relationship between vegetation and soil seed banks in an arctic coastal marsh. Journal of Ecology 89: 367-384. [ Links ]
12. Chisacá, M. L. 2002. Estructura y dinámica vegetal en el humedal de La Conejera (Bogotá, Cundinamarca). Trabajo de grado. Departamento de Biología. Universidad de los Andes, Bogotá, D.C. [ Links ]
13. Cooke J.G, A. Cooper & N. Clunie. 1990. Changes in the water, soil and vegetation of a wetland after a decade of receiving a sewage effluent. New Zealand Journal of Ecology. 14: 37 – 47.
14. Cronk, J. K. & M. S. Fennessy. 2001. Wetland Plants Biology and Ecology. CRC Press LLC Lewis Publishers. Nueva York. [ Links ]
15. Crosslé, K. & M. Brock. 2002. How do water regime and clipping influence wetland plant establishment from seed banks and subsequent reproduction? Aquatic Botany 74: 43-56. [ Links ]
16. DAMA, 2000. Historia de los Humedales de Bogotá con énfasis en cinco de ellos. Departamento Técnico Administrativo del Medio Ambiente. Bogotá. D.C. [ Links ]
17. DAMA, 2006. Humedales. Departamento Técnico Administrativo del Medio Ambiente. Bogotá. http://www.dama.gov.co/dama.php [ Links ]
18. EAAB & Conservación Internacional-Colombia. 2000. “Síntesis del estado actual de los Humedales Bogotanos”. Recuperación de los Humedales de la Sabana de Bogotá. Alternativas hacia su viabilidad Ecológica y Social. Bogotá. D.C.
19. EAAB. 2000. Recuperación de los Humedales de la Sabana de Bogotá”. Conservación Internacional, Bogotá, D.C.
20. Ervin, G. N. & R. G. Wetzel. 2001. Seed fall and seed germination of needlerush Juncus effusus L. Aquatic Botany 71: 233-237. [ Links ]
21. Ervin, G. N. & R. G. Wetzel. 2002. Influence of a dominant macrophyte, Juncus effusus, on wetland plant species richness, diversity, and community composition. Oecologia 130: 626–636.
22. Fenner, M. 1995. Ecology of seed banks. Seed dormancy and germination. 507-527. [ Links ]
23. Galatowitsch, S. & A. G. Van der Valk. 1996. The vegetation of restored prairie wetlands. Ecology App l 6:102. [ Links ]
24. Galinato, M. I., & A. G. Van der Valk. 1986. Seed germination traits of annuals and emergents recruited during drawdowns in the Delta Marsh, Manitoba. Can. Aquat. Bot. 26: 89–102.
25. Garwood, N. C. 1989. Tropical soil seed bank: a review. En: M. A. Leck, V. Parker y R. L. Simpson (Eds). Ecology of soil seed banks. Academic Press INC, Nueva York. [ Links ]
26. Grime, J. 1979. Plant Strategies and Vegetation Processes. En: Thompson, D (Ed). The Functional Ecology of Seed Banks, Nueva York. [ Links ]
27. Harper, J. L. 1977. Population biology of plants. Academic Press. Londres. [ Links ]
28. Hernández, J., J.O. Rangel-Ch. & A. Granés-M. 2005. La vegetación y el impacto antrópico del humedal de Jaboque. Universidad Nacional de Colombia. Instituto de Ciencias Naturales. Bogotá, D.C. [ Links ]
29. Hölzel, N. & A. Otte. 2004. Assessing soil seed bank persistence in flood-meadows: The search for reliable traits. Journal of Vegetation Science 15: 93-100. [ Links ]
30. Kercher, S. M. & J. Zedler. 2004. Flood tolerance in wetland angiosperms: a comparison of invasive and noninvasive species. Aquatic Botany 80: 89-102. [ Links ]
31. Leck, M. A. & R. L. Simpson. 1995. Ten year seed bank and vegetation dynamics of a tidal freshwater marsh. Am. J. Bot. 82: 1547–1557.
32. Louda, S. M. 1989. Predation in the dynamics of seed regeneration. En: M. A. Leck, V. Parker y R. L. Simpson (Eds). Ecology of soil seed banks Academic Press. INC, p 25-51. San Diego, California. [ Links ]
33. Matteucci, S. & A, Colma. 1982. Metodología para el estudio de la vegetación. Secretaría general de la Organización de los Estados Americanos. Washington, D.C. [ Links ]
34. Middleton, B. 2002. The flood pulse concept in wetland restoration. En: B. Middleton (Ed). Restoration the natural hydrological balance. John Wiley & Sons, Inc. Nueva York. [ Links ]
35. Montenegro, A. L. 2000. Estrategias de dispersión y regeneración por bancos de semillas en dos comunidades de bosque altoandino (Embalse de San Rafael). Trabajo de grado. Departamento de Biología. Universidad Nacional de Colombia, Bogotá, D.C. [ Links ]
36.Montenegro, A. L. & O. Vargas. 2005. Estrategias de regeneración del banco de semillas en una comunidad de bosque alto andino secundario. En: A. Bonilla (Ed). Estrategias adaptativas de plantas del bosque alto andino en la Cordillera Oriental de Colombia. p. 227-246. Bogotá, D.C. [ Links ]
37. NRCS (Natural Resources Conservation Service). 2005a. Soft rush Juncus effusus L. Plant Guide. United States Department of Agriculture. http://plants.usda.gov/plantguide/pdf/cs_juef.pdf [ Links ]
38. NRCS (Natural Resources Conservation Service). 2005b. Broad-leaved cattail Typha latifolia L. Plant Guide. United States Department of Agriculture. http://plants.usda.gov/plantguide/pdf/cs_tyla.pdf [ Links ]
39. Newman, S., J. B. Grace & J. W. Koebel. 1996. Effects of nutrients and hydroperiod on Typha, Cladium and Eleocharis: Implications for everglades restoration. Ecology appl. 6:774-783. [ Links ]
40. Nichols, S. A. 1991. The interaction between biology and the management of aquatic macrophytes. Aquatic Botany 41: 225-252. [ Links ]
41. Otálora, A. 2006. Influencia de las amenazas sobre la comunidad de aves silvestres residentes en el humedal Jaboque. Tesis de Maestría. Universidad Nacional de Colombia. Bogotá, D.C. [ Links ]
42. Parker, V., R. Simpson & M. Leck. 1989. Pattern and process in the dynamics of seed banks. En: M. A. Leck, V. Parker y R. L. Simpson (Eds). Ecology of soil seed banks. Academic Press INC, p 367-384. San Diego, California. [ Links ]
43. Peterson, J. E. & A. H, Baldwin. 2004. Seedling emergence from seed banks of tidal freshwater wetlands: response to inundation and sedimentation Aquat. Bot. 78:243-254. [ Links ]
44. Pickett, S. T. & M. J. McDonnell. 1989. Seed bank dynamics in temperate deciduous forest. En: M. A. Leck, V. Parker y R. L. Simpson (Eds). Ecology of soil seed banks. Academic Press INC, p 123-146. San Diego, California. [ Links ]
45. Reeves, P. N. & P. D. Champion. 2004. Effects of livestock grazing on wetland: Literature Review. National Institute of water & atmospheric Research Ltd. (NIWA). Hamilton. [ Links ]
46. Ricklefs, R. E. & G. L. Miller. 2000. Ecology. Ed. W. H. Freeman and Company. Fourth Edition. Nueva York. [ Links ]
47. Simpson, R., M. Leck & V. Parker. 1989. Seed banks: General concepts and methodological issues. En: M. A. Leck, V. Parker y R. L. Simpson (Eds). Ecology of soil seed banks. Academic Press INC, p 3-8. San Diego, California. [ Links ]
48. Smith, S. M., P. McCormick, J. Leeds & P. Garrett. 2002. Constraints of seed bank species composition and water depth for restoring vegetation in the Florida Everglades, USA. Restoration Ecology 10: 138-145. [ Links ]
49. Thompson, D. & J. P. Grime. 1979. Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats. Journal of Ecology 67: 893-921. [ Links ]
50. Thompson, K., J. P. Bakker & R. M. Bekker. 1997. The soil seed banks of North West Europe: methodology, density and longevity. Cambridge University Press, Cambridge. [ Links ]
51. Van der Valk, A. G. 1981. Succession in wetlands: A Gleasonian approach. Ecology 62: 688-696. [ Links ]
52. Van der Valk, A. & Pederson. 1989. Seed banks and the management and restoration of natural vegetation. En: M. A. Leck, V. Parker y R. L. Simpson (Eds). Ecology of soil seed banks. Academic Press INC, p. 329-346. San Diego, California. [ Links ]
53. Van der Valk, A. G., T. L. Bremholm & E. Gordon. 1999. The restoration of sedge meadows: seed viability, seed germination requirements and seedling growth of Carex species. Wetlands 19:759-764. [ Links ]
54. Vargas, O., M. A. Bonilla, Y. A. Ávila, A. L. Montegegro & H. A. Chaparro. 2005. Evaluación del banco de semillas germinable como estrategia de restauración del Humedal Jaboque. Empresa de Acueducto y Alcantarillado de Bogotá-Universidad Nacional de Colombia. Bogotá, D.C. [ Links ]
55. Vavrek, M. C. & W. J. Campbell. 2005. Contribution of seed banks to freshwater wetland vegetation recovery. http://www.epa.gov/oilspill/pdfs/vavrekpaper.pdf [ Links ]
56. Walck, J. L., M. Baskin, C. C. Baskin & S. W. Francis.1996. Sandstone rockhouses of the eastern United States, with particular reference to the ecology and evolution of the endemic plant taxa. Botanical Review 62: 311-362. [ Links ]
57. Warwick, N. W. & M. A. Brock. 2003. Plant reproduction in temporary wetlands: The effects of seasonal timing, depth and duration of flooding. Aquat. Bot. 77: 153-167. [ Links ]
58. Wheeler, B. D., R. P. Money & S. C. Shaw. 2002. Freshwater wetlands. En: Perrow, M. R. y A. J. Davy (eds). Handbook of Ecological Restoration. II. Restoration in practice. Cambridge University Press, Cambridge. p. 325-354. [ Links ]
59. Zedler, J. B. & S. Kercher. 2004. Causes and consequences of invasive plants in wetlands: opportunities, opportunists and outcomes. Critical Reviews in Pant Sciences 23:431-452. [ Links ]
Recibido: 20/01/2006
Aceptado: 05/09/2006

