










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.29 no.1 Bogotá Jan./June 2007
LOS MUSGOS (BRYOPHYTA) DE LA REGIÓN DE SANTA MARÍA-BOYACÁ (COLOMBIA)
The Mosses (Bryophyta) of the Region of Santa María-Boyacá (Colombia)
KARINA AVENDAÑO-TORRES
JAIME AGUIRRE-C.
Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Apartado 7495, Bogotá, D.C., Colombia. kavendanot@unal.edu.co; jaguirrec@unal.edu.co
RESUMEN
En la región de Santa María (Boyacá) se registraron 68 especies de musgos, pertenecientes a 45 géneros y 21 familias; 22 taxones son nuevos registros para el departamento. Las familias más ricas en géneros y especies fueron: Hypnaceae (8 géneros - 8 especies), Meteoriaceae (6-7), Pilotrichaceae (4-7) y Fissidentaceae (1-6). Los géneros más ricos en especies fueron: Fissidens (6) y Syrrhopodon (3). La composición por región de vida fue para la tropical de 63 especies en 45 géneros y 21 familias y para la subandina 22 especies de 19 géneros y 15 familias. Las dos regiones comparten 17 especies, en 17 géneros y 14 familias. Los hábitats (sustratos) más diversos fueron: epífitos (16 especies), epilíticos (8) y epilítico-terrestres (8), doce especies se presentaron en todos los sustratos. Las formas de vida con mayor representación fueron: cespitosas cortas (24) y felpas lisas (17).
Palabras clave. Briófitos, musgos, Santa María, Boyacá, formas de crecimiento.
ABSTRACT
In Santa María (Boyacá-Colombia) 68 species belonging to 45 genera and 21 families were recorded, of which 22 species were first records for the department. The richest families in genera/species were Hypnaceae (8 genera – 8 species), Meteoriaceae (6-7), Pilotrichaceae (4-7) and Fissidentaceae (1-6), while the most rich genera were Fissidens (6) and Syrrhopodon (3). In the Tropical regions 63 species belonging to 45 genera and 21 families were registered, and in the subandean regions 22 species of 19 genera and 15 families were found. Both regions share 17 species of 17 genera and 14 families. The substrates with greatest number of species were epiphytes (16), epilithic (8) and epilithic-terrestrial (8). Twelve species were found in all substrates. The commonest life forms were short turfs (24 species) and smooth mats (17).
Key Words. Bryophytes, mosses, Santa María, Boyacá, growth-forms.
INTRODUCCIÓN
Los briófitos se establecen con preferencia en la región de vida andina en donde desempeñan un papel muy importante en los procesos relacionados con la economía hídrica de la región cordillerana, interceptan la lluvia, absorben y retienen agua, previenen su escape por escorrentía y en algunas regiones, dada su gran biomasa detienen, la erosión (Churchill & Linares 1995).
En Colombia los estudios sobre la composición, la ecología y la vegetación de la brioflora (musgos y hepáticas) en diferentes regiones geográficas se ha incrementado en los últimos años, entre éstos podemos mencionar a Van Reenen & Gradstein (1983), Wolf (1993), Linares (1986), Rudas & Aguirre (1990), Orrego (2000), Ruiz (2001) y Aguirre & Ruiz (2001). La mayoría de las contribuciones se han centrado en las regiones de vida andina y paramuna, mientras que para las tierras bajas su número es menor.
Santos (2002) proporcionó para el departamento de Boyacá datos importantes sobre la composición florística de los musgos de la Serranía de las Quinchas (Magdalena Medio) Ruiz et al. (2002) sobre aspectos taxonómico-ecológicos de Sphagnum y Lagos et al. (2002) sobre los briófitos reófilos del Cerro de Mamapacha, Chinavita. Para la región de estudio, en cuanto a este grupo de plantas, es una de las primeras contribuciones.
Este trabajo es una contribución sobre el conocimiento de los musgos de tierras bajas, se desarrolló en un área que incluye los rangos altitudinales de 500 a 1200 m, que corresponden a la región de vida tropical (0-1000 m) y a la parte baja de la región de vida subandina (1000-2350 m) (Rangel et al. 1995). El propósito fundamental fue el de determinar la riqueza de especies por género y por familia en la región y proporcionar aportes a la ecología de las especies registradas, estableciendo la preferencia de hábitat (sustrato) y formas de vida más comunes en la zona.
MATERIALES Y MÉTODOS
Se estudiaron las colecciones realizadas como parte de la fase de campo de la asignatura Introducción a la Sistemática Vegetal en los años 2000, 2001 y 2005, se visitaron las siguientes zonas: Vereda Caño Negro, camino hacia la quebrada Catalina; vía a Mámbita, desvío a la carbonera; vía a San Luis de Gaceno, hacienda Cachipay; vía de Santa María a Mámbita; carretera a Mámbita (orillas del río Mapa); bosque “La Almenara” por la carretera Santa María a San Luis de Gaceno; vereda La Esmeralda, por la carretera entre Santa María y Guateque; carretera entre Santa María y San Luis de Gaceno por la margen derecha del río Lenguapá, que se encuentran entre 500 m – 860 m y en la Vereda La Almenara a 1200 m. Las muestras se colectaron bajo la numeración de Jaime Aguirre-C., Jaime Uribe-M y Karina Avendaño-Torres y se encuentran depositadas en el Herbario Nacional de Colombia (COL).
El material se determinó utilizando, entre otras contribuciones, a Buck (1998), Churchill & Linares (1995), Frahm (1991), Geir (1980), Ochi (1980, 1981), Reese (1961, 1977, 1978), Salazar (1991) y con la consulta de ejemplares depositados en el Herbario Nacional Colombiano (COL).
Se tomó la información sobre el sustrato (suelo, base de troncos, rocas, troncos en descomposición, hojas, hojarasca) clasificándolos en epífitos, epilíticos, terrestres y materia orgánica en descomposición. Las formas de vida se estudiaron de acuerdo con Mägdefrau (1952) y Gimingham & Birse (1957) y básicamente se tomaron las siguientes: Cespitosas altas (Ca), Cespitosas cortas (Cc), Felpas lisas (Fl), Felpas rugosas (Fr), Tramas (T), Dendroides (D), Cojines (Coj) y Formas péndulas (P).
ÁREA DE ESTUDIO
El área de estudio se localiza en el municipio de Santa María, al sur oriente del departamento de Boyacá, a los 4° 51’ 48’’de latitud norte y 73° 16´ 04’’ de latitud oeste, entre los 600m y 1200m de altitud. Este municipio limita por el norte con los municipios de Macanal y Campohermoso, por el oriente con San Luis de Gaceno, por el sur con el departamento de Cundinamarca y por el occidente con Chivor y Maconal; se distinguen dos unidades fisiográficas, la primera es de relieve montañoso, donde sobresalen las cuchillas de Guaneque, Negro y Calichona, la segunda de relieve bajo, parte de la faja del piedemonte llanero (IGAC 1996).
La temperatura media es de 24ºC y una precipitación anual de 4694,9 mm. De diciembre a febrero se presenta el período menos húmedo, el de mayor precipitación corresponde a los meses de abril a octubre. Enero es el mes más seco con 63 mm y julio el más lluvioso con 674,8 mm (IDEAM, estación pluviométrica de Santa María). En región la pluviosidad y condensación es alta, hasta el punto de que los meses menos lluviosos tiene 4 ó 5 días de precipitación (IGAC 1977).
Los suelos se presentan en tres asociaciones: la Asociación Santa Teresa que presenta un relieve plano y en algunos sectores ondulado y quebrado, erosión ligera a moderada, suelos bien drenados, permeabilidad rápida, regular a baja retención de humedad; la Asociación Lengupá con un relieve plano a inclinado, suelos moderadamente profundos, bien drenados, de rápida a regular permeabilidad, con una baja a regular retención de agua y la Asociación Santa María de relieve quebrado a escarpado, suelos moderadamente profundos a superficiales, con erosión moderada, bien a excesivamente drenados, con permeabilidad rápida y regular retención de humedad (IGAC 1977).
RESULTADOS Y DISCUSIÓN
Composición florística
Se registraron 68 especies en 45 géneros y 21 familias, que corresponden al 29,2% del total de las especies reportadas para Boyacá (233) (Churchill & Linares 1995). Las familias más ricas en géneros y especies fueron: Hypnaceae (8 géneros -8 especies), Meteoriaceae (6-7), Pilotrichaceae (4-7), Neckeraceae (3-3), Dicranaceae (3-3) y Fissidentaceae (1-6) (Figura 1) y los géneros: Fissidens (6), Syrrhopodon (4), Bryum (3), Campylopus (3) y Callicostella (3). Veintidós (22) taxones corresponden a nuevos reportes para el departamento.
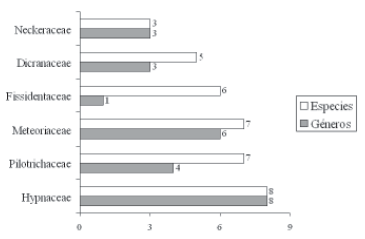
Figura 1. Familias de musgos con el mayor número de géneros y especies.
Composición por región de vida
Región de vida tropical (500m-860m): Se presentaron 63 especies en 45 géneros y 21 familias (Anexo 1). Las familias más ricas fueron: Hypnaceae (8-8), Pilotrichaceae (4-7), Meteoriaceae (6-7) y Calymperaceae (2-5) y los géneros Syrrhopodon (4), Fissidens (4) y Bryum (3). Para esta región de vida se estudiaron 274 colecciones, de las cuales las especies más frecuentes fueron Vesicularia vesicularis (5,84%), Neckeropsis undulata (5,84%), Racopilum tomemtosum (5,47%) y Sematophyllum subpinnatum (5,11%).
Aguirre (2006) registró para la región de vida tropical del sistema cordillerano 213 especies de 101 géneros y 38 familias, y para la región de vida tropical de la cordillera oriental 78 especies de 56 géneros y 31 familias, lo que indica que la zona de estudio se presenta el 30% de las especies, 45% de los géneros y el 55% de las familias con relación al sistema cordillerano, y 81% de las especies, el 80% de los géneros y el 68% de las familias con relación a la cordillera oriental.
Región de vida subandina (1200 m): Con 22 especies de 19 géneros y 15 familias (Anexo 1). Las familias con mayor número de géneros y especies fueron: Hypnaceae (3-3), Fissidentaceae (1-3), Pilotrichaceae (2-2) y Orthotrichaceae (2-2) y los géneros Fissidens (3) y Campylopus (2). Las especies más frecuentes fueron: Philonotis uncinata (14,81%), Racopilum tomemtosum, Sematophyllum galipense, Syrrhopodon prolifer, Cyclodictyon albicans y Leucobryum martianun todos con la misma frecuencia (9,26%), de un total de 55 colecciones estudiadas para está región de vida.
Para la región de vida subandina del sistema cordillerano Aguirre (2006) encontró 562 especies de 194 géneros y 57 familias, y para la región de vida subandina de la cordillera oriental 355 especies de 142 géneros y 49 familias, lo que indica que la zona de estudio se presenta el 4% de las especies, 9.8% de los géneros y el 26,3% de las familias con relación al sistema cordillerano, y 6,2% de las especies, el 13,4% de los géneros y el 30,6% de las familias con relación a la cordillera oriental. Los porcentajes de similitud son menores debido a la baja intensidad de muestreo en esta región de vida en el área de estudio.
Dada la diferencia en la intensidad de muestreo, de las nueve zonas estudiadas sólo una (1) se localiza en la región de vida subandina, una comparación aproximada de la composición florística entre las dos regiones muestra que la región de vida tropical presentó el mayor número de especies, géneros y familias (Figura 2), contrario a lo que comúnmente se presenta en el resto del país, en donde la riqueza de especies aumenta en la medida en que se asciende en el gradiente altitudinal.
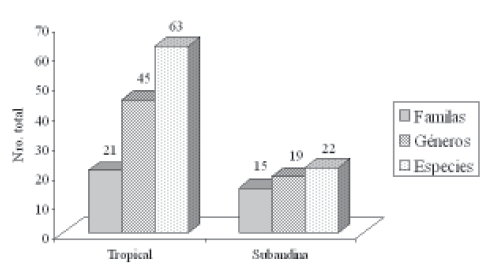
Figura 2. Número de familias, géneros y especies presentes en cada región de vida.
Relaciones de semejanza
Contigüidad (semejanzas entre regiones de vida): Tropical – subandina. Estas dos regiones de vida comparten 17 especies de 17 géneros y 14 familias. Las familias con mejor representación fueron: Hypnaceae (3-3) y Pilotrichaceae (2-2). Philonotis uncinata, Rhyncostegium serrulatum, Rhodobryum beyrichianum, Syrrhopodoon prolifer, Fissidens mollis, Ectroopothecium leptochaeton, Mittenothamnium reptans, Vesicularia vesicularis, Leucobryum martianum, Macromitrium punctatum, Hyophila involuta, Cyclodictyon albicans, Lepidopilum scabrisetum, Racopilum toomentosum, Sematophyllum galipense, Sphagnum subsecundum y Thuidium urceolatum.
Especies restringidas
Región de vida tropical. Se encontraron 46 especies en 35 géneros y 19 familias (Tabla 1). Las familias con mayor representación fueron: Meteoriaceae (7-6) y Hypnaceae (5-5).
Tabla 1. Especies de musgos restringidas a la región de vida tropical.

Región de vida subandina. Se presentaron cinco especies: Campylopus concolor, C. flexuosus, Fissidens crispus, F. prionodes y Groutiella husnotii.
Selectividad de sustrato
Se encontrarón 16 especies epífitas, 10 epilíticas y 4 terrestres. Un buen número de especies se presentaron en más de un sustrato, los de mayor número de especies fueron: epilítico-terrestres (8), epífito-materia orgánica en descomposición (5) y epífito-epilítico-terrestres (4). Doce especies se presentaron en todos los sustratos (Figura 3).
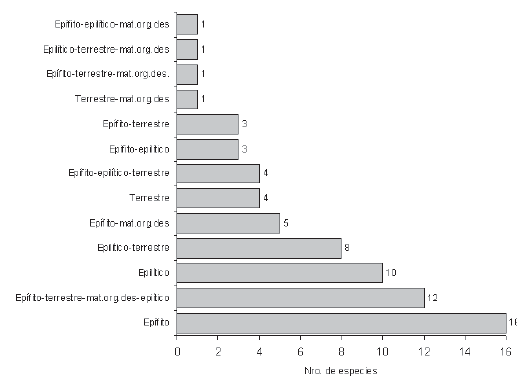
Figura 3. Número de especies por tipos de sustrato.
Las familias mejor representadas en los epifitos fueron: Calymperaceae (3 especies), Meteoriaceae (2), Dicranaceae (2) y Pilotrichaceae (2). En las epilíticas: Bryaceae (3), Fissidentaceae (2) e Hypnaceae (2). En las terrestres con una especie: Bryaceae, Dicranaceae, Orthotrichaceae y Fissidentaceae. Con elementos que crecen en más de un sustrato, epílitico-terrestre: Pilotrichaceae (3), Fissidentaceae (2). En las epífito-materia orgánica en descomposición: Meteoriaceae (2). En las epifito-epilítico-terrestres: Hypnaceae (2), Pilotrichaceaae (2). En todos los sustratos Hypnaceae (2), Sematophyllaceae (2) y Neckeraceae (2).
Papillaria nigrescens (12,5%) fue la especie epifita más frecuente. Entre las epilíticas las más importantes fueron Fissidens mollis (4,35%) y Campylopus sp1 (2,61%). En las terrestres: Bryum sp1 (40%). Las epilítico-terrestres más frecuentes fueron Cyclodictyon albicans (32,3%), Barbula indica (12,9%) y Callicostella rivularis (12,9%). Entre las epífito-materia orgánica en descomposición Octoblepharum albidum (33,3 %). Dentro de las especies epífito-epílitico-terrestres Vesicularia vesicularis (39%) y Philonotis uncinata (37%) fueron las más comunes. De las que crecen en todos los sustratos las más importantes fueron, Racopilum tomemtosum (14,8%), Neckeropsis undulata (11,9%), Sematophyllum galipense (11,9%) y Sematophyllum subpinnatum (10,4%).
Región de vida tropical (500m-860 m). Los sustratos en los que se encontraron mayor número de especies fueron epífitos (14 especies) y epilíticos (8 especies). En los epífitos las familias con mayor representación fueron: Calymperaceae (1-3), Meteoriaceae (2-2) y Pilotrichaceae (2-2) y el género Syrrhopodon (3). En los epilíticos: Hypnaceae (2-2) y Bryaceae (1-2) y el género Bryum (2). En más de un sustrato, en las epilítico-terrestres Callicostella rivularis, Barbula indica, Fissidens elegans, Octoblepharum pulvinatum y Callicostella columbica; en las epífito-maordes Octoblepharum albidum, Orthostichella pentasticha, Schoenobryum concavifolium, Entodon macropodus y Squamidium nigricans; en las epífito-epilítica Callicostella pallida, Leskeodon andicola y Meteoridium remotifolium; una sola especies epífito-terrestre-materia orgánica en descomposición Zelometeorim patulum. Seis (6) especies se presentaron en todos los sustratos: Neckeropsis undulata, Sematophyllum subpinnatum, Rhynchostegium scariosum, Thuidium tomentosum, Rhacopilopsis trinitensis y Porotrichum longirostre.
Región de vida subandina (1200 m): Se presento una especies epífita, Campylpus flexuosus; dos epilíticas: Fissidens mollis y Rhodobryum beyrichianum y dos terrestres: Fissidens prionodes y Groutiella husnotii. En más de un sustrato, epilítico-terrestre con tres especies: Campylopus concolor, Fissidens crispus y Cyclodictyon albicans y epífito-epilítico-terrestre con cinco: Lepidopilum scabrisetum, Philonotis uncinata, Rhynchostegium serrulatum y Vesicularia vesicularis. En todos los sustratos (6): Ectropothecium leptochaeton, Leucobryum martianun, Racopilum tomemtosum, Sematophyllum galipense, Syrrhopodon prolifer y Thuidium urceolatum.
Formas de vida
Se presentaron las siguientes formas de vida: cespitosas cortas (Cc, 24 especies), felpas lisas (Fl, 17) y felpas rugosas (Fr, 9). Las menos frecuentes fueron dendroides (D, 3), Cojines (Coj, 2) y trama (T, 2) (Figura 4).
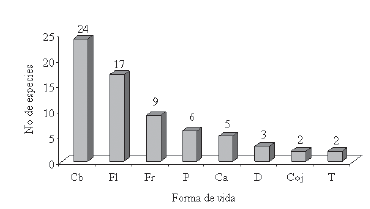
Figura 4. Distribución de las especies según las formas de vida.
Con forma de vida cespitosa baja las familias mejor representadas fueron: Dicranaceae (3-3), Calymperaceae (2-5) y Fissidetaceae (1-6). Con forma de vida felpas lisas Hypnaceae (6-6) y Pilotrichaceae (2-4) fueron las familias con mayor número de especies. Pilotrichaceae (3-3) y Sematophyllaceae (1-2) fueron las familias con mayor representación de especies con forma de crecimiento felpas rugosas.
Región de vida tropical: Se presentaron las formas de vida cespitosas cortas (Cc, 20 especies) y felpas lisas (Fl, 17), le siguen en orden de importancia felpas rugosas (Fr, 9), formas péndulas (P, 6) y cespitosas altas (Ca, 4). Las formas dendroides (D, 3), trama (T, 2) y cojines (Coj, 2) fueron las menos frecuentes en esta región de vida.
Región de vida subandina: La formas de vida con el mayor número de especie fueron: cespitosas cortas (Cc, 9 especies) y felpas lisas (Fl, 4), seguidas por felpas rugosas (Fr, 3) y cespitosas altas (Ca, 3). Las menos frecuentes fueron: trama (T, 1), cojines (Coj, 1) dendroides (D, 1) y formas péndulas (P, 0).
El número de especies en cada forma de vida es mayor para la región de vida tropical (Figura 5), probablemente por la mayor cantidad de especies registradas para está región debido a la intensidad de muestreo. Las formas de crecimiento cespitosas cortas y felpas lisas fueron las dominantes en las dos regiones.
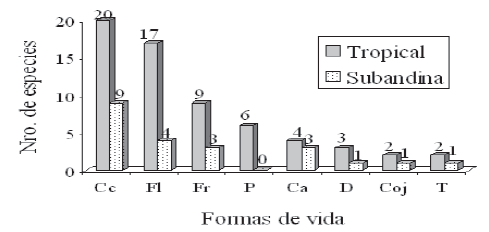
Figura 5. Distribución de las especies en las formas de vida en cada región de vida.
CONSIDERACIONES FINALES
Se encontraron 68 especies de musgos. Las familias más diversas fueron Hypnaceae (8), Meteoriaceae (7) y Pilotrichaceae (7) y los géneros Fissidens (6) y Syrrhopodon (3). Santos (2002) registró entre las familias más ricas en géneros y especies para las regiones de vida tropical y subandina de la Serranía de las Quinchas las mencionadas anteriormente, al igual que Fissidens y Syrrhopodon como los géneros más diversos.
En la región de vida tropical se encontraron 63 especies y en la subandina 22. Las familias más diversas fueron Hypnaceae y Pilotrichaceae y el género Fissidens. El menor número de especies en la región de vida subandina indica que es necesario incrementar las colecciones para una adecuada estimación y comparación sobre la riqueza de especies.
Los valores encontrados comparados con los registrados para Colombia en regiones geográficas diferentes para las mismas regiones de vida, como por Aguirre & Ruiz (2001) en la Serranía de Perijá (Cesar), Ruiz (2001) en el corregimiento de Tarapacá y Santos (2002) en la Serranía de las Quinchas (Magdalena Medio) indican que se mantiene el patrón de baja diversidad en los bosques de tierras bajas. Aguirre (2006), sintetizó la información sobre los musgos de Colombia y para el sistema cordillera encontró, que en general, en el gradiente altitudinal la riqueza de especies es menor a bajas altitudes.
Existe una preferencia por el hábitat epífito. Según Richards (1982) la mayoría de los briófitos del bosque lluvioso tropical de zonas bajas son epífitos, la gruesa capa de hojas que permanece sobre el suelo impide el establecimiento de briófitos sobre éste. De acuerdo con Pócs (1982), los briofitos forman en el bosque lluvioso tropical un sistema que evolucionó hacia una eficiente interceptación de agua, haciéndose innecesario el mantener contacto físico o químico directo con el suelo. Si bien, la luz es un factor esencial para el crecimiento de los epífitos, su demanda no fue el eje de su evolución. Frahm (2003) resalta que la alta cantidad de briófitos epífitos es un carácter especial de los bosques lluviosos tropicales como un resultado directo de la alta humedad y periodos más largos de luz.
La forma de vida cespitosa cortas (24 especies) y felpas rugosas (9) fueron las dominantes debido a las condiciones ambientales de alta humedad y alta intensidad lumínica presentes en gran parte de los sectores donde se realizo el estudio. Birse (1957), Gimingham & Birse (1957) y Magdefrau (1952) postularon que las formas de vida de los briófitos se ven influenciadas por las condiciones ambientales, en particular por la humedad relativa y la intensidad lumínica, en condiciones de alta humedad – alta intensidad lumínica el crecimiento tiende a ser vertical favoreciendo el desarrollo de las formas de vida cespitosas altas y cortas y felpas rugosas, mientras que las formas postradas (felpas lisas, tramas y algunas formas dendroides) están relacionadas con condiciones de baja humedad –alta intensidad de luz.
Las felpas lisas siguen en dominancia debido a que gran cantidad de especimenes fueron herborizados en sitos abiertos (senderos o bordes de carretera) en donde se presentan condiciones de alta intensidad lumínica y baja humedad relativa que les son favorables. Esta observación coincide con lo encontrado por Acebey et al. (2003), trabajo en el que se evidenció que las formas de vida felpas lisas se presentaban en un buen porcentaje en zonas abiertas, lo que podría reflejar las condiciones cálidas y secas del microclima de estas partes.
En investigaciones realizadas sobre las formas de vida de los briófitos en bosques de tierras bajas se menciona la forma de vida felpa lisa como la más importante y frecuentes (Cornelissen & Steege 1989, Ruiz 2001). Lo anterior no se evidencia en el presente estudio debido a que en las contribuciones mencionadas realizaron muestreos en toda la longitud de los forofitos, incluyendo el dosel donde se presentan las condiciones ambientales propias para el desarrollo de esta forma de vida.
La baja representación de las formas cespitosas altas, cojines y dendroides en la zona, se debe a que estos tipos de crecimiento están relacionadas con hábitats húmedos y sombríos, condiciones que se presenta en escasos lugares en el área de estudio.
Las dos regiones de vida presentan en común 17 especies, de 17 géneros y 14 familias, lo cual muestra que comparten el 25% de las especies, el 37,8% de los géneros y el 66,7% de las familias. Las especies compartidas más frecuentes fueron: Racopilun tomentosun, Vesicularia vesicularis, Philonotis uncinata y Sematophyllun galipense. En la región de vida tropical la mayoría de las especies fueron epifitas mientras que en la región de vida subandina no se presento preferencia de hábitat. En ambas, las formas de vida dominantes fueron cespitosas cortas y felpas lisas.
AGRADECIMIENTOS
Al Instituto de Ciencias Naturales y al Departamento de Biología de la Universidad Nacional de Colombia, al Herbario Nacional Colombiano (COL) por las facilidades brindadas. A Jaime Aguirre-C., por su orientación y apoyo para el desarrollo del presente trabajo. A Édgar Linares y a Maribel Pinzón por su colaboración en la determinación de algunas de las especies.
LITERATURA CITADA
1. Acebey, A., S. R. Gradstein & T. Kromer. 2003. Species richness and habitat diversification of bryophytes in submontane rain forest and fallows of Bolivia. Journal of Tropical Ecology 19:9-18. [ Links ]
2. Aguirre, J. 2006. Diversidad de los musgos (Briophita) y líquenes de Colombia-Una evaluación con propósitos de conservación. Tesis doctoral. Universidad Nacional de Colombia. Facultad de Ciencias. Instituto de Ciencias Naturales, Bogotá. [ Links ]
3. Aguirre, J. & C. Ruiz, 2001. Composición florística de la brioflora de la Serranía del Perijá (Cesar-Colombia): distribución y ecología. Caldasia 23(1):181-201. [ Links ]
4. Birse, E. M. 1957. Ecological studies on growth-form in bryophytes. II Experimental studies on growth-form in mosses. Journal of Ecology 45:721-733. [ Links ]
5. Buck, W.R. 1998. Pleurocarpous mosses of the West Indies. The New York Botanical Garden. Bronx, Nueva York. [ Links ]
6. Churchill, S.P. & E.L. Linares. 1995. Prodomus Bryologiae Novo Granatensis. Introducción a la flora de musgos de Colombia. Biblioteca José Jerónimo Triana. Tomos I y II. Instituto de Ciencias Naturales. Facultad de Ciencias. Universidad Nacional de Colombia, Bogotá. [ Links ]
7. Cornelissen, J.H.C. & H.T. Steege. 1989. Distribution and ecology of epiphytic bryophytes and lichens in dry evergreen forest of Guyana. Journal of Tropical Ecology 5:131-150. [ Links ]
8. Frahm, J. P. 1991. Dicranaceae. Campylopodioidae, Paraleucobryoideae. Flora Neotropica Monograph 54:1-128. [ Links ]
9. Geir, L.J. 1980. A preliminary study of the Thuidiaceae (musci) of Latin America. Journal of Bryology 11:253-309. [ Links ]
10. Gimingham, C.H. & E.M. Birse. 1957. Ecological studies on growth-form in bryophytes. I. Correlations between growth-form and habitat. Journal of Ecology 45: 533-545. [ Links ]
11. IDEAM, Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia. Estación pluviométrica de Santa María, Boyacá, Colombia. [ Links ]
12. IGAC.1977. Estudio general de suelos del Valle de Tenza, región de Lengupá y municipio de Pesca, departamento de Boyacá. Instituto Geográfico Agustín Codazzi, Bogotá. [ Links ]
13. IGAC. (eds.). 1996. Diccionario Geográfico de Colombia. Tomo IV. Tercera edición. Bogotá. [ Links ]
14. Lagos, M., F. Sáenz & M.E. Morales. 2002. Briófitos reófilos del Cerro de Mamapacha, Chinivita, Boyacá, Colombia. Pág. 71. En: Rangel-Ch., J.O., J. Aguirre-C & M.G. Andrade-C (eds.) 2002 Libro de Resúmenes Octavo Congreso Latinoamericano de Botánica y Segundo Congreso Colombiano de Botánica. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá. [ Links ]
15. Linares, E. L. 1986. Estudio taxonómico y ecológico de la brioflora en la franja altoandina de El Tablazo, Cundinamarca. Trabajo de Grado, Universidad Nacional de Colombia, Facultad de ciencias, Departamento de Biología. [ Links ]
16. Mägdefrau, K. 1952. Life – forms of bryophytes. Pág. 45-58. En: A.J.E Smith, (ed.) Bryophyte ecology. Londres.
17. Ochi, H. 1980. A revised of the Neotropica Bryoideae, Musci (First Part). The Journal of the Faculty of Education Tottori University. Natural Science. 29: 49-154. [ Links ]
18. Ochi, H. 1981. A revised of the Neotropica Bryoideae, Musci (Second Part). The Journal of the Faculty of Education Tottori University. Natural Science. 30: 21-55. [ Links ]
19. Orrego, O. 2000. Diversidad de los briófitos en los bosques relictuales de la zona cafetera del Departamento del Quindío. Trabajo de Grado, Universidad Nacional de Colombia. Facultad de Ciencias, Departamento de Biología, Bogotá, D.C. [ Links ]
20. Pócs, T. 1982. Tropical Forest Bryophytes. Pág. 59-104. En: A.J.E Smith, (ed.) Bryophyte ecology. Londres. [ Links ]
21. Rangel-Ch., J.O. 1995. Colombia Diversidad Biótica I. Convenio INDERENA-Universidad Nacional de Colombia, Instituto de Ciencias Naturales. Santafé de Bogotá. [ Links ]
22. Reese, W.D. 1961. The genus Calymperes in the Americas. The Bryologist 64:89-140. [ Links ]
23. Reese, W.D. 1977. The genus Syrrhopodon in the Americas I. The elimbate species. The Bryologist 80:1-31. [ Links ]
24. Reese, W.D. 1978. The genus Syrrhopodon in the Americas I. The limbate species. The Bryologist 81:189-225. [ Links ]
25. Richards, P.W. 1984. The ecology of tropical forest bryophytes. Capítulo 2 Pág. 1233-1270. En: Schuster, R.M. (ed.) New manual of bryology. [ Links ]
26. Ruíz, C.A. 2001. Estudio florístico y de vegetación del corregimiento de Tarapacá (Amazonas-Colombia). Trabajo de Grado, Universidad Nacional de Colombia, Facultad de Ciencias, Departamento de Biología, Bogotá, D.C. [ Links ]
27. Ruíz, E.J., J.C. Zabala & M.E. Morales. 2002. Aspectos taxonómico-ecológicos preliminares de Sphanum (Sphagnaceae) en Boyacá, Colombia. Pág. 66. En: Rangel- Ch., J.O., J. Aguirre-C & M.G. Andrade-C (eds.) 2002 Libro de Resúmenes Octavo Congreso Latinoamericano de Botánica y Segundo Congreso Colombiano de Botánica. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá. [ Links ]
28. Rudas, A. & J. Aguirre. 1990. Las Briofitas. Pág. 170-211. En J. Aguirre & O. Rangel (eds.) Biota y Ecosistemas de Gorgona. Fondo para la protección del medio ambiente-Instituto de Ciencias Naturales. Santafé de Bogotá. [ Links ]
29. Salazar-Allen, N. 1991. A preliminary treatment of the Central American species of Octoblepharum (Musci: Calymperaceae). Tropical Bryology 4:85-97 [ Links ]
30. Santos, G.A. 2002. Estudio florístico de los musgos (Bryophyta) en sectores de la Serranía de las Quinchas (Magdalena Medio – Colombia). Trabajo de Grado, Universidad Nacional de Colombia, Facultad de Ciencias, Departamento de Biología, Bogotá, D.C.
31. Van Reenen, G.B.A. & S.R. Gradstein. 1983. Studio on Colombia criptogams XX. A transect análisis of the bryophite vegetation along altitudinal gradient on the Sierra Nevada the Santa Marta, Colombia. Acta Bot. Neerl. 32(3):163-175. [ Links ]
32. Wolf, J.H.D. 1993. Ecology of epiphytes and epiphyte communities in montane rain forest, Colombia. University of Amsterdam, Amsterdam [ Links ]
Recibido: 07/02/2007
Aceptado: 25/05/2007

