










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.30 no.1 Bogotá Jan./June 2008
ALCIDES C. SAMPEDRO MARÍN
CATY M. MARTÍNEZ BRAVO
ANA MILENA MERCADO RICARDO
SULEIMA C. OSORIO OZUNA
YOHANA L. OTERO FUENTES
LUZ M. SANTOS ESPINOSA
RAMÓN DÍAZ TONO
Universidad de Sucre, Sincelejo, Sucre, Colombia. asampedro2002@yahoo.es
ICA, Sucre, Colombia.
RESUMEN
Esta investigación se realizó en la zona rural de los municipios Toluviejo, San Onofre y San Antonio de Palmito, pertenecientes al departamento de Sucre, Colombia, durante el período comprendido entre noviembre de 2004 y noviembre de 2005 y tuvo como objetivo la determinación de los tipos de refugio utilizados por Desmodus rotundus en las localidades mencionadas, así como conocer su composición social en esos sitios y la época reproductiva. Se hicieron capturas mediante redes de niebla, en huecos de troncos de árboles, cuevas y construcciones humanas, que mostraban evidencia de heces sanguinolentas. Los animales eran obligados a salir mediante el humo y una vez capturados eran conservados en alcohol al 70%. El número de animales en esos sitios fluctúa entre 4 y 93. La proporción de machos activos sexualmente resultó siempre menor que la de hembras con diferentes estadios reproductivos (1:6, 1:7, 1:3, 1:2, 1:2) para los diferentes refugios. Además aparecen varios machos inactivos sexualmente y neonatos. Esta composición y número parece influir en la eficiencia reproductiva, la estabilidad del grupo y en el establecimiento de su conducta de cooperación para la alimentación. Esta especie es monótoca y la reproducción puede efectuarse en cualquier época del año, lo cual garantiza la supervivencia de la misma, dadas las conocidas dificultades que afrontan cuando no pueden alimentarse.
Palabras clave.Chiroptera, Desmodus rotundus, Phyllostomidae, hematófago.
ABSTRACT
This investigation was carried out in the rural areas of Toluviejo, San Onofre and San Antonio de Palmito, in the Department of Sucre, Colombia, during November 2004 and November 2005. The purpose of the survey was to determine the type of refuge used by Desmodus rotundus in these towns, to know more about the social composition of the groups, and their reproduction times. Using various resources such as fog nets, the captures were made in the holes of trees trunks, caves, and human constructions that showed evidence of their characteristic bloody feces. The animals were forced to leave with the smoke and once captured they were preserved in alcohol 70%. The number of animals in those places is small (4 at 93), and the proportion of sexually active males was always smaller than that of females with different reproductive stadiums (1:6, 1:7, 1:3, 1:2, 1:2) for the different refuges. Several inactive males and neonates were also found. The analyzed data shows that the social composition and number affects their reproductive efficiency, the stability of the group, and its mutual feeding cooperation conduct. In spite of its feeding difficulties, this monotocous specie guarantees its survival by been able to reproduce all through the year.
Key words. Chiroptera, Desmodus rotundus, Phyllostomidae, hematofagous.
INTRODUCCIÓN
Desmodus rotundus se caracteriza por la variedad de refugios que utiliza, ya que se le puede encontrar en huecos de árboles y ramas, túneles y minas abandonadas, casas habitadas o no, así como en grietas de rocas y cuevas (Greenhall et al. 1983, Flores-Crespo & Arellano-Sota 1988, Visbal 1997, Instituto Nacional de Salud 2004, Novaes & Uieda 2004, Rodríguez 2004, Bernard 2005, Fernández 2005). Las viviendas humanas son las menos habitadas. Algunos autores han planteado que los refugios se ubican generalmente a lo largo de las márgenes de los ríos (Taddei et al. 1991, Brito et al. 2003).
La mayor parte de los autores coinciden en que las colonias de este hematófago son relativamente pequeñas y señalan números variables de animales que van desde 10 hasta 200 individuos, aunque han aparecido colonias de varios miles, las cuales son consideradas como raras por la mayoría de los investigadores (Crespo et al. 1961, Greenhall et al. 1983, Flores-Crespo & Arellano-Sota 1988, Taddei et al. 1991, Visbal 1997, Bredt et al. 1999, Rodríguez 2004, Bernard, 2005).
En cuanto a la composición social de D. rotundus en sus refugios, Wilkinson (1990) trabajó con refugios pequeños y señaló que en ellos aparece un grupo principal constituido por hembras e hijos que son protegidos por un macho dominante (alfa). Los individuos aislados y los pequeños grupos serían machos solteros, inferiores en jerarquía al macho alfa. Novaes & Uieda (2004), en refugios de mayor tamaño hallaron que una colonia está constituida por uno a tres grandes agrupamientos principales y diversos individuos aislados y/o en pequeños grupos en el interior del refugio.
La mayoría de los autores han encontrado que la reproducción de D. rotundus es poliéstrica asincrónica, sin picos de nacimientos, lactancia o preñez (Crespo et al. 1961, Langguth & Achaval 1972, Willig 1985, Alencar et al. 1994, Arellano-Sota 2004, Bernard 2005, Flores 2005); sin embargo, algunos investigadores han hallado un pico reproductivo en la estación lluviosa (Schmidt 1988, Núñez & Viana 1997). Existe una gran coincidencia en que sólo nace una cría por parto (Arellano-Sota 2004, Instituto Nacional de Salud 2004, Rodríguez 2004, Bernard 2005, Flores 2005).
No se encontraron datos en la literatura acerca de las características de los refugios, composición social ni período reproductivo para D. rotundus en Colombia. Por tal motivo, en el presente trabajo se pretende caracterizar los refugios de D. rotundus, conocer su composición social en los mismos y determinar el período reproductivo de la especie en algunas localidades del departamento de Sucre, que han sido afectadas por la actividad hematófaga sobre el ganado bovino.
MATERIALES Y MÉTODOS
El área de trabajo abarcó diferentes sitios de la zona rural de los municipios de San Onofre, Toluviejo y San Antonio de Palmito, todos pertenecientes al departamento de Sucre, Colombia. Se realizaron muestreos en los períodos de seca y de lluvia, entre noviembre de 2004 y noviembre de 2005, en horas de la mañana.
Los muestreos se realizaron en 16 refugios (9 en árboles, todos en Toluviejo y San Onofre, 6 en cuevas, una en San Antonio de Palmito y otra en Ojo de Agua, Toluviejo, y 1 debajo del puente "La Mena", en la carretera de occidente entre Sincelejo y Toluviejo. Los troncos de árbol son considerados refugios individuales, lo mismo que la cueva Ojo de Agua, el refugio "debajo del puente" y la cueva San Antonio de Palmito, solo que en esta última se efectuaron cinco visitas de muestreo. En todos los refugios la presencia de D. rotundus se verificaba por las características sanguinolentas de sus deposiciones. También se anotaban el alto y ancho de la entrada a los refugios.
La captura de ejemplares se efectuó utilizando redes de niebla de 12x2, que eran colocadas en las cercanías de los refugios y los animales eran inducidos a salir mediante humo. Cada individuo de la especie en estudio era adormecido con éter e inmediatamente se sexaba y pesaba mediante una balanza de campo Pesola con un error de ± 0.1g y se identificaba con una tarjeta numerada, antes de ser colocados en alcohol al 70%. El resto de los animales de otras especies, se identificaba con ayuda de la clave de Muñoz (2001) y posteriormente se liberaba.
Para determinar la condición reproductiva de las hembras de D. rotundus en el laboratorio, se consideraron varias categorías (Silva, 1979) atendiendo al estado de las glándulas mamarias y a la presencia o no de embriones, de la siguiente manera: hembras gestantes(G) (con embrión), hembras lactantes (L) (mamas inflamadas, con leche cuando se presionan y sin pelo alrededor del pezón), hembras ablactantes (AB) (terminaron la lactancia y presentan las mamas algo inflamadas, sin leche cuando se presionan y pocos pelos alrededor del pezón) y hembras en receso reproductivo aparente (NR). Los machos fueron clasificados como reproductores (R) si presentaban los testis escrotales y como inactivos sexualmente (NR) si estaban en posición inguinal. Además se midió el diámetro del testi derecho y se comparó entre los grupos antes mencionados.
Se verificó la normalidad de los datos referidos al diámetro de los testis mediante la prueba de bondad de ajuste de Kolmogorov " Smirnov (Zar, 1996), obtenidos en el programa StatSoft, Inc STATISTICA para Windows (1995). Los valores promedio pares fueron comparados mediante la prueba t de Student.
RESULTADOS
Las especies más abundantes resultaron, en orden, Desmodus rotundus, Notilio albiventris, Phyllostomus elongatus y Loncophyla thomasi. La mayoría de ellas se encontró en troncos de árbol de diferentes especies, algunas estaban además en cuevas y D. rotundus fue detectada en árboles, cuevas y debajo de un puente. D. rotundus puede aparecer asociado en los refugios con una o varias especies (Tabla 1). La especie de árbol donde D. rotundus apareció en mayor número fue el Caracolí (Anacardium excelsum). La asociación con otras especies no parece ser obligada, ya que en los distintos refugios pueden encontrarse varias de ellas. En los árboles de mayor talla como el Caracolí (Anacardium excelsum) y el Caucho (Ficus elastica) se detectó la mayor riqueza de especies.
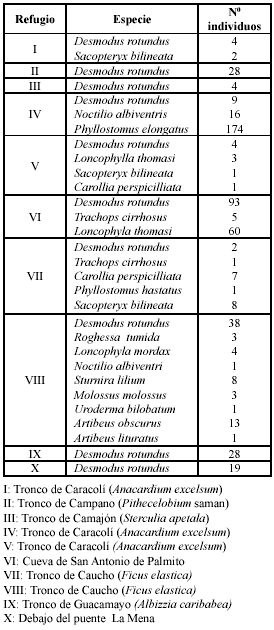
La composición social de D. rotundus en los diferentes refugios puede observarse en la Fig. 1. No se consideraron los refugios I, III, V y VII debido a que el número de individuos era pobre. En todos los casos la proporción de machos activos sexualmente resulta menor que la de las hembras (1:6, 1:7, 1:3, 1:2, 1:2) y estas se encuentran en diferente condición reproductiva (gestantes, lactantes, ablactantes, no reprodcutivas). También se observan machos inactivos sexualmente y neonatos. Debajo del puente La MENA no se hallaron machos reproductivos y en un tronco de árbol (guacamayo) situado a unos pocos metros de ese puente se hallaron machos reproductores en mayor cantidad que en otros casos (1:2). En la cueva de San Antonio de Palmito la proporción de machos reproductores en relación a las hembras reproductoras también es más baja (1:2).
De los refugios en troncos de árbol, el que presentó menor dimensión en su entrada fue el árbol de caucho (VIII), que midió 6,4cm de alto por 7,7 cm de ancho. El de mayores dimensiones fue un Caracolí (V) que midió 173 cm de alto por 60 cm de ancho, conformando lo que en esta región se acostumbra a llamar una "comba". En todos los casos los troncos de árbol se encontraban en cercanías de pequeños riachuelos. Debajo del puente La Mena se presentan algunos sitios, sobre todo en las esquinas, que constituyen refugio de los hematófagos. Una de esas estructuras tiene 267 cm de alto, un ancho de 62 cm. y una profundidad de 100 cm. Por debajo del puente fluye el agua en época de lluvias y en la de seca se forma un pozo permanente.
Se detectaron cuatro en la cueva de San Antonio de Palmito que presentaban diferentes dimensiones. La mayor es de 345 cm de ancho y una altura de 140 cm, mientras que la de menor dimensión es circular y presenta 30 cm de diámetro. A 100 m de esta cueva corre el arroyo San Antonio de Palmito.
El criterio de machos activos e inactivos sexualmente a partir de que los testis estuviesen o no en los sacos escrotales, se reforzó mediante la comparación del diámetro mayor de los testis entre cada grupo, ya que los que presentaban los testis escrotales (n = 57) resultaron tener un diámetro significativamente mayor (5.7 mm) que los que los tenían inguinales (n = 30; 3.24 mm) (t = 12.79 p < 0.001).
Se hallaron machos activos sexualmente durante el período de seca y el de lluvia y el mismo análisis en relación con las hembras demostró que se observan hembras en cualquier condición reproductiva a través de todo el año.
Fueron halladas dos hembras gestantes, que presentaban características evidentes de lactancia avanzada y con abundante leche en los pezones.
DISCUSIÓN
Desmodus rotundus resulta muy abundante en la región estudiada, debido seguramente a la gran cantidad y variedad de refugios que puede ocupar, tanto sola como asociada con otras especies y a la gran cantidad de recurso alimenticio disponible, dada la condición ganadera de esta región.
La poca cantidad de individuos de D. rotundus y otras especies halladas en algunos refugios, donde personal del Instituto Colombiano Agropecuario aplicó medidas de control con anticoagulante (Flores-Crespo et al. 1974), parece deberse a la limitación de esos animales para regresar a sus refugios, debido al estado en que los mismos se encuentran, por la acción del fuego que se ha utilizado para hacerlos salir obligatoriamente.
Ejemplares de D. rotundus fueron hallados solos en varios refugios, sobre todo en troncos de árbol de pequeña talla y esto quizás se deba a que esta especie no se relaciona directamente con otras dentro del refugio, si no que ocupa lugares dentro del mismo, que otras especies no visitan (Flores-Crespo & Arellano-Sota 1988). En efecto, se pudo comprobar que cuando estaba asociada con otras especies, éstas ocupaban sitios bien diferentes a D. rotundus, e incluso, cuando se utilizaba el humo para hacerlos salir, D. rotundus siempre era una de las últimas especies que lo hacían. Esto debe explicar la razón de que los árboles de mayor diámetro albergaran la mayor riqueza de especies. En la cueva de San Antonio de Palmito, donde también se encontraban individuos de diferentes especies, pudo observarse que D. rotundus siempre aparecía en grietas y agujeros de determinados salones, donde no había ninguna otra.
En todos los casos, los refugios se encontraban en las cercanías de algún cuerpo de agua, sobre todo pequeños arroyos o pozos con agua estancada en la época seca. Algunos autores han planteado que los refugios de D. rotundus encontrados por ellos, siempre estaban situados a lo largo de las márgenes de los ríos, en las partes bajas de las montañas (Taddei et al. 1991, Brito et al. 2003). Es posible que esto favoreciera la detección de presas a los murciélagos hematófagos ancestrales, ya que como es conocido, la mayoría de los animales pasan gran parte del tiempo en las cercanías de los cursos de agua. Esta característica debió constituir una adaptación de mucha importancia para la especie, si como todo parece indicar, se ha trasmitido de generación en generación.
Cuando se analizó la estructura social de las poblaciones de D. rotundus en los diferentes refugios, se pudo comprobar el planteamiento de Wilkinson (1990) acerca de que en los refugios estudiados por él aparecía siempre un grupo principal constituido por hembras e hijos que son protegidos por un macho dominante y los individuos aislados son machos solteros, inferiores en jerarquía al dominante. Se observó exactamente esa composición en los troncos de árbol, donde además se comprobó que los machos adultos, llamados por este autor, "solteros", se encontraban inactivos sexualmente.
Es muy posible que en la cueva estudiada los murciélagos hematófagos mantengan la misma distribución planteada anteriormente, pero en este caso no fue posible determinarlo porque el gran tamaño y distribución de la cueva no permitía capturar los animales directamente en los diferentes refugios que deben existir en su interior, sino que en las redes caían individuos perteneciente a cualquiera de esos sitios. Otros autores tuvieron la posibilidad de hacerlo en una colonia de grandes cantidades de individuos (Novaes & Uieda 2004) y determinaron que efectivamente aparecían varios grupos principales con una composición similar a la que Wilkinson (1990) encontró para pequeñas poblaciones, en este caso, troncos de árbol.
El número de individuos de D. rotundus hallados en los refugios coincide con lo reportado por varios autores (Crespo et al. 1961, Greenhall et al. 1983, Flores-Crespo & Arellano-Sota 1988, Taddei et al. 1991, Visbal 1997, Bredt et al. 1999, Rodríguez 2004, Bernard 2005) y aunque se ha dicho que esa composición y número debe estar en dependencia de la disponibilidad de alimento y espacio (Flores-Crespo & Arellano-Sota 1988, Visbal 1997), habría que considerar que los grupos pequeños brindan mayor posibilidad de actuación de la selección natural parental, también llamada familiar o consanguínea (Berovides & Alfonso 1995), que es la teoría de Hamilton, la cual favorecería la perpetuación de la conducta altruista de alimentación colectiva de las hembras de D. rotundus (Wilkinson 1990), de tanta importancia para esta especie.
Debajo del puente La Mena, no se encontró la misma proporción de machos y hembras que fue detectada en los casos discutidos anteriormente, de hecho, no se encontraron machos reproductores, quizás debido a las migraciones que caracterizan a esta y otras especies de murciélagos en diferentes épocas del año (Villa-R. 2004). En las cercanías del puente mencionado, en un árbol (guacamayo), se halló una elevada proporción de machos reproductores, lo que refuerza el planteamiento anterior.
Se ratificó el carácter monótoco de D.rotundus, como había sido señalado con anterioridad por otros autores (Arellano-Sota 2004, Instituto Nacional de Salud 2004, Bernard 2005). Tener más de un hijo cada vez, representaría un gasto energético notable, sobre todo considerando el largo período de tiempo que dedican las hembras a la gestación y cría de sus hijos (Arellano-Sota 2004, Instituto Nacional de Salud 2004, Schmidt, 2004). Algunas hembras lactantes fueron observadas en varias oportunidades cargando a su hijo durante el vuelo, que en muchas ocasiones tenían una talla y peso considerable.
Los resultados obtenidos coinciden con los de varios autores que plantean que el apareamiento parece ocurrir durante todo el año, sin "picos" de nacimientos, lactancia o preñez (Langguth & Achaval 1972, Willig 1985, Alencar et al. 1994, Visbal 1997, Arellano-Sota 2004, Bernard 2005). Se infiere que esta especie requiere para sobrevivir, una amplia y constante actividad reproductiva, lo cual puede ser considerado como otra adaptación, de las tantas que presenta la especie, para su supervivencia.
LITERATURA CITADA
1. ALENCAR, A.O., G. SILVA, M.M. DA ARRADA, A.J. SOARES & D.Q. GUERRA. 1994. Aspectos biológicos e ecológicos de Desmodus rotundus (Chiroptera) do Brasil. Pesquisa Veterinaria Brasileira 14(4): 95-103. [ Links ]
2. ARELLANO-SOTA, C. 2004. Control of bovine paralytic in Latin American and the Caribbean. National Institute for Livestock Research, México. 14 pp. [ Links ]
3. BERNARD, E. 2005. Morcegos vampiros. Sangue, raiva e preconceito. Ciencia Hoje 36 (214): 44-49. [ Links ]
4. BEROVIDES, V. & M. A. ALFONSO. 1995. Biología evolutiva. Pueblo y Educación. La Habana.1995. 407 pp. [ Links ]
5. BREDT, A., W. UIEDA & E.D. MAGAHAES. 1999. Morcegos cavernícolas da região do Distrito Federal, Centro Oeste do Brasil (Mammalia: Chiroptera). Revista Brasileira de Zoología 16(3): 731-770. [ Links ]
6. BRITO, E., H. PALACIOS, H. JUNDA, J. MARTÍNEZ & L. REYES. 2003. Sanidad animal. Rabia de origen silvestre en Colombia. Construcción de un modelo espacial para determinar áreas de riesgo. Ministerio de Agricultura y Desarrollo Rural 31(1): 7 pp. [ Links ]
7. CRESPO, J. A., J. M. VANILLA, B. D. BLOOD & J. M. DE CARLO. 1961. Observaciones ecológicas del vampiro Desmodus rotundus en el Norte de Córdoba. Revista del Museo Argentino de Ciencias Naturales Bernardino Rivadavia 4(6): 31-160. [ Links ]
8. FLORES-CRESPO, R. & C. ARELLANO-SOTA. 1988. Biology and control of the vampiro bat. In: G. M. Baer (eds.) The natural history of rabies. New York, NY, USA, Academic Press. [ Links ]
9. FLORES-CRESPO, R., R.J. BURNS & F.S. SAID.1974. Evaluación de una técnica para combatir los vampiros en sus refugios. Boletín de la Oficina Sanitaria Panamericana, 76: 427. [ Links ]
10. GREENHALL, A.M., G. JOERMANN, U. SCHMIDT, & M.R. SEIDEL. 1983. Mammalian Species, Desmodus rotundus". American Society of Mammalogists 202: 1-6. [ Links ]
11. INSTITUTO NACIONAL DE SALUD. 2004. La rabia transmitida por vampiros. Biomédica (3): 1-8. [ Links ]
12. LANGGUTH, A. & F. ACHAVAL. 1972. Notas ecológicas sobre el vampiro Desmodus rotundus en el Uruguay. Neotrópica 18:45-53. [ Links ]
13. MUÑOZ, J. 2001. Los murciélagos de Colombia. Sistemática, distribución, descripción, historia natural y ecología. Editorial Universidad de Antioquia. Medellin. 391 pp. [ Links ]
14. NOVAES, M. & W. UIEDA. 2004. Abrigo diurno, composiçao das colonias, dimorfismo sexual e reproduçao do morcego hematófago Desmodus rotundus (Chiroptera, Phyllostomidae) no Estado Sao Paulo, Brasil. Revista Brasiliana de Zoología 21 (3): 629-638. [ Links ]
15. RODRÍGUEZ, I. 2004. La rabia humana y bovina. México Ganadero. 4 pp. [ Links ]
16. SCHMIDT, C. 2004. Reproduction, p 99-109. In: A. M. Greenhall & U. Schmidt (eds.). Natural History of Vampire Bats. Boca Ratón. CRC Press. 246 pp. [ Links ]
17. SILVA, G. 1979. Los murciélagos de Cuba. Editorial Academia, La Habana, Cuba. 423 pp. [ Links ]
18. STATSOFT, INC. STATISTICA FOR WINDOWS. 1995. Computer Program Manual. Tulsa, OK: StatSoft,Inc., 2300 East 14 th Street, Tulsa, OK. [ Links ]
19. TADDEI, V.A., C.A. GONÇALVES, W.A. PEDRO, W.J. TADDEI, I. KOTAIT & C. ARIETA. 1991. Distribuçao do morcego vampiro Desmodus rotundus no Estado de Sao Paulo e a raiva dos animais domésticos. Campinas, Impresso Especial da CATI. 107 pp. [ Links ]
20. VILLA-R. B. 1966. Los murciélagos de México. Instituto de Biología, Universidad Nacional Autónoma de México. 491 pp. [ Links ]
21. VISBAL, J. 1997. Reconocimiento y control de murciélagos hematófagos. Mayores transmisores de la rabia bovina. Carta Fedegan. 8 pp. [ Links ]
22. WILKINSON, G. S. 1990. Food sharing in vampire bats. Scientific American 262: 76-82. [ Links ]
23. WILLIG, M. R. 1985. Reproductive patterns of bats from Coatingas and Cerrado Biornes in Northeast of Brazil. Journal of Mammalogy 66: 668-681. [ Links ]
24. ZAR, J.H. 1996. Bioestatistical Analysis. Tercera Edición, Prentice-Hall, Inc., Englewood Cliffs, New Jersey. 718 pp. [ Links ]
Recibido: 30/11/2006
Aceptado: 01/11/2007

