










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.30 no.1 Bogotá Jan./June 2008
HANNIER W. PULIDO-B.
M. GONZALO ANDRADE-C.
Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Apartado 7495, Bogotá, D.C., Colombia. hwpulidob@unal.edu.co;mgandradec@unal.edu.co
ABSTRACT
A new species of butterfly from the Serranía del Perijá, Cesar, Colombia, is described herein. A mountain system where the principal characteristic of the high altitude lands is the biogeographic isolation from other Andean mountain systems such as the Sierra Nevada de Santa Marta and the Sierra de Mérida in Venezuela. This new species is close to Forsterinaria punctata, but shows clear differences in the wing pattern coloration and the male genitalia morphology.
Key words. Butterflies, Satyrinae, Forsterinaria, new species, Serranía del Perijá, Colombia
RESUMEN
Se describe una especie nueva de mariposa para la Serranía del Perijá, Cesar, Colombia, un sistema montañoso cuyas partes altas presentan características de aislamiento biogeográfico con respecto al resto de los Andes y otros sistemas como la Sierra Nevada de Santa Marta y la Sierra de Mérida en Venezuela. La especie nueva es cercana a Forsterinaria punctata, pero se diferencia claramente en el patrón de manchas y en la morfología de los genitales.
Palabras clave. Mariposas, Satyrinae, Forsterinaria, especie nueva, Serranía del Perijá, Colombia.
INTRODUCTION
The Serranía del Perijá, Colombia, is the northern portion of the Andes and the Eastern Cordillera. It is located between Cesar and La Guajira, between 09º58' north 73º03' west and 10º15' north 72º57'west.The Serranía shows an altitudinal range from 150 m in the Guatapurí river Valley to 3700 m in the Cerro del Avión in Manaure Balcón del Cesar (Cesar). This area constitutes part of the border between Venezuela and Colombia. The Serranía del Perijá is an isolated narrow mountain chain from the rest of the Andes and other mountains like the Sierra Nevada de Santa Marta in Colombia, and the Sierra de Mérida in Venezuela. The paramo region and the high Andean forests of the Serranía del Perijá are unique areas in respect of the high level of butterfly endemism (Adams & Bernard 1979). The floristic composition of Hesperomeles ferruginea forests in the high elevation Andean range shows exclusive species such as Begonia corneta, Asplenium cuspidatum, Clusia cf. Multiflora, Weinmannia rollottii y Cybianthus tamanus. This flora, and the bamboo vegetation of Chusquea tessellata, is currently threatened, especially in the 2800 " 3000 m altitudinal range. The main threat factor is deforestation for farming and illicit crops. This situation has caused conditions for the establishment of vegetation mosaics of Andean region elements and paramo (Rangel & Arellano-P 2007).
These problems, in combination with construction of roads for colonization and exploitation of the natural resources of the Serranía, are the main causes of habitat fragmentation and deforestation, causing biodiversity loss.
The Serranía del Perijá is a region where craggy mountain peaks are very common. This feature of the landscape is the result of the accretion under unstable geological conditions (glacial periods, and tectonic mass movement). Its lithology is based in the upper Tertiary complex sediments formed by sandstome, except in the "Cerro del Pintao" in La Guajira, which is a Cretaceous limestone tableland (Viloria 1991).
Within the diurnal Lepidoptera, the Satyrinae is one of the most diverse groups, with a cosmopolitan distribution and nearly 2400 species occurring in every continent of the world, except in the Antarctica (Ackery et al. 1999). In the Andes, specially in forested environments of the high mountains, the Satyrinae is one of the few groups of butterflies inhabiting these altitudes (Adams 1985; Pyrcz & Wojtusiak 2002).
Within the Satyrinae, the genus Fosterinaria Gray, 1973 is placed in the subtribe Euptychiina, one of the most diverse butterfly linages in the Neotropics with more that 300 species (Murray & Pashley 2005). The Euptychiina are medium " sized butterflies with wide distributions in the Neotropics, occurring from the United States to Argentina (Peña & Lamas 2005; Murray & Pashley 2005). Although eptychiines are common in lowlands, it is possible to find some species in Andean forests. For example, species in the genus Forsterinaria can fly at 2600 m in habitats where plants of the genus Chusquea occur, the probable hostplants of the larvae (Peña & Lamas 2005).
MATERIALS AND METHODS
The specimens were collected using a butterfly net and Van Somery Rydon traps with rotten fish bait. We visited Manaure Balcón del Cesar (Cesar, Colombia) in the Vereda El Cinco and the paramo de Sabana Rubia twice (10º 21'46.4''N; 72º56'56.9''W), first in January and then in February of 2007 in a 2550 " 2700 m altitudinal range.
In the paramo of Sabana Rubia and El Cinco, that correspond to the type locality, the total annual precipitation is near 1247 mm and in April and June occurs the maximum rains, conferring a general appearance of a semihumid landscape. This physical factors promote the establishment in the type locality of habitats with chuscales and frailejonales, with open areas upper 3000m. and a dense higher Andean forest below 3000m. (Fig. 1) (Arellano et al. 2007), In the paramo, and in the transition to the forest, the typical species are Chusquea tessellate, Hypericum strictum, Calamagrostis intermedia and Espeletia perijaensis. This vegetation results in a typical physiognomy of herbaceous communities (Rangel & Arellano 2007).

Seven male specimens were collected and morphological characters were compared with a recent revision of the group (Peña & Lamas 2005). The male genitalia were extracted using hot KOH 10% during 10 minutes. Genitalia were observed using a Zeiss Stemi 2000-C stereoscope and conserved in vials with alcohol and glycerin. Photographs from the extracted aedeagus were taken in lateral and dorsal view using a digital Nikon Coolpix 8700 camera. We used AxioVision 3.1 for the measurement of genitalia and Photoshop CS2 to compose the pictures.
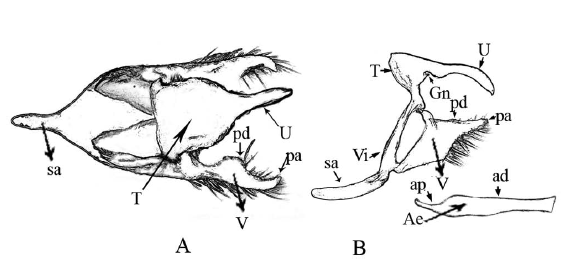
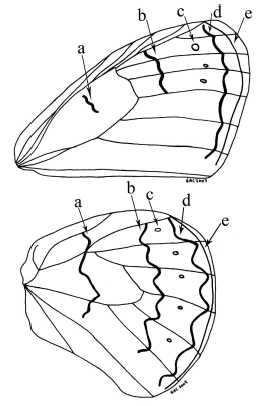
Genitalia terminology follows Klots (1970) and Peña & Lamas (2005), from which the terminology of the genitalic characters (Fig. 2) and wing lines and dots (Fig. 3) is taken. Wing vein terminology follows Miller (1970).
Color descriptions follow Ridgway (1912).
The following abbreviations are used throughout the text:
FW: Forewing
DFW: Dorsal forewing
VFW: Ventral forewing
HW: Hindwing
DHW: Dorsal hindwing
VHW: Ventral hindwing
Gen.: Genitalia
ICN-MHN-L: Instituto de Ciencias Naturales, Colección de Lepidoptera, Universidad Nacional de Colombia
RESULTS AND DISCUSSION
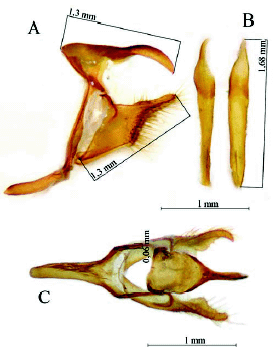
Forsterinaria anachoreta Pulido & Andrade, sp. nov. (Fig. 4, Fig. 5)

HOLOTYPE: Male, COLOMBIA, Cesar, Manaure Balcón del Cesar, Vereda El Cinco, 10º21'46,4'' north, 72º56'58,9'' west, 2650 m, 27 Feb. 2007, H. Pulido-B. Leg., HP 0954, Gen No. 1043, ICN-MHN-L 20246.
PARATYPES: Male, COLOMBIA, Cesar, Manaure Balcón del Cesar, Vereda El Cinco, 10º21'46,4'' north, 72º56'58,9'' west, 25 Feb. 2007, 2550 m H. Pulido-B. Leg., HP 1127, Gen. No. 10445, ICN-MHN-L 20231; male, HP 0951, Gen No. 1044, ICN-MHN-L 20242; male, 28 Feb. 2007, 2700 m, H. Pulido-B. Leg., HP 983, Gen. No. 1047, ICN-MHN-L 20244; male, HP 0995, ICN-MHN-L 20863; male HP 1004, Gen. No. 1046, ICN-MHN-L 20245; male, HP 1002, ICN-MHN-L 20864.
Diagnosis: FW length: 22,3 " 24,4 mm (n = 7). Separated from Forsterinaria punctata Peña & Lamas, 2005because the submarginal line in the VHW F. anachoreta is regular and ends in the cell Cu2-2A. The uncus, in the male genitalia of F. anachoreta is thinner, the dorsal process is proximal and shows three small teeth between the dorsal process and the distal end. The aedeagus can be folded on itself in the distal end in some individuals.
Description: Male, FW length: 22,6 mm. Head: labial palpi with long scales (2,4 mm), which is twice the eye length (1,29 mm); antenna sepia which total length is half of the costal vein length. Thorax: Color sepia. Abdomen: Color sepia. Genitalia (Fig. 5): Uncus (0,81 mm) twice the total length of the tegumen (0,49 mm), curved ventrally and distal end is like a hook; gnathos very reduced; valva dorsal process is rounded and located at the base, three little sharp teeth are in the inner margin of the valva (Fig. 5C), aedeagus tubular (1,68 mm) (Fig. 5B) with no torsion and no constriction. The scales in distal margin of both wings are jutted out like cilia. Forewing (FW): DFW color sepia with filiform scales in the basal area longer than those of the remainder of wing. VFW color sepia from basal area to submedial area and color raw umber in the remainder of wing; a slight and diffuse discal line fuscous black is present, postdiscal line fuscous black, better defined and irregular from vein R4 to vein M3; submarginal line fuscous black with a regular undulated pattern from vein R4 to the cell Cu2-2A; the marginal line fuscous black is present from costal margin to the cell Cu2"2A; with three submarginal white ocelli, largest in cell R5-M1, the smallest in cell M1-M2, and, a medium-sized in cell M2-M3. Hindwing (HW): DHW sepia with filiform scales in the basal area longer than those of the remainder of wing. VHW sepia from basal area to submedial area and color raw umber in the remainder of wing; discal line from Sc+R1 to the inner margin; postdiscal line with a regular undulated pattern from Rs to the cell Cu2-2A; submarginal line and marginal line with the same pattern from Rs to the cell Cu2-2A; five white ocelli in cells Rs-M1, M1"M2, M2"M3, M3"Cu1, Cu1"Cu2, respectively.
Female: Unknown.
Etymology: The specific epithet is derived from the medieval latin "Anachoreta", which means living in a solitary place. The new species is named anachoreta in reference to its isolation in the high-Andean forests of the Colombian Serranía del Perijá.
Distribution and ecology: This species is only known from the type locality, Manaure Balcón del Cesar in Cesar, Colombia. It flies between 2550 " 2700 m of altitude. The species is found in the non-deforested high " Andean forests.
Discussion: The new species is described into the genus Forsterinaria because shows several synapomorphies that define the genus: on ventral surface, both wings present undulated dark brown lines, present a discal, postdiscal submarginal and marginal lines and submarginal ocelli (Peña & Lamas 2005), and it differs from the most closely species of the genus Guaianaza in the elongated valva shape, its bigger size and the ocelli presented in the HW underside of Forsterinaria are simple white dots, while in Guaianaza these ocelli are light brown with a white dot in the center (Freitas & Peña 2006).
The discal, postdiscal, submarginal and marginal lines, and submarginal ocelli in VFW and VHW of F. anachoreta, are similar to F. punctata, but these can be distinguished from F. anachoreta because in VFW the postdiscal line is present from vein R4 to vein M3, whereas in F. punctata this line is present from the costal margin to vein M3; in F. anachoreta the submarginal line has a regular undulated pattern, while in F. punctata there is a slightly undulated pattern. In F. anachoreta, the VHW discal line goes from Sc+R1 to the cell Cu2-2A, while in F. punctata it reaches the inner margin. In F. anachoreta, the VHW postdiscal line has a regular undulated pattern from vein Rs to the cell Cu2-2A, and is present from the costal margin to near the inner margin; the submarginal line has a regular undulated pattern from vein Rs to the cell Cu2-2A, while in F. punctata the same line shows a zig-zag pattern, with segment M2 y M3 more developed and extends from costal margin to near the inner margin. In F. anachoreta genitalia, the uncus is narrower in lateral view (0.09 mm) (Fig. 5A) and the dorsal process is less proximal than in F. punctata; F. anachoreta shows three small sharp teeth between the dorsal process and the distal end in the inner margin of the valva, absent in F. punctata.
Forsterinaria anachoreta is superficially very similar to F. punctata and this might cause one to think it is best to describe the new taxon as a subspecies. However, proposing and recognizing subspecies requires evidence of gene flow among the putative races and not a subjective view that considers the observed differences are less than of "species" value. Pertinent to our proposal that F. anachoreta and F. punctata are different species are the following two facts: (1) there is not detected variation in either character used here (color pattern on the VFW and form of the uncus in our sample of F. anachoreta) and (2) the characters states used here are not shared by F. anachoreta and F. punctata.
ACKNOWLEDGMENTS
We are very grateful to Orlando Rangel, Biodiversity and Conservation Group Coordinator in the "Instituto de Ciencias Naturales" of the Universidad Nacional de Colombia, and with "Corporación Autónoma Regional del Cesar CORPOCESAR" for funding the Project "Estado actual del Páramo y Plan de Manejo Integral". To Gerardo Lamas and Carlos Peña for comments about the confirmation of this new taxon and to John D. Lynch for the discussion concerning species and subspecies. To Luis A. González and Lina R. Campos for mounting the specimens and/or discussion on the final text. To the TABDP Project by providing the photographic camera for taking the pictures of the specimens and genitalia.
LITERATURE CITED
1. ACKERY, P.R., R. DE JONG, & R.I. VANE-WRIGHT. 1999. The butterflies: Hedyloidea, Hesperoidea and Papilionoidea. pp. 263-300. In: Kristensen, N.P. (Ed.), Lepidoptera: Moths and Butterflies. Vol. 1. Evolution, Systematics and Biogeography. Handbook of Zoology, vol. IV. Walter de Gruyter, Berlin. Part 35. [ Links ]
2. ADAMS, M.J. & G.I. BERNARD. 1979. Pronophiline butterflies (Satyridae) of the Serranía de Valledupar, Colombia-Venezuela border. Syst. Ent. 4:95-118. [ Links ]
3. ADAMS, M. J. 1985. Speciation in the pronophiline butterflies (Satyridae) of the northern Andes. J. Res. Lepid. Suppl. 1:33"49. [ Links ]
4. ARELLANO-P, H. J.O. RANGEL-CH., GARCÍA-M., A.M. 2007. Clima y Topoclima. En: J.O. Rangel-Ch (Ed.) Colombia Diversidad Biótica V. Instituto de Ciencias Naturales - Corpocesar. 19-41 pp. Bogotá D.C. [ Links ]
5. FREITAS, A.V.L & C. PEÑA, 2006. Description of genus Guaianaza for "Euptychia" pronophila (Lepidoptera: Riodinidae): Are they consistently moving up in the world? Proc. R. Soc. B 272: 2457 - 2466. [ Links ]
6. KLOTS, A.B. 1970. Lepidoptera. pp. 115-130. In: Tuxen S.L (ed.). Taxonomist's Glossary of Genitalia in Insects. Ed. 2. Copenhagen, Munksgaard. [ Links ]
7. LAMAS, G (Ed). 2004. Atlas of Neotropical Lepidoptera. Checklist part 4A. Hesperioidea-Papilionoidea. Association for tropical Lepidoptera, Florida 439 p. [ Links ]
8. MILLER, L.D. 1970. Nomenclature of wing veins and cells. Journal of Research on the Lepidoptera 8(2):37-48. [ Links ]
9. MURRAY, D. & D.P. PASHLEY. 2005. Molecular phylogenetics and evolutionary history of the neotropical Satyrinae Subtribe Euptychiina (Nymphalidae: Satyrinae). Molecular phylogenetics and evolution 34 67-80. [ Links ]
10. PEÑA, C. & G. LAMAS. 2005. Revision of the butterfly genus Forsterinaria Gray, 1973 (Lepidoptera: Nymphalidae, Satyrinae). Rev. Peru. Biol. 12: 5-48 [ Links ]
11. PYRCZ, T.W. & J. WOJTUSIAK. 2002. The vertical distribution of pronophiline butterflies (Nymphalidae, Satyrinae) along an elevational transect in Monte Zerpe (Cordillera de Mérida, Venezuela) with remarks on their diversity and parapatric distribution. Global Ecol. Biogeogr. 11: 211-221. [ Links ]
12. RANGEL-CH., J.O. & H. ARELLANO-P. 2007. La Vegetación de la alta montaña de Perijá. En: J.O. Rangel-Ch (Ed.) Colombia Diversidad Biótica V. Instituto de Ciencias Naturales - Corpocesar. 163-189 pp. Bogotá D.C. [ Links ]
13. RIDGWAY, R. 1912. Color standards and nomenclature. Published by the author, Washington, D.C., pp. 52 plates. [ Links ]
14. VILORIA, A.L. 1991. Los páramos de Perijá. Natura 93:25-29. [ Links ]
Recibido: 31/10/2007
Aceptado: 03/04/2008

