










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.29 no.1 Bogotá Jan./June 2007
THE MYRMICINE ANT GENUS ALLOMERUS MAYR (HYMENOPTERA: FORMICIDAE)
El género de hormigas Myrmicinae Allomerus Mayr (Hymenoptera: Formicidae)
FERNANDO FERNÁNDEZ
Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Apartado 7495, Bogotá, D.C., Colombia. ffernandezca@unal.edu.co
ABSTRACT
The Neotropical myrmicine ant genus Allomerus Mayr is revised. The genus is apparently monophyletic based on the antennal club conFiguretion. I recognize 8 species (4 described as new): Allomerus brevipilosus n. sp. (Brazil), A. decemarticulatus Mayr (Brazil, French Guiana), A. dentatus n. sp. (Venezuela), A. maietae n. sp. (Brazil), A. octoarticulatus Mayr (=A. tuberculatus Forel n. syn. = A. octoarticulatus var. demerarae W. M. Wheeler n. syn. = A. novemarticulatus Wheeler & Mann n. syn. [Brazil, Bolivia, Colombia, French Guiana, Peru]), A. septemarticulatus Mayr status rev. (Brazil), A. undecemarticulatus n. sp. (Venezuela) and A. vogeli Kempf (Venezuela, Brazil). Better knowledge of the taxonomy of Allomerus is needed to understand the apparently sporadic differences in antennal flagellomere number and speciation processes that are probably linked to plant cavity colonization.
Key words. Allomerus, Ants, Formicidae, South America, Taxonomy.
RESUMEN
Se ofrece una revisión de las hormigas Myrmicinae del género Allomerus Mayr. Este es un género aparentemente monofilético debido a la conFigureción de la maza antenal. Se reconocen ocho species (cuatro nuevas): Allomerus brevipilosus n. sp. (Brasil), A. decemarticulatus Mayr (Brasil, Guayana Francesa), A. dentatus n. sp. (Venezuela), A. maietae n. sp. (Brasil), A. octoarticulatus Mayr (=A. tuberculatus Forel n. syn. = A. octoarticulatus var. demerarae W. M. Wheeler n. syn. = A. novemarticulatus Wheeler & Mann n. syn. [Brasil, Bolivia, Colombia, Guayana Francesa, Perú]), A. septemarticulatus Mayr status rev. (Brasil), A. undecemarticulatus n. sp. (Venezuela) y A. vogeli Kempf (Venezuela, Brasil). Se necesita saber más de la taxonomía y la biología de Allomerus para entender las diferencias aparentemente esporádicas en el número de segmentos de las antenas y los procesos de especiación ligados a la colonización de partes internas de las plantas.
Palabras clave . Allomerus, hormigas, Formicidae, América del Sur, Taxonomía.
INTRODUCTION
Allomerus includes a group of rarely collected, small ants, that inhabit internal cavities or plant structures in South American forests. The taxonomic understanding of the genus has been hampered by their small size, apparent homogeneity in external traits and the antennal flagelomere infraspecific variability found in the same species (or specimen: Wheeler 1942). On the other hand, this genus is known by its ability to associate with plants, which makes its promising taxa for studies in ant-plant associations or coevolution (e.g. Davidson & McKey 1993). Also, some recent studies refer to interesting aspects on the biology of the genus (Yu & Pierce 1997, Dejean et al. 2001, Corbara 2005, Dejean et al. 2005, Debout et al. 2005).
Although most classifications place Allomerus in the tribe Solenopsidini (Hölldobler & Wilson 1990, Bolton 1987, 2003), this genus (and the Old World Diplomorium) do not fit the tribal diagnosis offered by Bolton (1987, 2003). Bolton (1987) suggests Diplomorium (monotypic genus known from South Africa) as the genus closest to Allomerus, especially based on its clypeal conFiguretion. If this sister group relation is shown to be true, it might be a biogeographical distribution to be studied carefully, because no known group of sister ant taxa possesses such distribution that joins the Amazon Valley with South Africa.
This paper is not intended to be an exhaustive revision of the genus, due to problems such as the scarcity of available material and because some type material was not available (except for A. o. demerarae W.M. Wheeler, cotypes in LACM). The major justification for this study is to clarify the difficulty of species delimitation. It is fundamental to have more worker, queen and male material from the same nests, and with data regarding the plant in which the ants were collected, for further ant-plant association studies.
There is still an unsolved question regarding whether A. octoarticulatus is a variable species or a species complex (see further discussion below). The lack of observation of varieties related to that name (Wheeler 1942) makes it difficult to establish species limits within A. octoarticulatus, in case it is a complex containing sibling species.
MATERIALS AND METHODS
Measurements were made using a Nikon SMZ 2T stereomicroscope at 80X magnification and a fiber optic ring lamp. All measurements are in mm: HW - Head width: Maximum width in full face view (in males including eyes); HL - Head length: Maximum length, in full face view, from clypeus to middle of vertex; SL - Scape length (excluding basal condyle), in straight line distance; WL - Weber’s length: Length of mesosoma, diagonally from posteroventral corner of mesosoma to farthest point on anterior face of pronotum, excluding the neck; GL - Gaster length: From anterior edge of first tergum to posteriormost point; TL - Total length; CI - Cephalic index: HW/HL; SI - Scape index: SL/HW x 100. Total length includes a summation of the head, mandibular, mesosoma, petiole, postpetiole and gaster lengths, although some measurements (mandible, petiole, postpetiole) are not included in the species synopsis. Sex and castes are abbreviated as follow: l = larva, q =queen, m = male and w = worker.
Several Allomerus specimens with new name labels were found in the MZSP, but it was impossible to establish the origin of these names, as no one else is actually working on the revision of the genus (Brandão, personal communication); some of these names are used in the present paper.
Collections
CEPLAC. Centro de Pesquisas do Cacau, Comisão do Plano de Lavoura, Itabuna, Bahia, Brazil.
LACM. Los Angeles County Museum of Natural History, Los Angeles, USA.
MIZA. Instituto de Zoología Agrícola, Facultad de Agronomía, Universidad Central de Venezuela, Maracay, Venezuela.
MZSP. Museu de Zoologia, Universidade de São Paulo, Brazil.
UNCB. Insect Collection, Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá D.C., Colombia.
TAXONOMY
Genus Allomerus Mayr
Allomerus Mayr, 1878:873. Type species: Allomerus decemarticulatus Mayr, by subsequent designation of Wheeler, W.M. 1911:158.
Diagnosis. Ettershank (1966), Kempf (1975) and Bolton (1987) offered generic diagnosis, descriptions and additional notes. The following is the slightly modified diagnosis from these authors.
Monomorphic myrmicine ants within the tribe Solenopsidini and Solenopsis genus group. Worker with 6 to 11 segmented antennae, with a distinctive 3 segmented club in which each segment is constricted basally (Fig. 4B). Eyes well developed, slightly prominent, in full face view situated posterior to cephalic midline. Clypeus broad, shield-like, evenly rounded, anterior margin forming a distinct arc (Fig. 4B). Palpal formula 3,2. Propodeum unarmed to dentate.
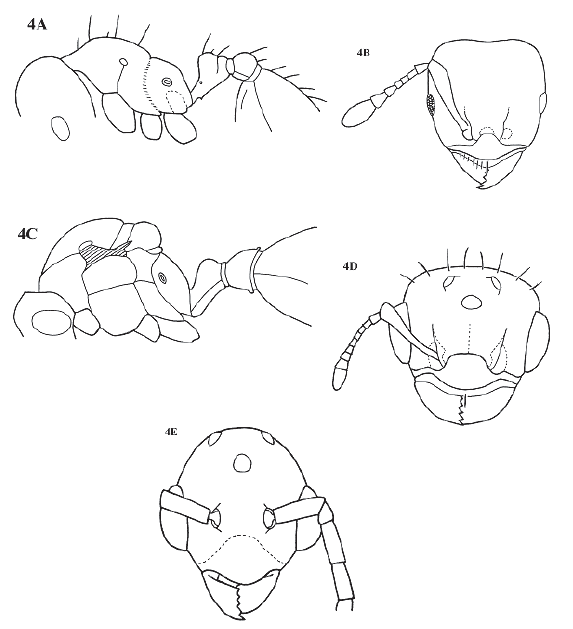
Figure 4. Allomerus octoarticulatus. A, worker in lateral view; B, worker head in full face view; C, queen in lateral view, wings omitted; D, Queen head in full face view; E, male head in full face view. Fig. 4E redrawn from Kempf 1975:349, figure 5.
Genus description, worker: Head slightly to clearly longer than wide, narrowing anteriad. Posterior border nearly flat to concave in middle, sides slightly convex. Occipital corners rounded. Mandibles with 5 teeth, generally with larger apical and subapical teeth. Clypeal anterior margin evenly rounded, forming distinct arc between mandibles, including anteclypeus or apron in most species. Frontal carinae short and widely spaced. Antennae 6 to 11 segmented, commonly 7 to 10 segmented; 3-segmented club, with each segment constricted basally. Scapes fail to reach posterior lateral corner of head, thickening apicad except in one species (A. undecemarticulatus sp. nov.). Eyes well developed, slightly convex in full face view, with numerous facets.
Promesonotum slightly to clearly convex. Promesonotal suture absent dorsally (as in Fig. 4A), impressed laterally (very feebly impressed dorsally in one species, A. undecemarticulatus n. sp., Fig. 6). Metanotal suture shallow and broad, propodeum unarmed to bidentate, dorsal face short to almost absent followed by declivity, sometimes angulate in middle. Propodeal spiracle distinct, circular, opening laterally. Bulla of metapleural gland conspicuous. Propodeal lobes small. Petiole with peduncle shorter or as long as petiolar node, node with rounded apex; petiolar spiracle near anterior side of petiole. Subpetiolar process produced as small spine or tooth followed by longitudinal carina. Postpetiole dorsally concave, lower than petiole. Postpetiole broader than long and broader than petiole. Basal portion of first tergum with shallow cuneate trench.
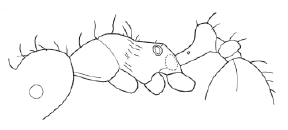
Figure 6. Allomerus undecemarticulatus n. sp. Worker in lateral view.
Body smooth and shining. Sides of mesopleura and propodeum with few longitudinal short and irregular carinulae.
Hairs scarce to abundant on body dorsum. Long, erect / suberect hairs on dorsum of head, near vertexal margin and decumbent, rows of several hairs on frons; several on promesonotum, none on propodeum, several on petiole, postpetiole and gaster. Short hairs on dorsum of mesosoma, few on propodeum. Anterior clypeal margin with setae generally short and inconspicuos, central seta sometimes feebly longer than others. Antennae and legs with appressed pubescence.
Concolorous brownish yellow to dark brown, hairs whitish.
Queen. As worker, with typical myrmicine queen morphology (Fig. 4C). Mandibles 9 to 11 segmented, with poorly defined 4 segmented club. Propodeum unarmed to slightly angulated. Wings as in Fig. 7D. Body hairy.
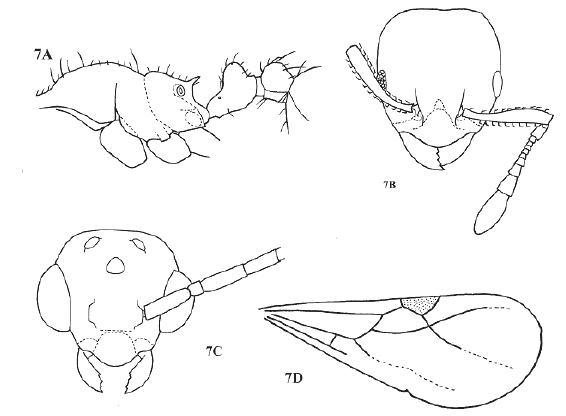
Figure 7. Allomerus vogeli. A, worker in lateral view; B, worker head in full face view; C, male head in full face view; D, male front wing. Redrawn from Kempf 1975:349, figures 1-4.
Male (Kempf 1975:350-351). Fig. 4E and Fig. 7C. Mandibles well developed, with 5 teeth on masticatory border. Clypeus shield-like, disc moderately convex, lacking carinae or grooves. Frontal carinae absent. Eyes protruding. Posterior ocelli on vertex slightly protruding. Notauli and parapsidial sutures present. Scutellum not overhanging metanotum. Basal face of propodeum longer than declivous face, unarmed. Front wing as in Figure 7D. Petiole without node, gradually thickened caudad.
Comments. Although there are no phylogenetic studies for tribe Solenopsidini, Allomerus seems to be a monophyletic genus. The clypeal conFiguretion and the constriction in the proximal base of each antennal club segment seem to be unique characteristics within the Solenopsidini (Bolton 1987). The clypeus, which is broadly inserted between the frontal antennal lobes, contradicts the tribal characterization (except for Diplomorium and Allomerus, the remaining tribe members possess a narrow insertion, making the carinae close together). As Bolton (1987) points out, the clypeal conFiguretion in Allomerus can be a plesiomorphic condition or a secondary broadening. For now, neither of the explanations can be stated as more valid than the other.
Besides the problem with its position within the tribe, another challenge relates with solving the taxonomy of species, subspecies and varieties that have been described so far in Allomerus. Species number has changed from one (Wheeler 1942) to three (Bolton 1995) with various species and varieties that are not well defined. Apart from the small size and external monotony, two other factors have hindered the taxonomical resolution of the genus. One of them is the relatively limited amount of material deposited in museums, as a result of the difficulty in capturing these organisms, which live in plant cavities and are not normally collected by standard traps or methods. Another possible factor is that the flagelomere number (reflected in the genus name) has a high intraspecific variability. Quoting Forel, Wheeler (1942) mentions that there can be a different number of antennal segments between both antennae of a single specimen! Also Kempf (1975) states that the second funicular segment can be sometimes divided or fused. Although this variation is not common, it is still an important issue in the taxonomy of the genus. Because the antennal segment number is generally unreliable, it is recommended to associate antennal segment number with other characters, especially propodeal morphology. The variation of flagelomere number (due to segment fusion or division) can be broader than observed, so large amounts of material and genetic studies will be needed to understand the nature and limits of this variation. Until then, it is understood that some conclusions made here are subject to change.
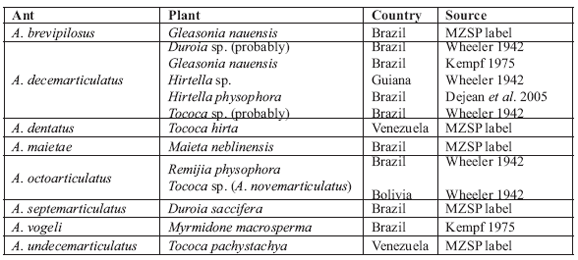
Allomerus plant associations are of high biological interest. Table 1 shows the species list and associated plants based on literature and specimen labels. Surely, more field collections will be needed before establishing the exclusiveness of some ants to certain plants. Additional information and taxonomically well defined species will result in further symbiosis and coevolutionary studies, as those dealing with the Crematogaster and Macaranga association in Asia (Itino et al. 2001).
Table 1. List of the Allomerus species here recognized and their plant records.
Dejean et al. (2001) studied predatory behavior in Allomerus decemarticulatus. More recently Dejean et al. (2005) registered trap building behavior for the first time in this species. These structures seem to help ants catch prey, and was an unknown behavior in ants.
Synonymic list of Allomerus
Allomerus brevipilosus n.sp. (w) Brazil.
A. decemarticulatus Mayr, 1878:874 (w, q) Brazil, French Guiana.
A. dentatus n.sp. (w) Venezuela.
A. maietae n.sp. (w) Brazil.
A. octoarticulatus Mayr, 1878:874 (w, q, m) Brazil, Bolivia, Colombia, French Guiana, Perú.
= A. tuberculatus Forel, 1912:2 (w, m). N. syn.
= A. octoarticulatus var. demerarae Wheeler, W.M. in Wheeler, G.C., 1935:92 (w, q, m).
= A. decemarticulatus octoarticulatus var. demerarae: Wheeler, 1942:200.
= A. novemarticulatus Wheeler, W.M. & Mann in Wheeler, W.M., 1942:199 (w). N.syn.
= A. decemarticulatus octoarticulatus var. exanguis Wheeler, W.M. & Mann, in Wheeler, W.M. 1942:200.
= A. decemarticulatus octoarticulatus var. angulatus Wheeler, W.M. & Mann, in Wheeler, W.M. 1942:201.
= A. decemarticulatus octoarticulatus var. tuberculatus: Wheeler, W.M. 1942:201.
= A. decemarticulatus octoarticulatus var. melanoticus Wheeler, W.M. & Mann, in Wheeler, W.M. 1942:202.
A. septemarticulatus Mayr, 1878:874 (w, q) Brazil. Status rev.
A. undecemarticulatus n.sp. (w) Venezuela.
A. vogeli Kempf, 1975:348 (w, m). Venezuela, Brazil.
Key for the species (workers).
1. Propodeum in side view with teeth, spines or tuberculae (as in Fig. 2) ....................… 2
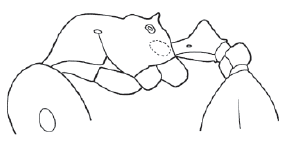
Figure 2. Allomerus dentatus n. sp. worker, lateral view.
- Propodeum in side view convex to angulate, never dentate (as in Fig. 1) ….................... 4
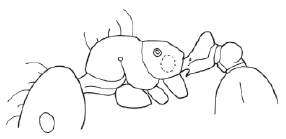
Figure 1. Allomerus decemarticulatus worker, lateral view.
2. Propodeal spiracle high, dorsal margin at same level or sligthy above propodeal dorsum (Fig. 3); hairs on head and promesonotum longer, about 0.2 mm in length; anterior clypeal setae long, especially medial seta …………........................... A. maietae n. sp.
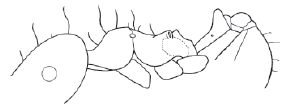
Figure 3. Allomerus maietae n. sp. worker, lateral view.
- Propodeal spiracle lower, dorsal margin distinctly below level of propodeal dorsum (Fig. 2); hairs on head and promesonotum shorter, less than 0.15 mm in length; anterior clypeal setae shorter …......................…… 3
3. Propodeal spiracle close to base of propodeal bulla (Fig. 2); antennae apparently 11 segmented …............... A. dentatus n. sp.
- Propodeal spiracle distant from propodeal bulla (Fig. 7A); antennae 10 - 11 segmented ....................................................… A. vogeli
4. Antennae 11 segmented; pronotal suture feebly impressed dorsally; propodeum sharply angulate in lateral view (Fig. 6) …..............….. A. undecemarticulatus n. sp.
- Antennae 10 segmented or fewer; pronotal suture absent dorsally; propodeum rounded or feebly angulate (Fig. 4A, Fig. 5A) …......… 5
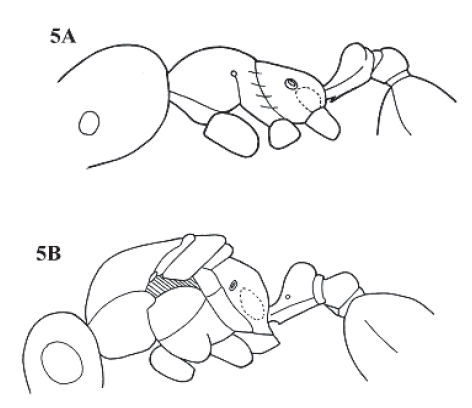
Figure 5. Allomerus septemarticulatus. A, worker in lateral view; B, queen in lateral view, wings omitted.
5. Antennae 10 segmented ….................… 6
- Antennae 9 segmented or less …............. 7
6. Clypeus with several short, longitudinal rugulae; body pilosity short, longer hairs about 0.07 mm; anterior clypeal setae very short, central seta inconspicuous …....................................................... A. brevipilosus n. sp.
- Clypeus smooth and shining; body pilosity longer, longest hairs about 0.12 mm; anterior clypeal setae longer, central seta conspicuous ….................................. A. decemarticulatus
7. Antennae 7 (rarely 6) segmented ………....................… A. septemarticulatus
- Antennae 8 (rarely 9) segmented ……… A. octoarticulatus (or A. octoarticulatus species complex, see discussion under this name).
Allomerus brevipilosus n. sp.
Worker measurements, holotype: HW 0.52 HL 0.55 SL 0.36 WL 0.60 GL 0.54 TL 2.15 CI 94 SI 69.
Worker diagnosis. Head nearly as wide as long. Antennae 10 segmented. In side view, promesonotum convex. Propodeal dorsal face very short, declivity slightly convex. Propodeal spiracle small. Petiole with peduncle shorter than node. Petilolar node with small, but well defined anteroventral spine. Body smooth and shining, especially head. Clypeal disc with several feebly irregular longitudinal carinulae. Lateral sides of propodeum finely rugose reticulate. Head with abundant short, decumbent hairs (less than 0.08 mm in length), moderate in number on promesonotum, few on propodeum, petiole and postpetiole. Few longer hairs on dorsum of head and promesonotum, none on propodeum, several on petiole, postpetiole and gaster. Row of very short, anterior clypeal hairs, medial inconspicuous. Body brown, most of head and gaster dark brown, hairs whitish.
Queen and male. Unknown.
Material examined. Holotype worker: BRAZIL, Amazonas, Naue River (MZSP), collected in Gleasonia nauensis. Paratypes: 3 workers (not available for measurements), same data (MZSP).
Comments. This species can be easily distinguished from other members of the genus with the following three characters: the short pilosity on the head, promesonotum and petiole and gaster, hairs of half or less than half the length seen in other species; short pilosity on the anterior clypeal margin without a differentiated central hair; and weak longitudinal clypeal rugulae, especially on the anterior half. The anteroventral petiolar spine is small but well defined. The label data are imprecise “Nauí river (?), Amazonas, Ducke on - Gleasonia nauensis (?)”, but according to A. decemarticulatus distributional data in Kempf (1975), it has to be treated as Naué River in Brazil’s Amazonas state.
Allomerus decemarticulatus Mayr
(Fig. 1)
Allomerus decemarticulatus Mayr, 1878:874 (w); Kempf, 1975:347 (q).
Worker measurements (n=1): HW 0.55 HL 0.58, 0.60 SL 0.33 0.55 WL 0.58 GL 0.54 TL 2.20 CI 94 SI 60.
Worker diagnosis. Antennae 10 segmented, scapes fail to reach posterior lateral corner of head, distal half thickened. Metanotal suture shallow and broad. Propodeum unarmed, dorsal face almost absent followed by declivous face, slightly angulate between faces. Propodeal spiracle small. Petiole with peduncle and with node well defined, node shorter than peduncle. Petiolar anteroventral spine well developed. Sides of mesopleuron and propodeum with few longitudinal short and irregular carinulae. Hairs abundant on dorsum of mesosoma. Long hairs (greater than 0.13 mm in length): six on front of dorsum of head, near vertexal margin and decumbent; two rows of several hairs on frons; several on promesonotum, none on propodeum, several on petiole, postpetiole and gaster. Short hairs (less than 0.07 mm) on all the dorsum of the mesosoma, few are present on propodeum. Row of short hairs present on the anterior clypeal margin, central being the longest, all projecting over anteclypeus. Antennae and legs with apressed pubescence. Concolorous brownish yellow, hairs whitish.
Queen (n=1). HW 1.05 HL 0.98 SL 0.58 WL 1.78 GL 1.50 TL 5.35 CI 108 SI 55.
Head longer than wide, eyes prominent. Antennae 11 segmented without defined club. Scutellum forms slight prominence with convex dorsal part in lateral view. Propodeum angulate between faces. Propodeal spiracle open laterally. Petiolar node shorter than peduncule; peduncule and node broadly joined. Anteroventral process of petiole acute and without tooth or spine. Smooth and polished body. Head with longitudinal striae especially on posterior part. Longitudinal rugulae on posterior part of mesoscutum. Axilar transverse band and scutellum with dense and conspicuous longitudinal striation. Propodeum with limited striation, obliquelly longitudinal below propodeal spiracle. Pilosity abundant on body (length of hairs less than antennal width), except on most of mesoscutum, anepisternum and katepisternum. Concolorous brown with black eyes and axillae.
Male. Unknown.
Material examined: 1 w, 1 q, FRENCH GUIANA, Petit – Saut, Piste de Relache, Km 7, 25 june 2000, B. Corbara Reg. 5294 (CEPLAC).
Additionally Kempf (1975) listed two Brazilian records: Amazon (Rio Naué, collected in Cleasonia (sic) nauensis, and Amapá (Serra do Navio, collected in Hirtella sp.).
Comments. Besides the 10 segmented antennae, this species can be distinguished by its mesosomal shape in lateral view (Fig. 1), in which the propodeum forms a weak angle between the faces. The petiolar shape (peduncule longer than the node) and the near lack of sculpture on the mesopleuron and lateral propodeum also makes this species easy to recognize. The anteroventral tooth of the petiole is larger than that of other workers of the genus. Almost the entire body (except for the propodeum) possesses long hairs that are longer than the last antennomere.
The holotype was not seen. The redescription above should be used cautiously because the original description is based on French Guiana queen and worker samples, in which worker possesed 10 segmented antennae. There is a worker from the MZSP (HW 0.50 HL 0.55 SL 0.28 WL 0.50 GL 0.55 TL 2.03 CI 90 SI 56) collected in Brazil (Amazonas, Manaus, Fazenda Esteio, Colosso, mata continua controle, 26 nov to 1 dic 1997, C. Klingenberg col.) with a 10 segmented antennae. This worker is smaller than the one studied from French Guiana and the propodeum is less angulate and does not show the angulation.
Although Kempf (1975:347) provides some data for the queen, he does not describe it, so the actual description has missing information. There is another queen from the same locality described above (French Guiana, Petit-Saut) collected by B. Corbara and coworkers in June 9th, 2000 (No. 5299, deposited in CEPLAC) and with a label that states “dans nid Allomerus 10-articulatus/Hirtella 4”. It is a dealate female and was collected in an apparently normal nest in terms of the population, including the queen itself. This specimen is very similar to what is here described as an A. decemarticulatus queen, although the virtual lack of erect pilosity raises attention. There are short hairs on the head, propodeum and ventral surface of the gaster, but the rest of the body can be considered as glabrous. Taking into account both queens (one described above) which were seen and referred to this species, plus the data provided by Kempf (1975:347), it can certainly be established that the A. decemarticulatus queen has moderate to dense pilosity. The glabrous queen collected in an A. decemarticulatus nest provides the bases to speculate about polygyny or symbiotic processes (parasitism) mentioned by Hölldobler and Wilson (1990). This interesting finding will be explored in further publications.
Wheeler (1942) reports an A. decemarticulatus collection made on expanded Hirtella peduncles and another (probably) on Tococa or Duroia, both from Brazil. Dejean et al. (2001) cite monogynous colonies of A. decemarticulatus in Hirtella physophora (Chrysobalanaceae), with populations of more than 1.000 workers, which patrol leaves during the day, searching for prey. The Allomerus material reported by Kempf (1975:347) as A. decemarticulatus could be A. brevipilosus, if there is species fidelity to the associated plants.
Recently Dejean et al. (2005) described trap construction to capture prey. The ants cut trichomae from the plant in which they live (Hirtella physophora), and use them to build a kind of a net on which a mycelium grows. The resulting construction is a platform-like structure with holes on it, on which the fungus grows. This platform works as a trap for jumping or flying insects that stand on it and are attacked by the ants from below. The holes have exactly the right diameter to let the ants’ heads and anterior legs pass (Corbara 2005). Dejean et al. (2005) suggest that this is a type of tripartite association (insect- plant- fungus) with the object of ambushing prey.
Allomerus dentatus n. sp.
(Fig. 2)
Worker measurements, holotype: HW 0.41 HL 0.50 SL 0.33 WL 0.45 GL 0.60 TL 2.00 CI 83 SI 79.
Worker diagnosis. Antennae apparently 10 segmented. In side view, promesonotum slightly convex. Propodeal dorsal face short, declivity and basal faces separated by pair of teeth. Propodeal spiracle aperture opens posterolaterally, with large aperture and close to base of propodeal tooth. Petiole with peduncle shorter than node. Body smooth and shining, especially head. Sides of propodeum finely reticulate-rugose. Dorsum of mesosoma with long hairs (about 0.12 mm in length), present on postpetiole and gaster; very few on petiole and none on head and mesosoma. Row of short clypeal hairs on anterior margin, medial hair conspicuous. Concolorous brown, hairs whitish.
Queen and male. Unknown.
Material Examined: Holotype: VENEZUELA, Territorio Federal de Amazonas, Río Guainía, Sabana, El Venado, Caño Pimichín, en Tococa hirta (NYBG WWB No. 568H), 23 nov 1953 (MZSP).
Comments. This new species is distinguished from the majority of the species in the genus because of its propodeal teeth and the larger propodeal spiracle that is located close to the propodeal tooth base and its opening is directed towards the sides and behind. Besides, the central apical clypeal seta is larger than its neighbors and the antennal scape thickens gradually apically. Allomerus vogeli and A. maietae are two other species with propodeal teeth or spines. Allomerus dentatus and A. vogeli are similar in their general aspect and with lateral reticulation on the mesosoma. In A. vogeli the spiracle opening is lateral (posterior in A. dentatus) and so is seen as larger in lateral view; there are also several long hairs on the thorax and head. In A. maietae the propodeal spiracle position (high in this species, Fig. 3) and hair length (nearly 0.2 mm) are characters that separate it easily from A. dentatus.
Allomerus maietae n. sp.
(Fig. 3)
Worker measurements, holotype: HW 0.54 HL 0.63 SL 0.40 WL 0.65 GL 0.59 TL 2.34 CI 86 SI 74.
Worker diagnosis. Antennae apparently 11 segmented. In side view, promesonotum weakly convex continuing into slightly concave propodeal dorsum. Propodeum with short teeth. Propodeal spiracle aperture opened lateral and upward, dorsal margin slightly above propodeal level. Petiole with peduncle larger than node. Body smooth and shining, especially head. Dorsal and lateral superior sides of propodeum with fine, short, longitudinal rugulae. Dorsum of body with very long hairs (about 0.2 mm in length); row of short anterior clypeal hairs long, medial very long, directed toward the middle of mandibles (when closed). Concolorous brown, hairs whitish.
Queen and male. Unknown.
Material Examined: Holotype: BRAZIL, Amazonas, Rio Negro, between Salesian Mission and Pirapucú Sierra, in Maieta neblinensis, 800-1000 m., 23.january.1966, N.T. Silva No. 60854 (MZSP).
Comments. Allomerus maietae is distinguished from the rest of the species by the shape of the mesosomal profile in lateral view (Fig. 3), where the weakly convex promesonotum passes smoothly into the propodeum, which is slightly concave. The propodeal spiracle is very high. The body hairs are longer than in any other Allomerus species, as well as the clypeal hairs, where the medial hair reaches half of the mandibularl length. The intermediate antennal flagelomeres are relatively slender. The type specimen was captured in Maieta neblinensis in the Amazon.
Allomerus octoarticulatus Mayr
(Figs. 4 A,B,C,D,E)
Allomerus octoarticulatus Mayr, 1878:874 (w); Forel, 1904:679 (q, m); Wheeler G.C. & Wheeler J., 1955:125 (l); Kempf 1972:19 (revised status as species); Bolton 1995:61 (catalogue).
Allomerus tuberculatus Forel, 1912:2 (w, m) ; Allomerus decemarticulatus octoarticulatus var. tuberculatus: Wheeler, W.M. 1942:201. N. syn.
Allomerus decemarticulatus octoarticulatus: Wheeler, W.M., 1942:199.
Allomerus octoarticulatus var. demerarae Wheeler, W.M. in Wheeler, G.C., 1935:92 (w, q, m); Allomerus decemarticulatus octoarticulatus var. demerarae: Wheeler, W.M. 1942:200. N. syn.
Allomerus decemarticulatus novemarticulatus Wheeler, W.M. & Mann in Wheeler, W.M., 1942:199 (w). N. syn.
Allomerus decemarticulatus octoarticulatus var. exanguis Wheeler, W.M. & Mann, in Wheeler, W.M. 1942:200 [Unavailable name].
Allomerus decemarticulatus octoarticulatus var. angulatus Wheeler, W.M. & Mann, in Wheeler, W.M. 1942:201 [Unavailable name].
Allomerus decemarticulatus octoarticulatus var. melanoticus: Wheeler, W.M. & Mann, in Wheeler, W.M. 1942:202 [Unavailable name].
Worker measurements (n=3): HW 0.49 - 0.54 HL 0.54 - 0.59 SL 0.31 - 0.36 WL 0.60 - 0.63 GL 0.58 - 0.73 TL 2.15 - 2.49 CI 91 SI 54 - 57.
Worker diagnosis. Antennae 8 segmented. Scapes fail to reach posterior lateral margin by about ¼ length, apical thickening less pronounced than in A. decemarticulatus. In side view, promesonotum convex. Dorsal face of propodeum very short, declivity convex to very slightly angulate in middle. Petiole with peduncle shorter than node; node thick, petiolar peduncle gradually curved to nodal apex. Body smooth and shining, especially head. Lateral sides of propodeum finely reticulate rugose. Large hairs (more or less 0.13 mm in length): two (sometimes three) in dorsum of head, near vertexal margin; two rows of several erect hairs on the frons; six on promesonotum, none on propodeum, several on petiole, postpetiole and gaster. Abundant short hairs (less than 0.07 mm), nearly appressed, over entire dorsum of body, few on propodeum. Row of very short hairs on anterior clypeal margin, medial longest, but nonetheless inconspicuous. Concolorous light brown, hairs whitish.
Queen measurements. HW 1.25 HL 1.12 SL 0.67 WL 1.62 GL 1.59 TL 5.39 CI 112 SI 53.
Head wider than long. Antennae 10 segmented with a poorly defined 4 segmented club. Eyes prominent, closer to the anterior head margin than the posterior border. Propodeum without teeth or spines, slightly convex to slightly angulate. Propodeal spiracle directed towards side and posteriorly. Petiole robust, node longer than peduncle, joined continuously to petiolar dorsum. Petiole anteroventral process acute, without forming spine or tooth. Lateral propodeal areas with longitudinal striation. Body covered with conspicuous pubescence. Abundant hairs (approximately as long as the last antennal segment) over head and some over propodeum, petiole and postpetiole. Abundant shorter hairs (approximately as long as penultimate antennal segment) over mesosoma and dorsum of gaster. Brown, with darker head and most of gaster; pilosity white.
Male. Not studied; head illustrated in Kempf (1975).
Material examined: 6 w, BRAZIL, Amazonas, Manaus, Fazenda Esteio, Colosso, 12 nov 1993, A.B. Casimiro leg. MZSP; 3 w, BRAZIL, Amazonas, Manaus, Reserva Ducke, in Hirtella myrmecophyla, 23 nov 1964, S. Vogel No. 221. MZSP; 2 w, BRAZIL, Mato Grosso, SINOP, 55°37’W 12°31’S, oct 1974, Alv. & Roppa legg. No. 12546 (MZSP); 1 w, COLOMBIA: Amazonas, Leticia (UNCB); 3 w, 1 q, FRENCH GUIANA: Petit – Salat No. 5299, in Cordia, 16.jun.2000, B. Corbara et al. legg. CEPLAC; 8 w, 1 q, FRENCH GUIANA: Petit – Salat, in Cordia nodosa, 26.feb.2000, A. Dejean leg. LACM; 3 w, GUIANA, Kartabo, august 1921, W.M. Wheeler leg. (Cotypes of A. decemarticulatus octoarticulatus var. demerarae) LACM; 3 w, PERU: Madre de Dios, Reserva Tambopata, TAM 566 ex Cordia, 28 feb 1987, D.W. Davidson leg. LACM; 2 w, 1 q, PERU: Madre de Dios, Reserva Tambopata, TAM 103 ex Cordia sp. 2, 26 mar 1987, D.W. Davidson leg. LACM; 6 w, 1 q, PERU: Madre de Dios, Estación Biológica Cosa Cashu, ca. 400 m, 10 nov 1985, D.W. Davidson leg. LACM; 4 w, PERU: Madre de Dios, Estación Biológica Cosa Cashu, ca. 400 m, sept - oct 1984, D.W. Davidson 5-13 leg. LACM.
Comments. Besides the 8 segmented antennae, this species is characterized by the mesosomal profile in lateral view, with the promesonotum remarkably convex, in contrast with the propodeum, which has a short dorsal face forming a continuous and slightly angulate convexity with the posterior face. The petiolar peduncle is shorter and the petiolar node is thicker. The apical and subapical teeth tend to be a little larger than the rest. The anterior clypeal hairs are very short and nearly invisible; the medial hair is hardly visible.
Both hairs on the dorsum of the head near the vertexal margin, are sometimes broken, and sometimes accompanied by a third hair. All of the short hairs are almost appressed (without being pubescent). In the majority of the observed specimens the promesonotum possesses 6 clearly defined hairs in contrast with a smooth and shiny integument. Some specimens have a shorter pilosity, as well as a slight propodeal angulation in lateral view.
This species has been collected on Hirtella myrmecophila (MZSP), leaf cavities in Remijia physophora, beneath leaves and caulinar cavities in Tocota setifera Pilger in Brazil, and caulinar cavities in Cordia hispidissima in Bolivia (Wheeler 1942). Debout et al. (2005) isolated 15 A. octoarticulatus (cf. demerarae) microsatelites, obtaining high intrapopulation variation, results that are also likely for A. decemarticulatus.
Allomerus octoarticulatus may be a species complex. It is not possible to be sure as no types of the subspecies or varieties linked to this named were seen except for those of Allomerus demerarae. I decided that the subspecies or varieties should be placed as octoarticulatus synonyms, due to the following reasons: the observation of Allomerus demerarae cotypes (LACM) and the A. tuberculatus Forel paratype picture (MCZ webpage) shows that the only difference between these ants and the A. octoarticulatus concept is based upon differences in the propodeum. In A. demerarae the propodeum is slightly longer, and in A. angulatus and A. tuberculatus it is slightly angulate. The Allomerus tuberculatus paratype photograph (MCZ webpage) shows no signs of the tubercles mentioned by Wheeler (1942) when referring to this variety. From the observed material it can be said that a gradation exists in terms of propodeal shape, from convex without angles and teeth, to highly angulate (in lateral view). On the other hand the posterior and basal propodeal faces can be simple or with a pair of lateral carinae. In the few observed females and the observations made by Wheeler (1942), it seems that the propodeum is also variable, from rounded and continuous to toothed, with the female in an intermediate condition. I prefer to interpret these observations in the sense of Allomerus octoarticulatus as a variable species. It is possible that it is really a complex with several species, but this can only be determined with more material and the observation of types.
The scarce female material shows pilosity variation as well, from sparse to moderate pilosity, with the absence of erect hairs on the propodeum, petiole and postpetiole. The orientation of the opening of the spiracle and the anteroventral petiolar process can also differ, but as the material is scarce it is difficult to make decisions.
In the case of Allomerus novemarticulatus, it has been synonymized with A. octoarticulatus for many reasons. In the first place, no 9-segmented antennae have been seen in workers. There is an A. octoarticulatus worker in which the left antennae seems to be 9-segmented, but the specimen’s level of conservation does not permit verification. On the other hand, Wheeler (1942:199) observed that A. novemarticulatus is almost identical to A. octoarticulatus, except for the 9 antennal segments. It is better to leave this species as conspecific with A. octoarticulatus until better material is available and ensures the existence of an independent Allomerus species that actually possesses 9 antennal segments.
Allomerus septemarticulatus Mayr stat. rev.
(Fig. 5 A,B)
Allomerus septemarticulatus Mayr, 1878:874 (w); Forel, 1904:680 (q); Allomerus octoarticulatus var. septemarticulatus: Forel, 1904:680; Allomerus octoarticulatus septemarticulatus: Wheeler W.M., 1942:203; Bolton 1995:61.
Worker measurements (n=4). HW 0.41 – 0.46 HL 0.46 – 0.48 SL 0.25 - 0.28 WL 0.49 – 0.53 GL 0.48 – 0.53 TL 1.83 – 1.93 CI 95 – 97 SI 57 – 59
Worker diagnosis. Anterior clypeal margin without distinct anteclypeus. Antennae 7 segmented (few workers with left antenna or both 6 segmented). Scapes fail to reach posterior lateral margin by about ¼ length, apical thickening less pronounced than in A. decemarticulatus. In side view, promesonotum convex. Metanotal suture reduced to single impressed line. Propodeal dorsal face very short, declivity and basal faces [same as dorsal face] with very feebly angulation. Propodeal spiracle small. Petiole with peduncle thickened posteriad, forming angle with node. Petiolar anteroventral spine small, well defined. Body smooth and shining, especially head. Sides of propodeum with short irregular carinulae. Long hairs (more or less 0.13 mm in length): two on the dorsum of head, near to posterior margin and down curved; two rows of several erect hairs on frons; about 8 on promesonotum, none on propodeum, several on petiole, postpetiole and gaster. Abundant short hairs (less than 0.07 mm), nearly appressed, on all body dorsum, few on propodeum. Row of very short anterior clypeal hairs, medial longest and conspicuous. All body light brown, hairs whitish.
Queen measurements (n=1). HW 0.88 HL 0.75 SL 0.50 WL 1.48 GL 1.45 TL 4.58 CI 117 SI 57
Queen. In specimen observed, both antennae with 9 segments. Smaller and paler than typical Allomerus queens. Eyes very large, filling most of head in lateral view. Most of anterior part of head, especially between eyes and frontal carinae with fine longitudinal rugulae, as well as posterior part of mesoscutum and all of scutellum. Sides of propodeum, below propodeal spiracle, with longitudinal rugulae. Eyes and ocelli black, contrasting sharply with the yellowish, pale body. Hairs golden.
Material examined. 1 w, 1 q, BRAZIL: Amazonas, Manaus, Fazenda Esteio, Colosso, in forest edge, 26.nov. – 1.dec.1997, C. Klingenberg leg. (MZSP); 1 w (antennae with six segments), BRAZIL: Amazonas, Manaus, Fazenda Porto Alegre, Reserva 3209, 6-10.nov.1997, C. Klingenberg leg. (MZSP); 1 w, 1 q, BRAZIL: Amazonas, Manaus, km 44.5 Br-174, in Duroia saccifera, secondary sandy forest, INPA, 19.vii.1981, WWB 406F (MZSP).
Comments. Types from this species were not examined. Wheeler (1942:203) stated that the A. septemarticulatus female is “smaller and paler” than that of A. octoarticulatus. The studied female matches in this aspect, that is why this material is probably conspecific with that studied by Mayr in the original description.
If the interpretation above is correct, A. septemarticulatus is not only characterized by the possession of 7 antennal segments (one specimen has 6 segments), but by the mesosomal profile, with its short propodeum and a weak angulation between posterior and basal faces. The sides of the propodeum (basal face) are defined laterally. The medial clypeal hair is evident and clearly protrudes between the others, which are very short. A specimen (Fazenda Porto Alegre, Manaus, Brazil) has 6 segmented antennae, but in its general morphology corresponds to the concept of A. septemarticulatus.
As was mentioned above, the queen is very different from typical queens that belong to the genus in being smaller, yellow colored and having large black eyes.
Allomerus undecemarticulatus n.sp.
(Fig. 6)
Worker measurements, holotype: HW 0.59 HL 0.69 SL 0.44 WL 0.76 GL 0.81 TL 3.00 CI 85 SI 74.
Worker diagnosis. Head clearly longer than wider. Antennae 11 segmented. Scapes not thickening apicad. Promesonotum feebly convex in lateral view. Pronotal suture feebly impressed dorsally. Propodeal dorsal face long, slightly below same plane of promesonotum. Propodeum sharply angulate, with dorsal face as long as posterior face. Propodeal spiracle and bullae conspicuous. Petiole with peduncle longer than node; node slender. Body smooth and shining, especially head. Sides of propodeum with longitudinal, irregular carinulae. Dorsum of body with long and short hairs (less than 0.08 mm in length): row of short anterior clypeal hairs, central conspicuous. Head light brown, rest of body brown yellowish, hairs whitish.
Queen and male. Unknown.
Material examinado: Holotype: VENEZUELA, Territorio Federal de Amazonas, Yatua River, Cerro La Neblina, in Tococa pachystachya, 1800 – 1900 m., 17.nov.1957, Maguirre et al. 42126 (MZSP).
Comments. This is an atypical Allomerus species, and may not belong to the genus. There are 11-segments, the majority of the species possesses 10 or fewer. The holotype antennae are lost, but the box in which it was separated had a label that said “undecemarticulatus”. The head is clearly longer than wide and the scape has the same width along the shaft. The propodeum is at almost the same level as the promesonotum. The presence of a weakly impressed promesonotal suture is discordant, because it is not well known from myrmicines and lacking in almost all solenopsidines. More specimens from this interesting species are needed, in order to confirm its placement in the genus.
Allomerus vogeli Kempf
(Fig. 7A, B, C, D)
Allomerus vogeli Kempf, 1975:348 (w, m); Bolton 1995:61.
In the original description, Kempf (1975) mentions that the A. vogeli worker possesses 9 flagelomeres, sometimes 10 due to the division of segments 2 or 3, the worker (Figure 1 in Kempf 1975) has 11 segments as a total. Allomerus vogeli, A. dentatus and A. maietae share in the possession of a propodeal tooth or spine, which separates them from the rest of the species that belong to the genus. Allomerus vogeli is distinguished from A. dentatus (the closest related species according to morphological traits) because of the position of the propodeal spiracle that in A. dentatus is closer to the propodeal spine than in A. vogeli.
This species is known from Mercés, Rio Negro, Amazonas, Brazil; the type material was collected in Myrmidone macrosperma (Kempf 1975).
New record. 2 w, VENEZUELA, Territorio Federal de Amazonas, San Carlos de Rio Negro, Nov.1987, K. Jaffé leg. (MIZA).
Concluding remarks. As mentioned in the introduction, the lack of type material (except for Allomerus octoarticulatus demerarae) for study and the modest number of specimens (especially females) has limited the taxonomic scope of this study. The scarcity of ants also has impeded the appreciation of species delimitation and intraspecific variation. Consequently, the synopsis proposed here must be used with caution. Surely, new collections, especially with associated workers and queens, can throw some light to the taxonomy of the genus. Some species, such as A. maieate or A. undecemarticulatus, are clearly well defined; other as A. septemarticulatus or A. brevipilosa need more data. The antennal number is of limited value, because of their apparent intraspecific variation, even in the same worker! The propodeal profile in lateral view as well as the pilosity can be potentially useful in species delimitation. The biology of his genus is promising (e.g. Corbara 2005), especially in their relationships with flowering plants belonging to families Rubiaceae, Chrysobalanaceae and Melastomataceae.
ACKNOWLEDGEMENTS
I owe special thanks to Carlos R. F. Brandão (MZSP), Jacques Delabie (CEPLAC), John Lattke & Klaus Jaffé (MIZA) and Roy R. Snelling (LACM) for the loan of critical material. To Bill MacKay (University of Texas at El Paso) for english improvement and important suggestions. To Claudia Martínez for her help in measurements and manuscript checking. Juanita Rodríguez helped with english in the first draft and Esteban Rodríguez rendered the final figures. Partial support from NSF grants DEB No. 9972024, 0205982 and EF-0337220 to Dr. Michael Sharkey (University of Kentucky) and Dr. Brian Brown (LACM) for Financial and / or logistic help from the DIB Divison of the Vicerectoría de Investigación of the Universidad Nacional de Colombia and Instituto Humboldt (Villa de Leyva). Also thanks to two anonymous reviewers for their helpful corrections and comments.
LITERATURE CITED
1. Bolton B. 1987. A review of the Solenopsis genus group and revision of Afrotropical Monomorium Mayr. Bulletin of the British Museum of Natural History (Entomol.) 54:263-452. [ Links ]
2. Bolton B. 1995. A New General Catalogue of the Ants of the World. Harvard University Press, 504 pp. [ Links ]
3. Bolton B. 2003. Synopsis and classification of Formicidae. Memoirs of the American Entomological Institute 71:1-370. [ Links ]
3. Corbara B. 2005. Les piéges des fourmis Allomerus. Insectes 138(3):15-17. [ Links ]
4. Davidson D.W. & D.B. McKey. 1993. The evolutionary ecology of symbiotic ant-plant relationships. Journal of Hymenoptera Research 2:13-83. [ Links ]
5. Debout G., R. Pereyra, B.C. Emerson y D.W. Yu. 2005. Characterization of polymorphic microsatellites in the castration parasite plant-ant Allomerus octoarticulatus cf. demerarae (Formicidae: Myrmicinae). Molecular Ecology Notes 6:182-184. [ Links ]
6. Dejean A., P.J. Solano, M. Belin-Depoux, P. Cerdan & B. Corbara. 2001. Predatory behavior of patrolling Allomerus decemarticulatus workers (Formicidae: Myrmicinae) on their host plant. Sociobiology 37:571-577. [ Links ]
7. Dejean A., P.J. Solano, J. Ayroles, B. Corbara & J. Orivel. 2005. Arboreal ants build traps to capture prey. Nature 434:973. [ Links ]
8. Ettershank G. 1966. A generic revision of the world Myrmicinae related to Solenopsis and Pheidologeton. Australian Journal of Zoology 14:73-171. [ Links ]
9. Forel A. 1904. In und mit Planzen lebende Ameisen aus den Amazonas-Gebiet und aus Peru, gesammelt von Herrn E. Ule. Zoologische Jahrbücher. Abtheilung für Systematik, Geographie und Biologie der Thiere 20:677-707. [ Links ]
10. Forel A. 1912. Formicides Néotropiques. Part 4. 3me sous-famille Myrmicinae Lep. (suite). Mémoires de la Société Entomologique deBelgique 20:1-32. [ Links ]
11. Hölldobler B. & E.O. Wilson. 1990. The Ants. Harvard University Press. 732 p. [ Links ]
12. Itino, T., S.J. Davies, H. Tada, Y. Hieda, M. Inoguchi, T. Itioka, S. Yamane & T. Inoue. 2001. Cospeciation between ants and plants. Ecological Research 16: 787-793. [ Links ]
13. Kempf W. W. 1972. Catálogo abreviado das formigas da Região Neotropical. Studia Entomologica 15:3-344. [ Links ]
14. Kempf W.W. 1975. Miscellaneous studies on neotropical ants. VI. (Hym. Formicidae). Studia Entomologica 18:341-380. [ Links ]
15. Mayr G. 1878 Formiciden. Gesammelt in Brasilien von Professor Trail. Verhandlungen der k.k. Zoologish-Botanischen Gesellschaft in Wien 27:867-878. [ Links ]
16. Wheeler G.C. 1935. The larva of Allomerus (Hym.: Formicidae). Psyche 42(2):92-98. [ Links ]
17. Wheeler G.C. & Wheeler J. 1955. The ant larvae of the myrmicine ant tribe Solenopsidini. American Midland Naturalist 54:119-141. [ Links ]
18. Wheeler W.M. 1911. A list of the type species of the genera and subgenera of Formicidae. Annals of the New York Academy of Science 21:157-175. [ Links ]
19. Wheeler W.M. 1942. Studies on Neotropical ant-plants and their ants. Bulletin of the Museum of Comparative Zoology 90(1):1-262. [ Links ]
20. Yu D. & N. Pierce. 1997. A castration parasite of ant-plant mutualism. Proceedings of the Royal Society London B 265:375-382. [ Links ]
Recibido: 12/04/2007
Aceptado: 22/05/2007

