










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.30 no.2 Bogotá July/Dec. 2008
GLADYS CÁRDENAS-ARÉVALO
ORLANDO VARGAS-RÍOS
Instituto de Ciencias Naturales, Grupo Biodiversidad y Conservación. Universidad Nacional de Colombia, Apartado 7495, Bogotá D. C., Colombia. gladysc4@hotmail.com
Departamento de Biología. Universidad Nacional de Colombia. Bogotá D. C., Colombia. jovargasr@unal.edu.co
RESUMEN
Se estudiaron once rasgos de historia de vida (morfológicos y de regeneración) de plantas vasculares, en cuatro sitios con diferente grado de alteración, que hacen parte de una misma unidad de paisaje y condiciones topográficas similares, ubicados en los valles de los ríos Tunjo y Piedras Gordas, Páramo de Palacio, Parque Nacional Natural Chingaza, Cundinamarca, entre 3400 y 3600 m de altitud, a los 4º45'03''N, 73º50'50''W. Estos valles, principalmente el valle del río Tunjo, tienen una larga historia de disturbio por quema y pastoreo de ganado vacuno; su comunidad natural es de Espeletia killipii, Chusquea tessellata y Calamagrostis effusa. Mediante un análisis de clasificación y un procedimiento de componentes principales (ACP) se agruparon las especies de acuerdo con rasgos similares y se analizó su abundancia de acuerdo con el grado de alteración. Se identificaron tres grupos de especies, diferenciados principalmente por los rasgos morfológicos: tipo de planta, forma de crecimiento, hábito y altura. El grupo uno se caracteriza principalmente por especies graminoides (73 %), erectas (100 %) en macolla (68 %) con alturas mayores a 5 cm (60 %); este grupo no es favorecido en su abundancia-cobertura al aumentar el grado de alteración, presenta su mayor abundancia en los sitios con alteración leve y sin alteración (33 y 30 %). El grupo dos por hierbas rastreras o postradas (76 %) que forman cojines o tapetes (86 %) con alturas menores a 5 cm (79 %), es característico del sitio con alteración alta donde presentó la mayor abundancia (44 %). El grupo tres por hierbas (88 %) en roseta (100 %), que no forman cojines o tapetes, presentó su menor abundancia en el sitio con mayor grado de alteración (8 %).
Palabras clave. Alteración, historias de vida, páramo húmedo, rasgos.
ABSTRACT
Eleven life history traits of vascular plants morphology and regeneration) were studied in four sites with different degrees of disturbance. They belong to the same landscape unit and have similar topographic conditions. The four study sitesare located in the Tunjo and Piedras Gordas valleys, Paramo of Palacio, Chingaza National Natural Park, Cundinamarca, between 3400-3600 m and at 4º45' 03'N, 73º50`50' W. These valleys (mainly the Tunjo River valley) have a long history of disturbance by burning and livestock grazing; the native community is composed of Espeletia killipii, Chusquea tessellata and Calamagrostis effusa. Through analysis of vegetation classification and principal components analyses (PCA), the species were grouped according to their similar traits; the abundance was analyzed according to the degree of disturbance. Three species groups were identified and differentiated mainly by their morphologic traits: type of plant, growth form, habit and height. Group one is characterized mainly by graminoid species (73%), erect habit (100%) in tussocks (68%) with height above 5 cm (60 %); the -abundance cover of the group is lower when the degree of disturbance increases, it is most abundant in places with light or no disturbance (33 and 30 % respectively). Group two is characterized by dragging or prostrate herbs (76%) that form cushions or carpets (86%) with height of under 5 cm (79%), occuring in places with low disturbance, where it is most abundant (44 %). Group three is characterized by herbs (88%) in rosettes (100%), that do not form cushions or carpets, was least abundant at the most disturbed site (8%).
Key words. Disturbance, life histories, humid paramo, traits.
INTRODUCCIÓN
Durante las dos últimas décadas, varios autores han hecho énfasis en la importancia de las características autoecológicas y los rasgos de historia de vida, para identificar patrones de las especies en una comunidad, encontrando importantes asociaciones entre los rasgos, las condiciones ambientales particulares y el régimen de disturbios (Grime 1979, 1989, Noble & Slatyer 1980, Halpern 1989, Pickett et al. 1989, Rydin & Borgegard 1991, Vargas 1997). Lavorel et al. (1999), usan el término rasgo (trait) para referirse a las características de las plantas, el cual puede tener varios atributos que son diferentes expresiones o estados de un rasgo (en el mismo sentido en que lo usan Campbell et al. 1999, Kleyer 1999 y McIntyre et al. 1999 b). Los atributos se pueden medir con varios niveles de resolución y se pueden expresar en categorías o rangos, una combinación específica de atributos define un tipo funcional, los cuales son grupos de especies que comparten algunos rasgos adaptativos con una función específica; la identificación y estimación de su abundancia es importante para describir los mecanismos bajo los cuales, responde la vegetación a diferentes factores ambientales (McIntyre & Tremont 1995, Díaz et al. 1999, McIntyre et al. 1999 a), para contribuir al entendimiento de la función de un ecosistema (Gitay & Noble 1997) y como una herramienta para predecir respuestas de los ecosistemas a cambios introducidos por el hombre a escala global (Díaz & Cabido 1997, Díaz et al. 1999).
En la actualidad muchos ecosistemas en el mundo son quemados y pastoreados por ganado vacuno, equino, ovino y caprino; en la mayoría existen comunidades vegetales muy sensibles a estos disturbios. Los páramos, son extremadamente vulnerables a los factores antrópicos, la vegetación apenas tolera bajas frecuencias de quema y pastoreo (Hofstede et al. 1998); este ecosistema no evolucionó bajo fuegos frecuentes ni bajo pastoreo de grandes herbívoros. Cuando se estudia el impacto de disturbios antrópicos como fuego y pastoreo sobre la vegetación de páramo, es importante tener en cuenta los eventos que ayudaron a su conformación y las condiciones que actualmente lo caracterizan (Van der Hammen & Cleef 1986), para entender el origen de las adaptaciones de sus especies, que son una consecuencia del clima extremo; Monasterio (1980), señala que estos ecosistemas se estructuraron bajo cambios drásticos, muchas veces catastróficos y que las oscilaciones polianuales condicionan e intercalan ciclos de mayor estrés.
Los factores limitantes del páramo, sumados a los disturbios naturales y antrópicos como fuego y pastoreo, seleccionan ciertas especies con rasgos de historias de vida, que les permiten permanecer bajo estas condiciones. Dentro de las formas de crecimiento, los biotipos o bioformas en roseta que predominan en el páramo, están mejor adaptadas a resistir bajas temperaturas y alta radiación que fuegos anuales (Sturm & Rangel-Ch. 1985, Hofstede 1995, Hofstede et al. 1998, Vargas 1997). En fondos de valles y lugares planos, el pastoreo se relaciona con una matriz de hierbas rastreras y pastos cortos, que tienen ventaja sobre formas altas intolerantes al disturbio por pastoreo (Verweij & Budde 1992). Verweij (1995) en su investigación en el Parque Nacional Natural Los Nevados, encontró para las especies en roseta con tallo como las espeletias, que bajo la influencia de quema y pastoreo en el páramo, se puede observar un gran número de individuos pequeños, mientras que los individuos altos mueren o tienden a desaparecer, con lo cual se corre el riesgo de una extinción local de estas especies. Pels & Verweij (1992), registran en sitios con pastoreo moderado del páramo, plantas mas bajas en altura y en diámetro que las de sitios no pastoreados.
Actualmente el páramo, experimenta un acelerado proceso de transformación por diferentes actividades humanas, principalmente la ganadería y la agricultura que casi siempre van ligadas a la utilización del fuego, lo que pone en peligro su vegetación, biodiversidad y función reguladora de la hidrología de los Andes. Con el fin de entender cómo se ve afectada la vegetación por diferente grado de alteración, se estudiaron algunos rasgos de historia de vida de especies que conforman una comunidad vegetal alterada en un páramo húmedo y se identificaron grupos de plantas con rasgos de historia de vida semejantes y se analizó su respuesta en abundancia de acuerdo con el grado de alteración.
MÉTODOS
Área de estudio
Está ubicada a los 4º45'03''N, 73º50'50''W, en el páramo de Palacio entre 3400 y 3600 m de altitud, páramo húmedo o 'bamboo páramo' según Cleef (1981), que actualmente hace parte del Parque Nacional Natural Chingaza, Cundinamarca, Colombia. El clima se caracteriza por temperaturas medias que fluctúan entre 5.7 y 10.5 ºC; pluviosidad con un promedio anual de 1680 mm en el área de la laguna de Chingaza, aumentando hasta un promedio anual de 2327 mm en el campamento de Chuza (monte redondo) y de 3322 mm en el sitio denominado El Dedal (cuenca del río Chuza). La distribución de la precipitación a lo largo del año, tiene un solo pico de lluvias (unimodal), con un máximo en junio y julio (EAAB 1997, Rangel-Ch. 2000). Los suelos son ácidos (pH: 4.3-4.9), con capacidad de intercambio catiónico alto, muy ricos en calcio, con notable cantidad de aluminio y escasez de potasio. La vegetación del área ha sido estudiada por Cleef (1981), Franco (1982), Sturm & Rangel-Ch. (1985), Rangel-Ch & Ariza-N. (2000), entre otros. De acuerdo a estos estudios predomina la vegetación de tipo chuscal, donde Chusquea tessellata forma comunidades muy homogéneas; la presencia de esta especie en el área se explica tanto por la precipitación alta como por la constante humedad atmosférica y suelos permanentemente húmedos (Rangel-Ch. & Ariza-N. 2000, Zuluaga 2002). En la zona también se encuentran pajonales de Calamagrostis effusa, grandes áreas abiertas de frailejón (Espeletia grandiflora) con suelos bien drenados, áreas de suelos anegados con Espeletia killipii y zonas mal drenadas con turberas (Ciperáceas, Juncáceas y musgos del género Sphagnum).
Los sitios de muestreo hacen parte de una misma unidad de paisaje con condiciones topográficas similares (pendiente suave < 5 %), suelos negros y con un nivel freático cerca de la superficie; ubicados tres en el valle del río Tunjo (sitios I-III), cuyo patrón geomorfológico es analizado detalladamente por Páez (2002) y uno en el valle de Piedras Gordas (sitio IV). Estos valles, principalmente el del río Tunjo tienen una larga historia de disturbio por fuego y pastoreo de ganado vacuno. Actualmente se presenta pastoreo ocasional, en la parte media y baja del valle del río Tunjo; aunque el fuego estuvo presente en el pasado con una última quema en febrero de 1991 (según registros del Parque), que abarcó la parte media y baja del valle, sus efectos directos ya no son evidentes. Vargas et al. (2002), establecieron de manera indirecta un gradiente de intensidad de pastoreo actual a lo largo del valle. Con base en este estudio y los resultados de Premauer (1999) y Posada & Cárdenas (1999), se seleccionaron cuatro sitios: uno con alto grado de alteración (Sitio I), cuya vegetación actualmente consiste en un prado con un estrato de vegetación rasante, dominada por Lachemila orbiculata, Trifolium repens, Bidens triplinervia entre otras; uno con grado de alteración medio (Sitio II), cuya vegetación está dominada por Chusquea tessellata y Calamagrostis effusa y dos sitios, uno con grado de alteración leve (Sitio III) y otro sin alteración antrópica (Sitio IV), donde dominan Espeletia killipii, Chusquea tessellata y Calamagrostis effusa.
Muestreo y toma de datos
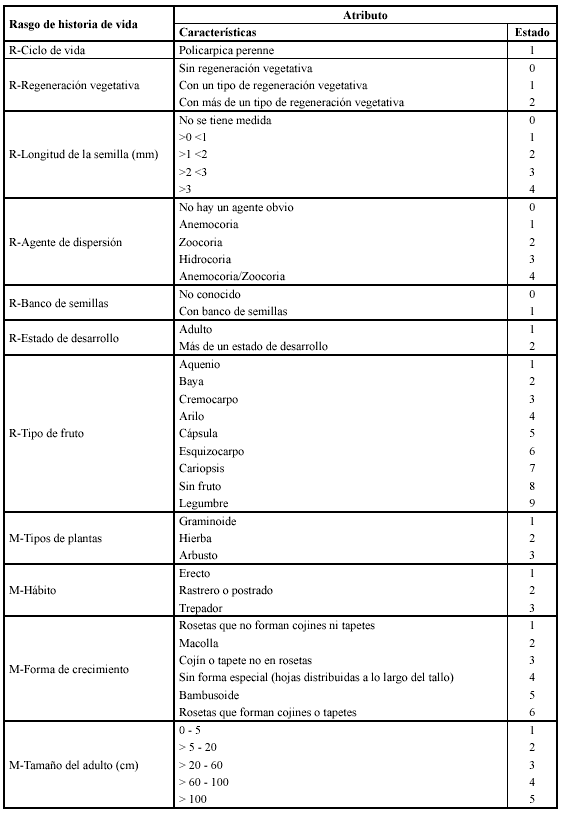
El muestreo se realizó cada dos meses de enero a diciembre de 1999; en cada uno de los cuatro sitios seleccionados, se trazaron tres transectos de 30 metros de longitud, en estos se ubicaron cuadrados de 0.5 m x 0.5 m separados tres metros entre sí (diez cuadrados por transecto). Por cuadrado se registraron las especies presentes, altura, cobertura, presencia de plántulas, juveniles y adultos por especie y se colectaron ejemplares de cada una para su identificación. Se consideraron once rasgos de historia de vida (de regeneración y morfológicos) para las especies vasculares, que se encontraron en los cuatro sitios. Los atributos de los rasgos considerados se establecieron de acuerdo a medidas y observaciones en campo y a la información disponible en literatura y se expresaron en categorías o clases para su análisis (Tabla 1). La información sobre si las especies tienen banco de semillas se tomó de la investigación desarrollada por Posada & Cárdenas (1999) y Cárdenas et al. (2002), en los mismos sitios de muestreo del presente trabajo.
Análisis de datos
Para ver qué grupos de especies con rasgos similares se tienen de acuerdo con los rasgos estudiados, se tomó como factor: los once rasgos; variable de observación: la presencia - ausencia de cada uno de los rasgos en las especies; parámetro o variable de respuesta: la distribución porcentual de las especies en cada uno de los rasgos; estrategias de análisis: la cuantificación de las variables categóricas, grupos de especies con rasgos similares, asociación entre variables y factores o rasgos que más inciden en la agrupación; prueba estadística: procedimiento de componentes principales según Young et al. (1978). El procedimiento ha sido integrado al paquete estadístico SAS/STAT (1990) con la denominación PRINQUAL; análisis de clasificación jerárquica utilizando como medida de similitud el índice de Ward (Ward's Minimum Variante Cluster Análisis); análisis de componentes principales y Test de Duncan (Sokal & Rohlf 1979).
Para ver si existen cambios en la estructura de los grupos de las especies, con rasgos de historia de vida semejantes de acuerdo con el grado de alteración, se tomó como factor 1: grado de alteración (alta, media, leve, sin alteración antrópica); factor 2: grupos de plantas con rasgos similares; variable de observación: cobertura de las especies; parámetro o variable de respuesta: distribución de cobertura en función de cada grupo por sitio; estrategias de análisis: variación de la cobertura de los grupos entre sitios, diferencias entre pares de sitios para cada grupo; prueba estadística: prueba de Chi- cuadrado y test de proporciones (Sokal & Rohlf 1979).
RESULTADOS
Grupos de especies con rasgos de historia de vida similares
En los cuatro sitios se registraron 82 especies, de las cuales doce son musgos, 42 hierbas, 22 graminoides y seis arbustos. Para los once rasgos de historia de vida estudiados, se tienen 44 estados de carácter (estados de atributos) (Tabla 1).
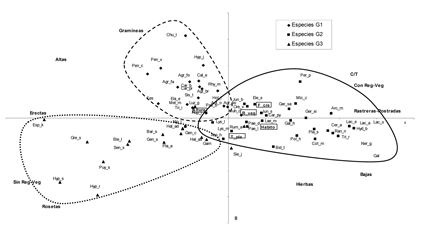
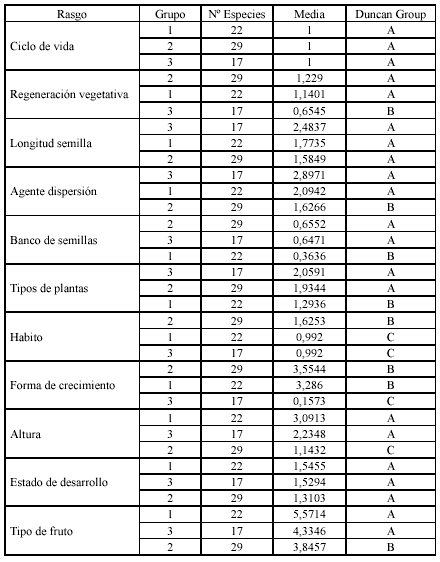
De acuerdo con el análisis de clasificación de la matriz especies vs rasgos (Anexo 1), se identificaron tres grupos principales de especies vasculares; cada grupo se caracteriza por varios rasgos que predominan sin ser exclusivos (Tabla 2). La tendencia de variación entre grupos se refleja principalmente con los rasgos morfológicos: tipo de planta, hábito, forma de crecimiento y altura; en el primer eje del plano obtenido con el análisis de componentes principales (Figura 1), se separan en el extremo derecho, las plantas con alturas generalmente entre 0 y 5 cm de hábito rastrero o postrado que forman cojines o tapetes (grupo 2) de las plantas con alturas generalmente mayores de 5 cm y hábito erecto (grupos 1 y 3). En el segundo eje se separan las graminoides (grupo 1) de las hierbas en roseta (grupo 3). Las especies en el plano aparecen distribuidas o segregadas de acuerdo a la similitud entre rasgos, por ejemplo Chusquea tessellata hace parte de las graminoides, pero es la única con forma de crecimiento bambusoide y presenta un rango de altura máximo, por esta razón se encuentra en un extremo y casi al mismo nivel (en altura) con Espeletia killipii y las especies de Pentacalia. De acuerdo con el test de Duncan (Tabla 3), se tienen diferencias significativas entre el grupo uno con los grupos dos y tres en banco de semillas y tipo de planta; entre el grupo dos con los grupos uno y tres en hábito, altura, agente de dispersión y tipo de fruto y entre el grupo tres con los grupos uno y dos en el tipo de regeneración vegetativa y forma de crecimiento. No se encontraron diferencias significativas entre los grupos con respecto a la longitud de la semilla y al estado de desarrollo. El ciclo de vida para todas las especies fue policárpico perenne.
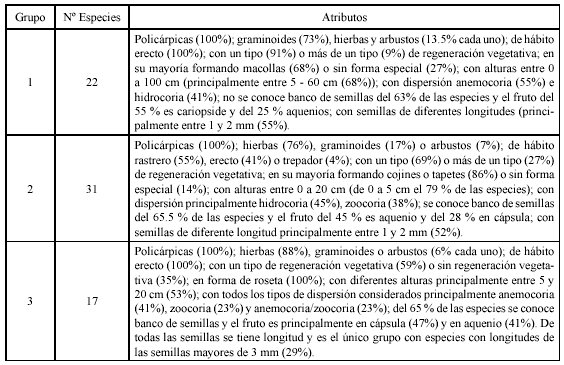
Teniendo en cuenta que aún existen interrogantes en varios aspectos, principalmente en los rasgos de regeneración, en las fig. 2, fig. 3 y fig. 4, se muestran gráficamente dentro de cada grupo obtenido con el análisis de clasificación, las variantes de plantas de acuerdo a los rasgos analizados. En el grupo uno se tienen cinco variantes: macollas compactas representadas por Calamagrostis effusa (variante 1); macollas laxas por Rhynchospora macrochaeta (variante 2) y los bambusoides por Chusquea tessellata (variante 3), estas tres variantes hacen parte de las graminoides que son las que caracterizan el grupo, los rasgos de las dos primeras están muy relacionados, en la tercera variante se encuentran interrogantes en los rasgos de regeneración; en la variante 4 'sin forma especial' se incluyen las especies que no fue posible ubicar en las otras variantes y está representada por varios pteridofitos como Huperzia rufescens, Elaphoglossum engelii y Melpomene moniliformis; la variante 5 conformada por arbustos como Hypericum juniperinum; el análisis de clasificación por altura y hábito dejó las dos últimas variantes en este grupo, cuyas especies no son dominantes en la comunidad.
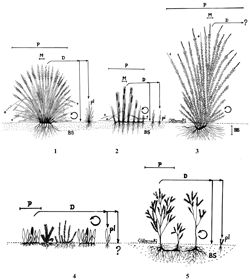
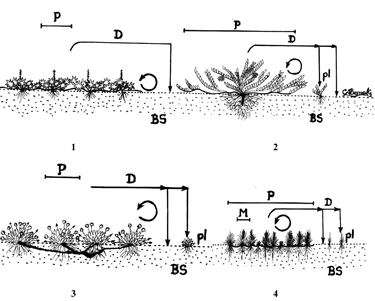
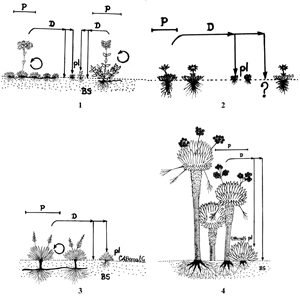
En el grupo dos se tienen cuatro variantes (principalmente en hierbas): los cojines o tapetes representados por hierbas como Lachemilla orbiculata y Arcytophyllum muticum (variante 1) y arbustos como Pernettya prostrata (variante 2); las rosetas que forman cojines o tapetes representados por hierbas como Paepalanthus dendroides (variante 3); las graminoides que forman cojines o tapetes, cuya especie tipo es Oreobolus venezuelensis (variante 4). En el grupo tres se tienen cuatro variantes: las rosetas que no forman cojines o tapetes representadas por hierbas como Gentianella corymbosa, Halenia adpressa y Bartsia santolinifolia (variante 1); las rosetas solitarias por hierbas como Hypochaeris radicata (variante 2); las graminoides en roseta representadas por Luzula gigantea (variante 3) y las caulirrosulas como Espeletia killipii (variante 4).
Estructura de los grupos de especies con rasgos de historia de vida semejantes de acuerdo con el grado de alteración
Con el test de Chi-cuadrado, se encontró que la cobertura entre los sitios es diferente (X2 = 13742.5047, p < 0.0001) para cada uno de los grupos de especies definidos con el análisis de clasificación. Excluyendo el sitio con alteración alta, también se encontraron diferencias significativas (X2 = 945.8941, p < 0.0001) de la abundancia-cobertura entre los sitios para cada grupo.
Mediante el test de proporciones entre pares de sitios, se encontraron diferencias significativas (p <0.05) en la abundancia-cobertura en los tres grupos para el sitio de alteración alta con los otros sitios o grados de alteración (p <<0.0001). Además se encontraron diferencias significativas entre el sitio sin alteración antrópica con los sitios de alteración media y leve para el grupo uno (p <<0.0001); entre todos los sitios para el grupo dos (p <<0.0001); entre los sitios con alteración media y leve (p =0.002) para el grupo tres y entre los sitios de alteración leve y sin alteración (p =0.006). Indudablemente, el grupo dos es característico del sitio con alteración alta en número de especies y cobertura; el 33 % (44 % de cobertura) de sus especies son de este sitio. El grupo uno, es característico de los sitios con alteración media, leve y sin alteración; tiene el 31% de sus especies en el sitio con alteración media mientras que el 33% de su cobertura es del sitio con alteración leve. En el grupo tres, el 30 % de sus especies se registraron en el sitio sin alteración antrópica y el 31 % de su cobertura en el sitio con alteración media; es baja su abundancia en el sitio con alteración alta (Tabla 4).

Las especies del grupo uno con mayor cobertura en el sitio con alteración alta son: Paspalum bonplandianum, Carex bonplandii y Muhlenbergia cleefii; en el sitio con alteración media: Chusquea tessellata, Calamagrostis effusa y Agrostis foliata; con alteración leve: Chusquea tessellata, Calamagrostis effusa y Rhynchospora macrochaeta y sin alteración antrópica: Chusquea tessellata y Calamagrostis effusa. Del grupo dos en el sitio con alteración alta: Lachemilla orbiculata, Plagiocheilus solivaeformis y Eleocharis stenocarpa; con alteración media: Paepalanthus dendroides y Arcytophyllum muticum, con alteración leve: Pernettya prostrata, Paepalanthus dendroides y Geranium santanderensis; sin alteración antrópica: Paepalanthus dendroides y Pernettya prostrata. Del grupo tres en el sitio con alteración alta: Gamochaeta sp, Plantago australis y Rumex acetosella; con alteración media: Bartsia santolinaefolia, Halenia adpressa y Blechnum loxense; con alteración leve: Espeletia killipii, Halenia adpressa y Bartsia santolinaefolia y sin alteración antrópica: Espeletia killipii, Gentianella corymbosa, Halenia adpressa y Bartsia santolinaefolia.
DISCUSIÓN
Grupos de especies con rasgos de historia de vida similares
Varias de las especies que sólo se registraron en el sitio con alteración alta (50 %), son típicas de sitios sometidos a pastoreo como Rumex acetosella, Trifolium repens, Anthoxanthum odoratum, Bidens triplinervia, Lachemilla orbiculata y Agrostis boyacensis, datos que los confirman varios trabajos (Lozano & Schnetter 1976, Franco et al. 1986, Sánchez et al.1989, Verweij & Budde 1992, Verweij 1995, Premauer 1999 y Jaimes 2000); actualmente la comunidad vegetal en este sitio está conformada por especies diferentes a las presentes en los otros sitios, los cuales comparten la mayoría de las especies.
Aunque en general para cada grupo predomina un atributo de cada rasgo, es necesario tener en cuenta la heterogeneidad en los grupos cuando se analiza su comportamiento frente a la alteración; McIntyre et al. (1999 b), afirman que al realizar el análisis de todos los rasgos posiblemente se oscurezcan patrones asociados únicamente con algunos grupos mayores y proponen analizar los rasgos dentro de las formas principales. Una razón de la falta de diferencias claras entre los grupos en relación con los rasgos de regeneración posiblemente sea las características ambientales del páramo junto con la falta de conocimiento sobre algunos de estos rasgos en las especies.
Las especies registradas en los cuatro sitios son policárpicas perennes con una gran diversidad de formas de crecimiento; este rasgo es muy importante para la existencia de las especies bajo las condiciones particulares del páramo, independiente de la existencia o no de disturbio en esta comunidad. El estudio biotipológico de Cuatrecasas (1968) en cinco comunidades de páramo, muestra que las plantas herbáceas son perennes y las plantas anuales no están o son extremadamente raras. Esta observación concuerda con lo registrado por Billings (1974) para las plantas de la tundra, donde en su mayoría son herbáceas perennes, con grandes raíces o tallos/rizomas como sistema de almacenaje, son muy pocas las especies bianuales y muy raras las anuales.
Las especies con expansión vegetativa, como las macollas compactas de las gramíneas (Calamagrostis effusa) y las macollas laxas de las ciperáceas (Rhynchospora macrochaeta), o de hierbas como Bartsia, Halenia y Gentianella florecen a medida que se van desarrollando, fructifican y luego mueren, pero la planta sigue viva; es decir que a nivel del individuo cada ramet o vástago es monocárpico pero el individuo como tal es policárpico.
Por lo general Chusquea tessellata se considera como una especie monocárpica, en la comunidad estudiada, se pudo observar que muchos de los vástagos de cada bambusoide florecieron y formaron espigas, pero no formaron frutos; según Clark (1989), algunas especies de bambusoides pueden mostrar floración esporádica sin producción de semillas en los intervalos entre los episodios de la floración gregaria; esto puede estar ocurriendo con la población de Chusquea tessellata en el valle del río Tunjo. En otro valle cerca al área de estudio, igualmente se observó la muerte masiva de una población de esta misma especie, en la cual, O. Vargas (com. pers.) estimó que aproximadamente el 20 % de los individuos o clones estaban rebrotando a partir de sus rizomas, como se ha encontrado en otras especies de bambúes.
Dadas las condiciones drásticas del páramo y el disturbio por quema y pastoreo, la existencia de reproducción sexual y vegetativa en la mayoría de especies podría estar favoreciendo la permanencia de sus poblaciones. Mientras que la falta de reproducción vegetativa en algunas especies, como Espeletia killipii y Greigia stenolepis, puede conllevar a que sus poblaciones no respondan de igual manera ante cambios producidos en el ambiente y finalmente desaparezcan, aún más teniendo en cuenta que el establecimiento de las plántulas, es un proceso limitante en la dinámica poblacional de varias especies de las zonas de alta montaña, como se encontró para varias especies del género Espeletia (Monasterio 1979, Fagua 2002).
Es interesante e importante considerar en futuros estudios, la baja o aparente falta de producción de frutos de algunas especies, teniendo en cuenta que en varias especies de pastos se ha registrado viviparidad y que en Chusquea tessellata se observó la producción de estructuras diferentes a frutos en las espigas. Según Lee & Harmer (1980) y Latting (1972), la viviparidad en gramíneas ayuda a mantener la especie y acelera la reproducción en condiciones adversas a la reproducción sexual, por este medio los pastos son capaces de existir bajo condiciones ambientales extremas que de otro modo no podrían soportar. La presencia de rosetas, densos cojines y macollas son comunes en ambientes en los cuales existen cambios bruscos de temperatura, fuertes heladas, fuertes vientos y alta radiación solar, entre otros; indudablemente estas formas son importantes en especies en los cuatro sitios estudiados.
A pesar de que no es la misma comunidad y que existen diferencias en el criterio para formar los grupos, existen algunas equivalencias de nuestros resultados con respecto a los presentados por Verweij (1995), quien define seis grupos funcionales de plantas para los pastizales de páramo en relación a quema y pastoreo: las graminoides erectas en macolla como Calamagrostis efusa, Festuca sp., Agrostis boyacensis y Agrostis breviculmis que en este trabajo hacen parte del grupo uno, la autora los separa en dos grupos: pastos en macolla y pastos cortos.
En nuestro caso las rosetas caulescentes que están representadas por especies como Espeletia killipii y Blechnum loxense y las rosetas rasantes por especies como Hypochaeris sessiliflora, hacen parte del grupo tres donde las rosetas se segregan por altura y por no presentar regeneración vegetativa, la autora separa las rosetas caulescentes de las rasantes y en estas últimas incluye especies con hábito postrado, menores de 5 cm de altura, sin importar que no sean rosetas y mezcla hierbas y arbustos; el grupo dos obtenido en este trabajo, conformado por especies que forman cojines o tapetes y/o que son rastreras o postradas, haría parte del grupo de rosetas rasantes dado por la autora. Finalmente el análisis incluye los arbustos (representados por pocas especies en la comunidad estudiada) dentro de los tres grupos de acuerdo a sus atributos, principalmente de hábito y altura, mientras que Verweij (1995) deja los grupos hierbas (mayores de 5 cm de altura) y arbustos (no incluye las especies de hábito postrado).
Estructura de los grupos de especies con rasgos de historia de vida semejantes de acuerdo con el grado de alteración
De acuerdo con los resultados, indudablemente se tiene una tendencia de variación en la abundancia de cada uno de los grupos de especies con rasgos similares, en relación al grado de alteración. En general cada grupo, presenta su máxima abundancia en un sitio de acuerdo con el grado de alteración. Según McIntyre et al. (1999 b), la persistencia de las especies bajo un régimen de disturbios como quema y pastoreo se puede considerar como una expresión de varios rasgos que en combinación permiten a la planta persistir, evitando el peligro en estados críticos o tolerando y permitiendo el recrecimiento o reclutamiento. El grupo uno representado principalmente por especies graminoides, erectas en macolla o bambusoides, disminuye su abundancia al aumentar el grado de alteración. Las diferencias que se presentan en la abundancia del grupo en los diferentes sitios, se deben a la disminución de las especies dominantes Chusquea tessellata y Calamagrostis effusa cuando aumenta el grado de alteración. En el sitio con mayor alteración también se encuentran especies de este grupo, pero se caracterizan por ser en su mayoría pastos cortos y registrados solo para este sitio.
El grupo dos representado principalmente por hierbas que forman cojines o tapetes, rastreras o postradas y con alturas entre 0 y 5 cm, es más abundante en el sitio con mayor alteración, las especies más altas no son muy abundantes en este sitio; el efecto del disturbio, por quema y pastoreo sobre el tamaño de la planta concierne no solo a la altura sino también a la expansión lateral, con un incremento de especies con amplia capacidad de expansión lateral. La tendencia de variación de las rosetas características del grupo tres, no es tan marcada como la de los otros grupos, sin embargo es indudable que la abundancia de este grupo también cambia con el grado de alteración, presentando su menor abundancia en el sitio con alto grado de alteración donde la mayoría de especies son rasantes; en este grupo se tienen las caulirrosulas, representadas principalmente por Espeletia killipii que puede alcanzar grandes alturas, esta especie característica de la comunidad natural (sitios sin alteración antrópica), es afectada negativamente, no se encuentra en los sitios con alteración media y alta, donde el grupo es bien representado por otras especies de rosetas de menor altura, razón por la cual no se encontraron diferencias significativas en la abundancia de las rosetas entre estos sitios.
Aunque se tienen las anteriores tendencias, especies rastreras o postradas que forman cojines o tapetes también se encuentran en los sitios con grado de alteración muy bajo, esta forma junto con las rosetas son características del páramo; probablemente pocas especies tienen atributos relacionados directamente para sobrevivir a procesos actuales de alteración, pero muchos de los atributos que persisten a través de filtros climáticos y de suelo pueden a la vez conferir alguna resistencia a diferentes disturbios. Por ejemplo, un número de atributos de las plantas para evadir, resistir o tolerar extensos periodos de sequía, proveen beneficios similares a las plantas sujetas a pastoreo o quema, esto incluye meristemo basal, pequeña estatura y alto potencial de crecimiento (Coughenour 1985 citado en Landsberg et al. 1999).
En la comunidad estudiada se pueden identificar especies sensibles al régimen de disturbio por quema y pastoreo al que ha sido sometida, dentro de estas podríamos ubicar especies de los grupos uno y tres, como las graminoides Calamagrostis effusa y Chusquea tessellata las cuales van disminuyendo su abundancia-cobertura hasta desaparecer con una alteración alta y la caulirrósula Espeletia killipii, localmente extinta con alteración media; igualmente se pueden incluir otras especies cuya abundancia es mayor con alteración media pero cuando esta es alta se extinguen como las rosetas Halenia adpressa y Bartsia santolinaefolia. Especies tolerantes al disturbio que hacen parte del grupo dos, la mayoría de las cuales son características del sitio con alto grado de alteración, y registradas en otros trabajos (Verweij y Budde 1992, Verweij 1995, Jaimes 2000), como tolerantes al pastoreo entre las cuales se encuentran Lachemilla orbiculata, Trifolium repens, Paspalum bonplandianum y Bidens triplinervia entre otras. Finalmente un pequeño grupo de especies que se encuentran bien representadas tanto en los sitios con alteración alta como sin alteración antrópica, entre estas tenemos a Geranium sibbaldioides y Arcytophyllum muticum.
AGRADECIMIENTOS
Expresamos nuestros agradecimientos a COLCIENCIAS y a la Universidad Nacional de Colombia por la financiación, como parte del proyecto 'Sucesión regeneración del páramo después de quema y pastoreo'. (Código: 1101-13-607-96). A la Unidad Administrativa Especial de Parques Nacionales Naturales y al Parque Nacional Natural Chingaza. A los botánicos del COL que colaboraron en la determinación del material. A los biólogos: Alba L. León, Emil Hernández, Yolima Martínez, Hernán Sánchez y Gerardo Arenas por su colaboración en labores de campo. Al profesor Santiago Grillo por la asesoría estadística. A los profesores Olga V. Castaño, Silvio Zuluaga y Orlando Rangel-Ch. por sus comentarios y sugerencias. A Carlos Bernal por la elaboración de los dibujos. A Mario Vargas y Jimena Cortés por la revisión del manuscrito. A los dos revisores anónimos por sus valiosos aportes y críticas al manuscrito original.
LITERATURA CITADA
1. BILLINGS, W.D. 1974. Arctic and alpine vegetation: Plant adaptations to cold summer climates. Pp 403-443. In: D. Ives & R. Barry. Arctic and alpine environments. [ Links ]
2. CAMPBELL, B.D., S. STAFFORD, D. MARK & A.J. ASH. 1999. A rule-based model for the functional analysis of vegetation change in Australasian grasslands. Pp 723 - 730. In: S. Lavorel & W. Cramer (eds). Plant functional types and disturbance dynamics. Journal of vegetation Science 10(5): 603-762. [ Links ]
3. CÁRDENAS, C., C. POSADA & O. VARGAS. 2002. Banco de semillas germinable de una comunidad vegetal de páramo húmedo sometido a quema y pastoreo (Parque Nacional Natural Chingaza, Colombia). Ecotrópicos 15 (1): 49-58. [ Links ]
4. CLEEF, A.M. 1981. The vegetation of the Paramos of the Colombian Cordillera Oriental. Tesis de Doctorado, Universidad de Utrecht, Utrecht. [ Links ]
5. CLARK, L.G. 1989. Systematics of Chusquea Section Swallenochloa, Section Verticillatae, Section Serpentes, and Section Longifoliae (Poaceae-Bambusoideae). Systematic botany monographs. The American Society of Plant Taxonomists 27: 1-127. [ Links ]
6. CUATRECASAS, J. 1968. Paramo vegetation and its life forms. Pp 163-186. In: C. Troll (ed.): 'Geo-ecology of the mountains regions of the Tropical Americas'. Colloquium Geographicum 9. [ Links ]
7. DÍAZ, B.M.C., M. ZUNZUNEGUI, R. TIRADO, F. AIN-LHOUT & F. GACÍA. 1999. Plant functional types and ecosystem function in Mediterranean shubland. Pp 709-716. In: S. Lavorel & W. Cramer (eds). Plant functional types and disturbance dynamics. Journal of vegetation Science 10(5): 603- 762. [ Links ]
8. DÍAZ, S. & M. CABIDO. 1997. Plant functional types and ecosystem function in relation to global change. Journal of Vegetation Science 8: 463-474. [ Links ]
9. DÍAZ, S., M. CABIDO, M. ZAK, E. MARTÍNEZ & J. ARANIBAR. 1999. Plant functional traits, ecosystem structure and land-use history along a climatic gradient in central-western Argentina. Pp 651- 660. In: S. Lavorel & W. Cramer (eds). Plant functional types and disturbance dynamics. Journal of vegetation Science 10(5): 603- 762. [ Links ]
10. EAAB. 1997. Términos de referencia para el estudio de impacto ambiental y plan de manejo del proyecto Chingaza II. [ Links ]
11. FAGUA, G.C. 2002. Estrategias reproductivas de reproducción sexual de una población de Espeletia grandiflora H & B, en el parque Nacional Natural Chingaza (Cundinamarca). Trabajo de grado, departamento de Biología, Universidad Nacional de Colombia, Bogotá. [ Links ]
12. FRANCO, P. 1982. Estudios fitoecológicos en el parque Nacional Natural Chingaza. Trabajo de grado, Departamento de Biología, Universidad Nacional de Colombia, Bogotá. [ Links ]
13. FRANCO, P., O. RANGEL-CH. & G. LOZANO-C. 1986. Estudio ecológico en la cordillera Oriental II: Las comunidades vegetales de los alrededores de la laguna de Chingaza (Cundinamarca). Caldasia 15(71-75): 219-248. [ Links ]
14. GITAY, H. & I.R. NOBLE. 1997. What are functional types and how should we seek them?. Pp 3-19. In: T.M. Smith, H.H. Shugart and F.I. Woodward (eds). PLANT FUNCTIONAL TYPES. Their relevance to ecosystem properties and global change. International Geosphere-Biosphere Programme Book Series 1. [ Links ]
15. GRIME, J.P. 1979. Plant Strategies and vegetation processes. John Wiley & Sons. Londres. 222 pp. [ Links ]
16. GRIME, J.P. 1989. Estrategias de adaptación de las plantas y procesos que controlan la vegetación. Editorial LIMUSA. México, D.F. 291 pp. [ Links ]
17. HALPERN, C.B. 1989. Early successional patterns of forest species: interactions of life history traits and disturbance. Ecology 70 (3): 704-720. [ Links ]
18. HOFSTEDE, R. 1995. Effects of burning and grazing on a Colombian Páramo ecosystem. Universidad de Amsterdam. ICG. Holanda. 196 pp. [ Links ]
19. HOFSTEDE, R., J. LIPS, W. JONGSMA & Y. SEVINK. 1998. Geografía, ecología y forestación de la Sierra Alta del Ecuador. Revisión de literatura. ed. Abya-Yala. 242 pp. [ Links ]
20. JAIMES, V. 2000. Estudio ecológico de una sucesión secundaria y mecanismos de recuperación de la fertilidad en un ecosistema de páramo. Tesis Posgrado. Universidad de los Andes de Mérida. Venezuela. [ Links ]
21. KLEYER, M. 1999. Distribution of plant functional types along gradients of disturbance intensity and resource supply in an agricultural landscape. Pp 697-708. In: S. Lavorel & W. Cramer (eds). Plant functional types and disturbance dynamics. Journal of vegetation Science 10 (5): 603-762. [ Links ]
22. LANDSBERG, J., S. LAVOREL & J. STOL. 1999. Grazing response groups among understorey plants in arid rangelands. Pp 683 - 696. In: S. Lavorel & W. Cramer (eds). Plant functional types and disturbance dynamics. Journal of vegetation Science 10 (5): 603-762. [ Links ]
23. LATTING, J. 1972. Differentiation in the grass inflorescence. In: Youngner, V. B. & C.M. McKell (eds). The biology and utilization of grasses. Academic Press, Londres. Pp 365-399. [ Links ]
24. LAVOREL, S., S. MCINTYRE & K. GRIGULIS. 1999. Plant response to disturbance in a Mediterranean grassland: How many functional groups?. Pp 661- 672. In: S. Lavorel & W. Cramer (eds). Plant functional types and disturbance dynamics. Journal of vegetation Science 10 (5): 603- 762. [ Links ]
25. LEE, J.A. & R. HARMER. 1980. Vivipary, a reproductive strategy in response to environmental stress?. Oikos 35: 254-265. [ Links ]
26. LOZANO-C., G. & R. SCHNETTER. 1976. Estudios ecológicos en el páramo de Cruz Verde, Colombia II. Las comunidades vegetales. Caldasia 2(54):53-68. [ Links ]
27. MCINTYRE, S. & R.M. TREMONT 1995. Plant life - history attributes: their relationship to disturbance response in herbaceous vegetation. Journal of Ecology 83: 31-44. [ Links ]
28. MCINTYRE, S., S. DÍAZ, S. LAVOREL & W. CRAMER. 1999 a. Plant functional types and disturbance dynamics. Introduction. Pp 604-608. In: S. Lavorel & W. Cramer (eds). Plant functional types and disturbance dynamics. Journal of vegetation Science 10 (5): 603-762. [ Links ]
29. MCINTYRE, S., S. LAVOREL, J. LANDSBER & T.D.A. FORBES. 1999 b. Disturbance response in vegetation - towards global perspective on functional traits. Pp 621- 630. In: S. Lavorel & W. Cramer (eds). Plant functional types and disturbance dynamics. Journal of vegetation Science 10 (5): 603- 762. [ Links ]
30. MONASTERIO, M. 1979. El páramo desértico el altiandino de Venezuela. El medio ambiente Páramo. Pp 117-146. En: M. L. Salgado-Labouriau (ed). Entro de estudios avanzados, IVIC. Caracas. [ Links ]
31. MONASTERIO, M. 1980. Los páramos andinos como región natural. Características biogeográficas generales y afinidades con otras regiones andinas. En: M. Monasterio (ed). Estudios ecológicos en los páramos andinos. Ediciones de la Universidad de los Andes. Mérida. Venezuela. Pp 15-27. [ Links ]
32. NOBLE, I.R. & R.O. SLATYER. 1980. The use of vital attributes to predict successional changes in plant communities subject recurrent disturbances. Vegetatio 43: 5-21. [ Links ]
33. PÁEZ, V. 2002. Comunidades vegetales de páramo en un valle quemado y pastoreado (Parque Nacional Natural Chingaza). Trabajo de grado, Departamento de Biología, Universidad Nacional de Colombia, Bogotá. [ Links ]
34. PELS, B. & P.A. VERWEIJ. 1992. Burning and grazing in a bunchgrass Paramo ecosystem: Vegetation dynamics described by a transition model. Pp 243-263. In: H. Balslev & J. L. Luteyn (eds). Páramo, an Andean Ecosystem under human influence. Academic Press. Londres. [ Links ]
35. PICKETT, S.T., J. KOLASA, J.J. ARMENSO & S.L. COLLINS. 1989. The concept of ecological disturbance and its expression at various hierarchical levels. OIKOS 54: 129-136. [ Links ]
36. POSADA, C. & C. CÁRDENAS.1999. Banco de semillas germinable de una comunidad vegetal de páramo sometida a quema y pastoreo (Parque Nacional Natural Chingaza). Trabajo de grado, departamento de Biología. Universidad Nacional de Colombia, Bogotá. [ Links ]
37. PREMAUER, M.J.M. 1999. Efecto de diferentes regímenes de disturbio por quema y pastoreo sobre la estructura horizontal y vertical de la vegetación de Páramo (Parque Nacional Natural Chingaza). Trabajo de grado, departamento de Biología, Universidad Nacional de Colombia, Bogotá. [ Links ]
38. RANGEL-Ch., J.O. 2000. Clima de la región paramuna. Pp 85-125. En: J. O. Rangel-Ch. (ed). La región de vida paramuna. Colombia diversidad Biótica III. Universidad Nacional de Colombia, Facultad de Ciencias, Instituto de Ciencias Naturales. Bogotá. [ Links ]
39. RANGEL-CH., J.O. & C. ARIZA-N. 2000. La vegetación del parque Nacional Natural Chingaza. Pp 720-753. En: J. O. Rangel-Ch. (ed). La región de vida paramuna. Colombia diversidad Biótica III. Universidad Nacional de Colombia, Facultad de Ciencias, Instituto de Ciencias Naturales. Bogotá. [ Links ]
40. RYDIN, H. & S.O. BORGEGARD. 1991. Plant characteristics over a century of primary succession on islands: Lake Hjalmaren. Ecology 72 (3): 1089-1101. [ Links ]
41. SÁNCHEZ-M., R., J.O. RANGEL-CH. & J. AGUIRRE-C. 1989. Estudios ecológicos en la cordillera Oriental IV: Aspectos ginecológicos de la brioflora de los depósitos turbosos paramunos de los alrededores de Bogotá. Caldasia 16(76): 41-57. [ Links ]
42. SAS/STAT User Guide 1990. Volumen 2, Version 6, cuarta edición. [ Links ]
43. SOKAL, R. & J. ROHLF. 1979. Biometría. Principios y métodos estadísticos en la investigación biológica. Ed. Blume. 832 pp. [ Links ]
44. STURM, H. & O. RANGEL-CH. 1985. Ecología de los páramos andinos: una revisión preliminar integrada. ICN-MHN. UN. Bogotá. [ Links ]
45. VAN DER HAMMEN, T & A.M. CLEEF. 1986. Development of the high Andean Páramo flora and vegetation. In: F. Vuilleumier & M. Monasterio (eds). High Altitude Tropical Biogeography. Oxford University Press. Pp 153-201. [ Links ]
46. VARGAS, O. 1997. Un modelo de sucesión-regeneración de los páramos después de quema. Caldasia 19 (1-2): 331-345. [ Links ]
47. VARGAS, O., J. PREMAUER & C. CÁRDENAS. 2002. Efecto del pastoreo sobre la estructura de la vegetación en un páramo húmedo de Colombia. Ecotrópicos 15(1): 33-48. [ Links ]
48. VERWEIJ, P. & P.E. BUDDE. 1992. Burning and grazing gradients in Páramo vegetation: Initial ordination analyses. Pp 177-196. In: H. Balslev & J. L. Luteyn (eds). Páramo, an Andean Ecosystem under human influence. Academic Press. Londres. [ Links ]
49. VERWEIJ, P. 1995. Spatial and temporal modeling of vegetation patterns. Burning and grazing in the paramo of Los Nevados National Park, Colombia. ITC, Holanda. 233 pp. [ Links ]
50. YOUNG, F.W., Y. TAKANE & J. LEEUW. 1978. The Principal Components of Mixed Measurement Level Multivariate Data: An alternanting least squares method with optimal scaling features. Psychometrika 43: 279-281. [ Links ]
51. ZULUAGA, S. 2002. Estructura de dos comunidades de frailejón Espeletia grandiflora Kunth y Espeletia killipii Cuatrec., en laderas y valles de la quebrada de Piedras Gordas, Parque Nacional Natural Chingaza. Tesis de Maestría. Departamento de Biología. Universidad Nacional de Colombia, Bogotá, D.C. [ Links ]
Recibido: 24/08/2006
Aceptado: 14/05/2008

