










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.30 no.2 Bogotá July/Dec. 2008
JAVIER RODRÍGUEZ-BARRIOS
RODULFO OSPINA-TORRES
JUAN DAVID GUTIÉRREZ
Laboratorio de Invertebrados Acuáticos, Universidad Nacional de Colombia. Bogotá D. C., Colombia. jarodriguezba@gmail.com; rospinat@unal.edu.co; jdgutierrezt@unal.edu.co
RESUMEN
Se determinó la cantidad y la variación de las entradas laterales, verticales y el transporte de materia orgánica particulada (MOP) durante un período de cinco meses en un tramo de una quebrada tropical de montaña, ubicada en los cerros orientales de Bogotá – Colombia, haciendo énfasis en el efecto de la magnitud de la precipitación y el caudal, sobre las entradas y el transporte de MOP. Las entradas verticales de MOP gruesa (MOPG vertical) y laterales de MOPG (MOPG lateral) y MOP fina (MOPF lateral) correspondientes a los cinco meses de muestreo fueron 450.8 g.m-2, 191 g.m-1 y 12.65 g.m-1 de peso seco libre de cenizas (PSLC) respectivamente. La MOPF lateral mostró una tendencia clara a asociar su incremento con la intensidad de las precipitaciones, como sucedió con la MOPG vertical pero con valores atípicos que afectaron su correspondencia estadística. El transporte promedio de MOPG y MOPF para períodos de baja descarga fue de 0.44 g.m-3 y 1.13 g.m-3 y durante períodos de alta descarga fue de 0.102 g.m-3 y 3.1 g.m-3 respectivamente, siendo el transporte de MOPF 30 veces mayor al de MOPG para los cinco meses de muestreo. Se presentaron diferencias en la MOPF transportada por el cauce durante eventos de alto y bajo caudal. No se presentó una asociación estadística entre la magnitud de la descarga y el transporte de las fracciones de MOP.
Palabras clave. Materia orgánica alóctona, Precipitación, Neotrópico, Quebradas andinas de montaña.
ABSTRACT
Quantity and variation of inputs -lateral and vertical- and transport of particulate organic matter (POM) were estimated over a five month- period in a section of a tropical mountain stream, in the eastern hills of Bogotá, Colombia, with emphasis on the effects of precipitation and flow over inputs and transport of POM. Vertical inputs of coarse POM (vertical CMOP) and lateral inputs of CPOM (lateral CPOM) and fine POM (lateral FPOM ) were 450.8 g.m-2 , 191 g.m-1 and 12.65 g.m-1 of ash free dry matter (AFDM), respectively. Lateral FPOM showed a clear tendency to increase with the intensity of precipitation; the same tendency appeared inthe vertical CPOM but atypical values affected its statistical correspondence. Transport of CPOM and MOPF during low discharge periods was 0.44 g.m-3 and 1.13 g.m-3 respectively and 0.102 g.m-3 and 3.1 g.m-3 during high discharge periods. FPOM was 30 times greater than CCPOM over the five months of sampling. Differences in FPOM transport by the stream channel were observed between low and high discharge periods. No statistical relationship was evident between discharge magnitude and POM transport fractions.
Key words. Organic Matter, Precipitation, Neotropics, Andean mountain stream.
INTRODUCCIÓNPequeños ríos representados por Quebradas de primer orden, reciben la materia orgánica particulada (MOP) alóctona proveniente de la vegetación riparia como la mayor fuente de energía para sus comunidades acuáticas (Anderson & Sedell 1979, Cummins et al. 1983, Minshall et al. 1985). El funcionamiento de las Quebradas depende de su grado de interacción bosque-río (Minshall et al. 1985, Ward 1986, Hassan et al. 2005). Cuando la MOP entra a las quebradas una fracción es fragmentada o degradada y otra es transportada aguas abajo (Webster et al. 1999). La importancia relativa de estos procesos varía fuertemente con el orden del río (Webster & Benfield 1986). El transporte de materiales en forma de detrito, depende del tamaño y la densidad de las partículas que son movilizadas y de las características físicas de la quebrada, incluida la profundidad, la presencia de estructuras retentivas y la intensidad del caudal (Webster et al. 1999).
La materia orgánica particulada gruesa (MOPG) es generada por la vegetación riparia, a través de procesos físicos o fisiológicos, exportando al interior de la quebrada sus hojas, ramas, flores y otros componentes (Webster et al. 1999, Richardson et al. 2005). También pueden presentarse procesos biofísicos que intervienen en la floculación de la materia orgánica particulada fina (MOPF) para la producción de MOPG (Paul & Hall, 2002). Cuando esta MOPG ingresa a la quebrada, se moviliza algunos metros en un flujo unidireccional debido a la fuerza del caudal (Vannote et al. 1980), hasta encontrar obstáculos en el canal que detienen su movimiento, promoviendo procesos de fragmentación y degradación biológica por colonización microbiana y el posterior consumo por macroinvertebrados trozadores (Cummins & Klug 1979). De esta forma la MOPG se constituye en la mayor fuente energética para quebradas de cabecera (Cummins 1974).
La MOPF Alóctona es generada por procesos físicos en la cabecera de los ríos, ya sea por degradación de la MOPG, por la floculación de la MOD (Paul & Hall 2002) o por la acción combinada de procesos físicos, químicos y biológicos (Webster et al. 1999) y puede ingresar al cauce debido a su escorrentía desde las laderas en la interfase bosque-río. Esta MOPF se constituye junto a la MOPG en fuente esencial de energía para las comunidades ubicadas en el curso inferior de los ríos (Paul & Hall 2002).
La importancia de la zona riparia como fuente generadora de MOPG y MOPF es afectada de acuerdo con el orden del río (Vannote et al. 1980), predominando la MOPG en ríos de orden menor (orden: 1-3). La MOPF se presenta en mayor proporción en sectores del río de orden superior en forma de materia en suspensión o almacenada y transportada en el lecho del río (Cushing et al. 1993, Vannote et al. 1980). Tabacchi et al. (1998) demostraron que la vegetación riparia juega un papel importante como fuente y sumidero de materia y energía, en el contexto de sus atributos estructurales y funcionales de acuerdo con procesos de ciclado de nutrientes, generación de MOP, retención de agua y difusión o dispersión de su biota.
Eventos hidroclimáticos como las crecientes e inundaciones promueven la fragmentación física de la MOP por agregaciones de detritos y una mayor superficie expuesta a la fragmentación biológica oxidativa (Hauer & Lamberti 1996). Procesos simultáneos de transporte y fragmentación de la MOP son considerados por Webster & Patten (1979) en sus trabajos con “espiralamiento de las quebradas”, destacando la eficiencia en la utilización de esta MOP en los procesos mencionados de transporte y retención. De acuerdo con Webster et al. (1999), la distancia de transporte de partículas de MOP puede ser modelada de acuerdo con la siguiente ecuación:
dx/dt = kXX = - SXX
Donde dx/dt es la tasa de cambio de X con respecto a la distancia; X corresponde a la cantidad de material que es movilizado; kX corresponde a una tasa exponencial negativa que representa a las pérdidas de material por retención en relación a una concentración inicial que es liberada. El inverso de kX representa la distancia promedio de transporte (SX). Este modelo ha sido utilizado para la deriva de insectos (Elliot 1971), transporte de hojas (Cummins et al. 1989) y el transporte de trozos de madera (Ehrman & Lamberti 1992).
De acuerdo con Fisher & Likens (1973) las entradas de material alóctono a una Quebrada, se encuentran dominando los procesos de la dinámica de la MOP, lo cual puede ser modelado de acuerdo con la siguiente ecuación:
dx/dt = I – kX – k'X
Donde dx/dt es la tasa de cambio de X con respecto al tiempo; X corresponde a la cosecha de MOP (masa o energía por unidad de área); I es la entrada de MOPG (masa o energía por unidad de área); k es el exponente negativo del modelo; k'X corresponde a la tasa exponencial de pérdidas por transporte aguas abajo.
El entendimiento de los disturbios que afectan la dinámica de las entradas y el transporte de MOP en las quebradas, puede ser utilizado en estudios de manejo y restauración de estos sistemas (Bilby & Bisson 1992, Karlsson et al. 2005). Se tiene conocimiento del efecto que tiene la reducción de la vegetación riparia en la alteración de la cosecha y el transporte de MOPG para las quebradas, en adición al efecto perturbador sobre los grupos funcionales de invertebrados (Webster et al. 1999, Bilby & Bisson 1992). Disturbios hidroclimáticos como las tormentas y las crecientes, muestran un efecto puntual y cíclico en el transporte y almacenamiento de MOP (Richardson et al. 2005).
Eventos hidroclimáticos de fuerte intensidad pueden afectar las propiedades estructurales y funcionales de las Quebradas, modificando además la cantidad, tipo y tiempo en la entrada de MOP y su posterior transporte en las quebradas (Reeves et al. 1995). Se sabe que en regiones del Pacífico holártico, la biota puede adaptarse a la estacionalidad de las entradas de esta MOP alóctona, acondicionando sus eventos fenológicos de acuerdo a las fluctuaciones en la oferta de este alimento y la generación de refugios por la entrada de grandes cantidades de palizada al lecho de las Quebradas durante eventos de lluvia (Richardson et al. 2005).
La condición tropical que identifica a la Quebrada La Vieja, promueve procesos que se diferencian de las zonas templadas, debido a la estabilidad de factores como la intensidad de la radiación solar, la temperatura anual, el caudal, el patrón de las precipitaciones, las entradas y el transporte de MOPG.
El presente estudio intenta hacer una cuantificación de las entradas de MOP procedente del bosque ripario durante eventos de lluvia y sequía, así como la MOP que es transportada a escala de tramo en una quebrada tropical de montaña, durante eventos de alto y bajo caudal, con el fin de confrontar los resultados con otros estudios, que han sido desarrollados en otras regiones del mundo. En este sentido, se pretende tener herramientas que evalúen el grado de interacción bosque-río en una Quebrada de montaña bien conservada. Con base en lo anterior, se desea abordar las siguientes hipótesis: (1) a nivel de escala de tramo, la dinámica de las entradas a la Quebrada de diferentes fracciones y fuentes de MOP, es regulada por eventos de lluvia y sequía. En este sentido se presentará un umbral de precipitación de suficiente intensidad para generar un incremento significativo en las entradas de MOPG y MOPF al cauce de la Quebrada, (2) a nivel de escala de tramo la intensidad del caudal presenta una relación directa con la magnitud del transporte de MOP en sus fracciones fina y gruesa.
METODOLOGÍA
Sitio de estudio
El estudio se realizó en la microcuenca de la Quebrada La Vieja, la cual se encuentra ubicada en la zona Nor-Oriental de la ciudad de Bogotá (74°08' W, 4°36' N) y es un afluente de la macrocuenca del río Bogotá. En dicha quebrada, las condiciones físicas corresponden a las de un río de montaña (cotas máx. 3200 y 3300 m.s.n.m, cota mín. 2690 m.s.n.m.). El régimen climático de esta quebrada, se encuentra dominado por la localización de la misma sobre los cerros orientales de la Sabana de Bogotá y su forma encañonada, mostrando una alta humedad (Humedad relativa promedio de 85%) con una distribución bimodal de las lluvias (precipitación media anual de 1100 mm). Las oscilaciones climáticas a lo largo del día, se deben al desplazamiento de las nubes y el ascenso de neblina, ocasionando variaciones repentinas y pronunciadas de la humedad relativa (Gutiérrez 2006).
Esta quebrada presenta un buen estado de conservación, complementado con condiciones “tropicales” de una temperatura del agua relativamente constante a lo largo del año (promedio de 14 ºC) y un caudal poco variable (el caudal medio es de 15.6 l.s-1), producto de una cuenca pequeña y encañonada.
Todos los suelos de la quebrada son ácidos, pobres en nutrientes, con altos contenidos de aluminio e hidrógeno, y de baja fertilidad, pero con un buen desarrollo de su vegetación (Vargas 2003). En su parte alta, la quebrada presenta suelos de la serie Páramo que corresponden a Dystrudeps Húmicos, en las laderas se encuentran suelos de la asociación Monserrate de composición Troporthents líticos de baja profundidad con afloramientos rocosos. En las zonas de menor pendiente sobre arcillolita se encuentra el suelo Cabrera, correspondiente a un Dystrudept (Cortés 1976).
Adicionalmente, la vegetación riparia se halla bien desarrollada y provee sombrío a lo largo del año. Esta vegetación es típica de bosques secundarios jóvenes (40 años), donde se encuentran especies juveniles como: encenillo (Weinmannia tomentosa), gaque (Clusia multiflora), arrayán (Myrcianthes leucoxyla) y tuno (Miconia spp.). Adicionalmente se encuentran plantaciones de especies exóticas como: eucalipto (Eucalyptus globulus), acacia negra (Acacia melanoxylon), ciprés (Cupressus sp.) y pino pátula (Pinus patula). Las partes inferior y media de la microcuenca corresponden a la clasificación de “bosque húmedo Montano Bajo” y la parte superior se clasifica como “bosque húmedo Montano” según las categorías de zonas de vida de Holdridge (Espinal & Montenegro 1963).
Escala espacio-temporal
Entradas de MOP- Se seleccionó un tramo de 200 m (aproximadamente) que resumiera la heterogeneidad de la vegetación riparia, así como la heterogeneidad geomorfológica de las laderas ubicadas en la zona que limita con la columna de agua. Se realizaron 10 muestreos entre marzo y agosto de 2005, desarrollando 2 campañas por mes, dejando las trampas instaladas para la colecta de fracciones de MOPF y MOPG durante 15 días para cada muestreo.
Transporte de MOP- En la Quebrada se seleccionó un tramo de 30 m con zonas de rápidos y remansos, tratando de resumir la heterogeneidad geomorfología que le brinda identidad física al lugar de estudio (sensu Frissell et al. 1986). Se realizaron 10 muestreos entre Abril y Agosto de 2005 (2 campañas por mes), abarcando un período de aguas altas y otro de aguas bajas (Figura 1A).
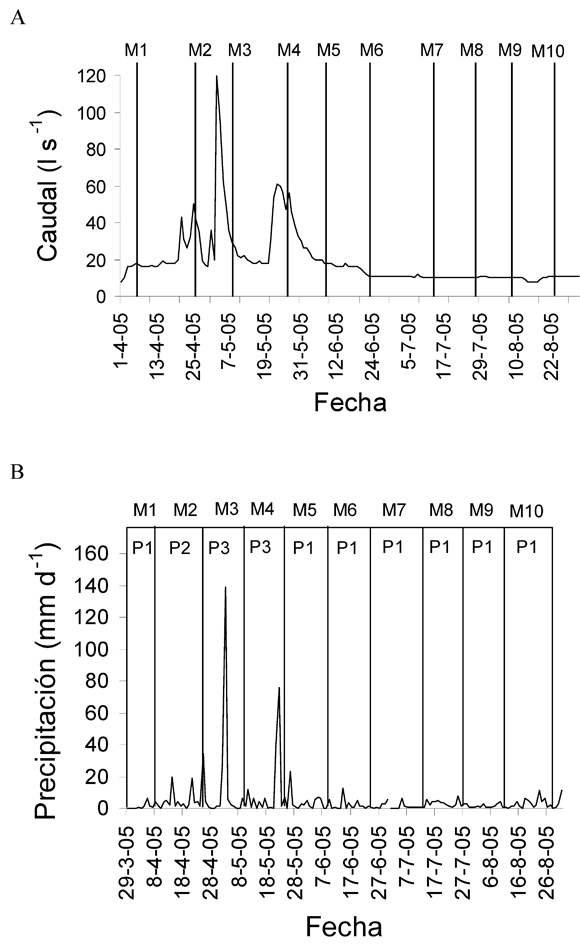
Precipitación y caudales
Aunque los datos de precipitación y caudal obtenidos de la Empresa de Acueducto y Alcantarillado de Bogotá, se registraron con una resolución diaria, se hará referencia a los valores de la intensidad de la precipitación y del caudal promedio para los 10 días anteriores a las fechas de muestreo, debido a que en cuencas pequeñas como la Quebrada La Vieja, la cual presenta pocos eventos de máxima precipitación, los incrementos del caudal obedecen más a la saturación del suelo de la cuenca por efecto de precipitaciones acumuladas en días anteriores, que a un solo evento de precipitación (Larrota, com. pers).
Cuantificación de MOP
En el presente estudio se consideró como MOPG aquella MOP con diámetro mayor a 1000 µm y MOPF como aquella MOP con diámetro entre 1 µm y 1000 µm de acuerdo a los procedimientos desarrollados por Vannote et al. (1980).
En el laboratorio, la MOPG fue cuantificada mediante el método de calcinación en una mufla a 550 °C, de acuerdo a los procedimientos sugeridos por APHA (1985). La MOPF fue filtrada con la ayuda de una bomba de vacío y retenida en un filtro de microfibra de vidrio (Whatman GF/C). La MOPF retenida en el filtro fue secada a 110 °C y luego calcinada a 550 °C, de acuerdo a los procedimientos sugeridos por Goodman (1964), Gratteau & Dick (1968) y APHA (1985).
Toma de muestras
Entradas de MOPF lateral- Se utilizaron 8 bandejas (réplicas) con una superficie de captura de 10 cm, cubiertas con papel aluminio que impidiera la entrada vertical de materiales y a una altura superior al nivel máximo de incremento de la columna de agua. De forma aleatoria se dispusieron cuatro bandejas en cada orilla de la Quebrada.
Entradas de MOPG lateral- se ubicaron ocho trampas que presentaron un ojo de malla de 1 mm y una longitud de captura de 30 cm (Figura 2B). Se ubicaron aleatoriamente siete trampas en el límite superior al nivel máximo de incremento de la columna de agua en la ladera de la Quebrada, teniendo en cuenta la heterogeneidad de la misma.
Entradas de MOPG vertical- se ubicaron 6 trampas de hojarasca (Área: 1 m2) en puntos aleatorios a lo largo del tramo de 200 m. Estas trampas presentaron marco fijo de tubo de 1/2`` para impedir su movimiento por acción del viento.
Transporte de MOPG- en el tramo de estudio se desarrollaron 10 campañas de muestreo con una duración de 24 h para cada una. Se sumergieron parcialmente cuatro trampas de deriva (ojo de malla de 250 µm) durante un período de 24 h, a intervalos de 12 h (mañana-noche), para evitar que se colmataran. En el laboratorio se separó la fracción superior a 1000 µm por tamizado de la muestra.
D = (g)/(T.V.A)
Donde, D = Densidad de deriva de MOPG (g PSLC de MOPG x m-3); g = Gramos de PSLC de MOG; T = Tiempo de exposición de la red (segundos); V = Velocidad de la corriente en la boca de la red (m/s); A = Área sumergida de la red (m2)
Transporte de MOPF- En el tramo de estudio se tomaron 3 muestras de 500 ml de agua (cada una con tres réplicas) a intervalos de 8 horas en un período de 24 h por muestreo. Posteriormente en el laboratorio se realizó un filtrado al vacío de 200 ml por muestra. La MOPF retenida en el filtro fue secada a 110 °C y luego calcinada a 550 °C, de acuerdo a los procedimientos sugeridos por Goodman (1964), Gratteau & Dick (1968) y APHA (1985) en filtros de fibra de vidrio (Whatman GF/C, tamaño de poro: 1 µm). El transporte MOPF se calculó con base en los procedimientos de Hauer & Lamberti (1996) de acuerdo a la siguiente fórmula:
D = (g)/(C.T.Al)
Donde, D = Densidad de deriva de MOPF (g PSLC de MOPF x m-3); g = Gramos de PSLC de MOPF; C = Caudal de la sección en el tramo de muestreo (m3.s-1); T = Tiempo de exposición del recipiente (se asume un segundo); Al = Alícuota filtrada de 0.002 m-3.
Análisis estadístico
Previo al desarrollo de las pruebas estadísticas, se probaron los supuestos de Aleatoriedad de residuos (prueba de corridas arriba y debajo de la mediana) homogeneidad de varianza (prueba de Bartlett) y normalidad (bondad de ajuste de Kolmogorov-Smirnov: K-S), mostrando un comportamiento aleatorio de las variables, pero los supuestos de normalidad y homogeneidad resultaron no ser cumplidos en la gran mayoría de los casos, por lo que todo el análisis estadístico fue hecho con base en pruebas no paramétricas. La prueba Kruskal-Wallis (K-W), fue utilizada para buscar diferencias entre las variables: (1) entradas de MOP alóctona con los eventos de precipitación (P1, P2, P3) y (2) transporte de MOPG y MOPF con relación a los eventos de alto y bajo caudal (C1, C2, C3), mientras que para los análisis de asociación entre las variables y los parámetros ambientales (precipitación y caudal) se empleó la prueba de rangos de Spearman (rs). El paquete estadístico utilizado fue el Statgraphics versión 4.0.
RESULTADOS
Patrones ambientales
Los datos arrojados por el limnígrafo ubicado en la Quebrada La Vieja, mostraron que los valores de precipitación para el primer muestreo (M1) correspondieron a un período de baja precipitación (P1), ya que la precipitación durante este muestreo no superó los 6 mm. M2 representó un período de aguas medias (P2), con una precipitación promedio baja (8.4 mm d-1), sin embargo un día antes de culminar el muestreo se registró una precipitación de 34.1 mm. M3 y M4 correspondieron al período de aguas altas (P3). Durante M-3 se presentó mayor registró de precipitación (139 mm d-1) simultáneo al mayor registro de caudal en los últimos 11 años (120 l.s-1). M4 igualmente corresponde a un período de lluvias altas, donde se presentó un valor máximo de precipitación de 76 mm d-1, el cual supera el mayor registro reportado para el año 2004 (Gutiérrez 2006). Finalmente durante M5 hasta M10 los valores de precipitación no superaron los 12.5 mm d-1, lo cual corresponde a un período P1 (Figura 1A).
Los valores de precipitación previos al muestreo 1 (M1) se manifestaron en un caudal promedio bajo (C1) de 12.5 l.s-1 (Figura 1). M-2 representó el período de caudal medio (C2) con un valor promedio de 27 l.s-1. M3 y M4 correspondieron al período de alta descarga (C3). Durante M3 se presentó el mayor registro de precipitación (139 mm d-1), simultáneo al mayor registro de caudal en los últimos 11 años (120 l.s-1). M4 presentó un valor máximo de precipitación diaria (76 mm.d-1) y una descarga promedio de 38 l.s-1, superando al mayor registro de precipitación y caudal reportado para el año 2004 (Gutiérrez, 2006). Finalmente de M5 a M10 los valores de precipitación no superaron los 12.5 mm d-1, con un caudal promedio de 12.45 l.s-1, correspondiendo a un período de aguas bajas (C1) (Figura 1 B).
Los muestreos desarrollados durante M3 y M4 mostraron un valor de precipitación promedio significativamente mayor en comparación con el período seco correspondiente a M1 y M5 hasta M10 (K-W=79.58, p<0.01, n=114), aunque en términos generales se presentó un régimen de precipitación constante con bajos valores Se presentó una diferencia significativa entre el período de alto caudal correspondiente a M3 y M4 (valor promedio de 21.45 l.s-1) y el período de bajo caudal (M1, M5-M6), que mostró un valor promedio de 4.93 l.s-1 (K-W=82.18, p<0.01, n=120), aunque en términos generales el régimen de descarga fue constante y de bajos valores.
Entradas de MOP
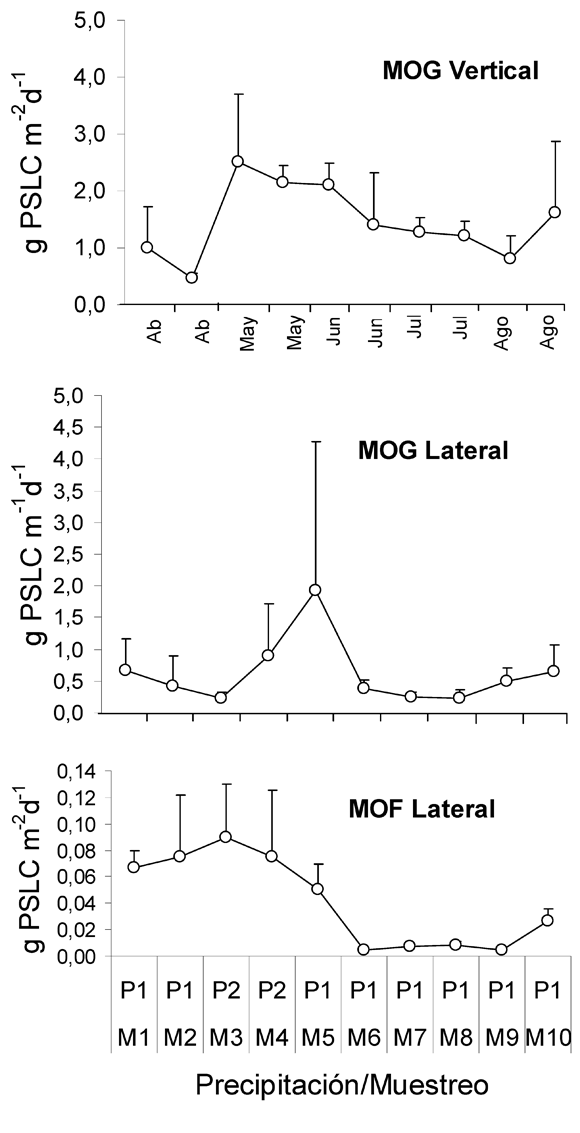
Las cantidades de MOPG vertical, MOPG lateral y MOPF lateral correspondiente a los 5 meses de muestro (155 días) fueron 450.8 g PSLC m-2, 191 g PSLC m-1 y 12.65 g PSLC m-1 respectivamente, con valores promedio de entradas diarias durante de 3.12 g PSLC m-2, 1.23 g PSLC m-1 y 0.08 g PSLC m-1 respectivamente.
La entrada de MOPG vertical para el período de sequía fue de 1.55 (±0.52) g PSLC m-2 d-1, 0.46 (±0.08) g PSLC m-2 d-1 para aguas medias y para el período de lluvias fue de 1.85 (±0.29) g PSLC m-2 d-1. La cantidad promedio de MOPG lateral para el período de sequía fue de 0.66 (±0.55) g PSLC m-1 d-1, 0.42 (±0.48) g m-1 d-1 durante aguas medias y para el período de lluvias fue de 0.57 (±0.45) g PSLC m-1 d-1. Finalmente los valores encontrados para la MOPF lateral en el período de sequía fue de 0.02 (±0.01) g PSLC m-1 d-1, 0.075 (±0.047) g PSLC m-1 d-1 en aguas medias y para el período de lluvias fue de 0.08 (±0.05) g PSLC m-1 d-1. Los resultados muestran que, en promedio, la entrada lateral de MOPG fue aproximadamente 30 veces mayor a las entradas de MOPF (Figura 2).
Entradas de MOPG vertical- mostraron su mayor registro en M3 con un valor de 2.50 g PSLC m-2 d-1 y una diferencia altamente significativa en estos aportes con respecto al resto de muestreos que correspondieron a menores intensidades de precipitación (K-W=26.26, p=0.002, n=53). En términos descriptivos, la Figura 3 muestra que el pico máximo de entrada de esta materia se presentó en M-3, correspondiente al período de mayor intensidad de la precipitación, aunque la asociación entre éstas es muy baja y poco significativa (rs= 0.19, p=0.16, n=53). Para el período de Agosto se muestra una tendencia al incremento en la MOPG vertical como sucede con las otras entradas de MOP (Figura 2).
Entradas MOPG lateral- presentaron su mayor valor en M5 (1.93 g PSLC m-1 d-1) mostrando diferencias significativas con los otros muestreos y con la intensidad de la precipitación (K-W=23.82, p=0.004, n=66). Se presenta una asincronía entre los incrementos de estos aportes alóctonos de MOPG y la intensidad de la precipitación (Tabla 1), lo cual es sustentado por una correlación poco significativa e inversa (rs= -0.06, p=0.58, n=66).
Entradas de MOPF lateral- presentaron su mayor valor en M3 (0.09 g PSLC m-1 d-1) mostrando diferencias significativas con el resto de muestreos y con la intensidad de la precipitación (K-W=23.3, p=0.005, n=32). A diferencia de las entradas laterales de MOPG, se demuestra una tendencia al incremento los las entradas de MOPF lateral con la intensidad de la precipitación aunque se presenta una baja correlación pero significativa (rs= 0.47, p=0.04, n=32).
En concordancia con los patrones mencionados anteriormente, se presenta una asociación baja entre la MOPG vertical y la MOPF lateral, pero significativa (rs= 0.47, p=0.008, n=32). Además se muestra una correlación inversa y poco significativa entre la MOPG lateral y MOPF lateral (rs= -0.16, p=0.36, n=32) y entre la MOPG vertical y la MOPG lateral (rs= -0.12, p=0.21, n=32).
Con base en los resultados anteriores, se deduce que existen diferencias en las cantidades de los aportes de MOP para eventos de lluvia y de sequía. De igual forma se presenta efecto directo de la precipitación sobre la MOPF lateral, lo cual no es visible para la MOPG lateral. Además se presentan umbrales de precipitación que se traducen en incrementos significativos de MOPG vertical, los cuales permanecen meses después de terminado el período de lluvias.
Transporte de MOP
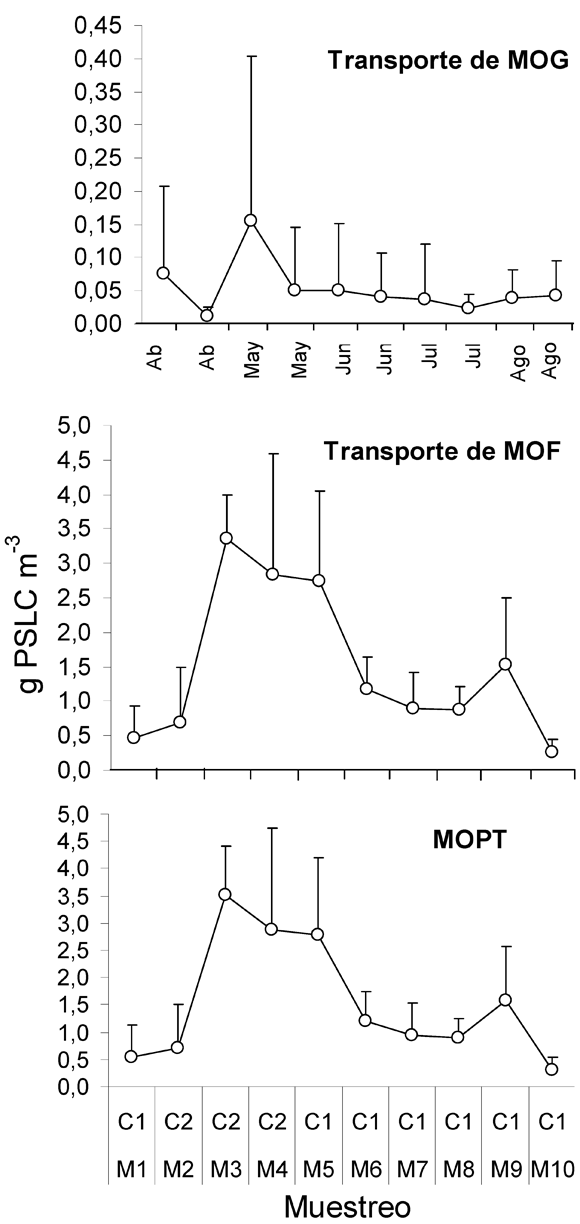
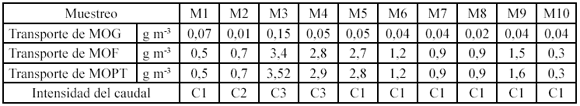
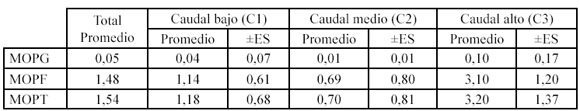
El transporte promedio de MOPG, MOPF y MOP total (MOPT= MOPG+MOPF) en el tramo de estudio fue de 0.05 g m-3, 1.48 g m-3 y 1.54.6 m-3 respectivamente. Los valores de pérdidas por transporte de MOPG para el período de bajo caudal fue de 0.04 (±0.07) g m-3, 0.01 (±0.01) g m-3 para aguas medias y para el período alto caudal fue 0.1 (±0.17) g m-3. La cantidad promedio de MOPF transportada aguas abajo para el período de sequía fue de 1.14 (±0.61) g m-3, 0.69 (±0.8) g m-3 durante aguas medias y para el período de lluvias fue de 3.1 (±1.2) g m-3. Finalmente los valores encontrados para la MOPT, correspondientes a la sumatoria de la MOPG y la MOPF transportada durante el período de sequía fue de 1.18 (±0.68) g m-3, 0.7 (±0.81) g m-3 en aguas medias y para el período de lluvias fue de 3.2 (±1.37) g m-3. Cabe resaltar que la cantidad de MOPF transportada por la Quebrada fue treinta veces superior (aproximadamente) al transporte de MOPG para períodos de alto y bajo caudal (Tabla 1).
Transporte de MOPG- La MOPG transportada presentó el mayor registro con un valor promedio de 0.154 g m-3, simultáneo al período de mayor intensidad del caudal promedio para los diez días previos a M3; aunque este incremento se debe a un valor atípico del caudal de gran magnitud (Figura 1). No se presentó una variación en el transporte de MOPG en relación con el resto de muestreos representados por la baja intensidad del caudal (K-W= 1.51, p=0.46, n=77). La asociación entre esta variable y la magnitud del caudal fue muy baja y poco significativa (rs= 0.08, p=0.46, n=77). Cabe resaltar que la menor cantidad de MOPG transportada se presentó durante el período de caudal medio, lo cual puede afectar la significancia del modelo de correlación (Figura 3).
Transporte de MOPF- Se presentó un comportamiento similar al mostrado por la MOPG, ya que para M3 se presenta un valor máximo de 3.36 g m-3, correspondiente al período de alto caudal, aunque su incremento fue más prolongado que lo mostrado por la MOPG, ya que permaneció hasta M5 (Figura 3). Además, se presentaron diferencias en estos aportes de MOPF con respecto al resto de muestreos correspondientes a bajos caudales (K-W= 8.98, p=0.011, n=42). La asociación entre esta variable y la magnitud del caudal fue muy baja (rs= 0.31, p=0.04, n=42), debido a los datos atípicos que presentó esta variable durante M5 y M9 (Figura 3).
Transporte de MOPT- Las pérdidas totales de MOP fueron expresadas en términos de la MOPT transportada aguas abajo (MOPG+MOPF), (Figura 2). Se presentó una alta asociación entre la MOPT y el transporte de MOPF (rs= 0.99, p<<0.001, n=42) debido a que la MOPF aportó casi la totalidad de la MOP en deriva (MOPT). Se presentaron diferencias entre la MOPT y la magnitud del caudal (K-W= 9.28, p=0.01, n=42) aunque no se presentó una fuerte relación funcional entre estas variables (rs= 0.32, p<<0.04, n=42), igual a lo sucedido con la MOPF.
DISCUSIÓN
Régimen hidroclimático y estructura física
Se presentaron pocos eventos de variación de la precipitación y un régimen de caudal con valores bajos y estables, dos eventos puntuales de incremento extremo en la descarga durante el mes de Mayo correspondiente a M3 y M4 y un incremento de magnitud intermedia a finales de Abril correspondiente a M2 (Figura 1B). Se pudo apreciar que estos sucesos se producen como respuesta a los incrementos en la intensidad de la precipitación (Figura 1A), con magnitudes que presentan períodos de retorno de aproximadamente 10 años. Las condiciones climáticas y la respuesta en las entradas y el transporte de MOP en la Quebrada La Vieja, son el reflejo de un sistema bien conservado que presenta una buena cobertura vegetal, poca saturación de los suelos debido a su textura arenosa y abundantes tapetes de musgos en sus laderas (Vargas 2003). Esta cualidad de los suelos pudo ser reflejada por los bajos valores en la escorrentía de la MOPF lateral que ingresó a la Quebrada, en comparación con las entradas de MOPG lateral, la cual fue almacenada en el suelo ripario para su posterior ingreso.
Entradas de MOP alóctona
En términos generales, se presentaron picos de incremento en los aportes de MOP a la Quebrada en el mes de Mayo, correspondiente a M-3 y M-4 (tabla1), generados por umbrales de precipitación. Esto guarda relación con estudios desarrollados en regiones templadas por Bärlocher (1983), Abellho & Graça (1998) y Larned (2000) en relación con el aporte de hojarasca a Quebradas de primer orden de Suiza, España, Portugal y Hawai, respectivamente. Trabajos desarrollados en el trópico muestran un patrón similar (McClain et al. 1996, Taner 1980, Taner et al. 1998, Marrs et al. 1988). Se destacan los trabajos desarrollados en la Ciudad de Bogotá por León & Quiroga (1982), Arenas (1988) y Hernández & Murcia (1994) quienes también resaltan la asociación existente en la producción de hojarasca en términos de peso seco con la precipitación de la región, en correspondencia con Vargas et al. (2003) para la Quebrada La Vieja.
La variación temporal en los aportes de MOPG alóctona se ha descrito a escala de paisaje (Naiman, 1982, Peterson et al. 1989, Richardson 1992, Wallace et al. 1995) debido a la variabilidad de los parches o mosaicos que forman la vegetación riparia, la fenología de la vegetación y el efecto de las lluvias y la sequía (Larned 2000). En el presente estudio se pudo encontrar que el aporte de materiales fue más o menos constante durante los cinco meses de muestreo, aunque la MOPG vertical se incrementó 2 veces en período de lluvias, como sucedió con la MOPF lateral, que se incrementó 4 veces en magnitud con relación a este evento climático.
La MOPG vertical presentó una respuesta directa en relación con la precipitación, a pesar de que su descenso se dio meses después de haber comenzado el período seco. Un patrón similar es reportado en los trabajos de León & Quiroga (1982), Arenas (1988) y Hernández & Murcia (1994). Webster & Benfield (1986) reportan incrementos en la MOPF alóctona en relación significativa con eventos hidroclimáticos extremos como las tormentas sobre Quebradas de Norteamérica. Trabajos sobre la interacción bosque-río en las cabeceras de las montañas, han demostrado por la relación directa entre la densidad del bosque ripario y el efecto hidroclimático, las entradas de material alóctono a las Quebradas (Zah & Uehlinger 2001).
Este estudio puede ser visto como una herramienta complementaria para la interpretación de otros procesos ligados a la dinámica de MOP, como la tasa de su producción por parte de comunidades autotróficas (producción autóctona de MO) y el aprovechamiento energético de esta materia por parte de comunidades heterotróficas (procesos de respiración), como lo describen Benke et al. (1988) y Abellho & Graça (1996) en sus trabajos con las comunidades presentes en las biopelículas, en los cuales encontraron una fuerte relación entre la tasa de aportes de MOP alóctona y la biomasa de estas comunidades, a pesar de que la penetración lumínica fue muy escasa. Factores estos que son similares en la quebrada La Vieja, en donde se presenta un aporte relevante en la producción alóctona por el bosque ripario, en especial de la fracción gruesa. Webster et al. (1999) registran que las entradas de hojas de los árboles riparios son, por lo general, la mayor fuente de energía para los organismos heterotróficos.
Un aspecto que no pudo tenerse en cuenta fue el aporte de palizadas por caídas de árboles hacia el cauce de la quebrada, suceso que pudo observarse durante el período de muestreo, como resultado de los eventos puntuales de fuertes precipitaciones registrados durante los muestreos M-3 y M-4. Richardson (1992) y Larned (2000) hacen especial énfasis en la dificultad para hacer cuantificaciones de esta categoría de MOP debido a los limitantes que muestran los métodos tradicionales para la cuantificación de la cosecha de MOPG. Campbell et al. (1992) encontraron que la biomasa de palizadas que ingresa a las quebradas y es transportada, puede ser mayor a la que cae al bosque ripario y que por lo general es subestimada debido a la dificultad de su muestreo.
En este sentido Abellho & Graça (1996) mencionan que la forma más cercana a la realidad para estimar las entradas de MOPG a las quebradas, es por medio de la cuantificación de la biomasa en forma de palizadas que ingresa en un momento dado al cauce. Webster et al. (1999) mencionan que la dificultad para evaluar el aporte de MOPG por la entrada de palizadas se presenta debido a la distribución parchada de su aporte en espacio y tiempo. Aunque el aporte de MOPG de este componente puede llegar a los 880 g m-2 año-1 en bosques adultos y extensos de coníferas, la tasa de su fragmentación y/o asimilación por comunidades heterótrofas no excede a la de hojarasca. Esto se demuestra debido a que el área superficial expuesta para esta intervención biológica es demasiado pequeña en relación con su volumen y por lo tanto, el aporte al flujo energético en sistemas riparios de montaña es muy pobre. El aporte de madera de estas palizadas tiene un valor agregado en cuanto a su importancia en la estructura física de quebradas de cabecera que se encuentran en íntima interacción bosque-río, además de aportar un substrato importante para el asentamiento de las biopelículas y sus procesos sucesionales (Tank et al. 1993).
Si se proyectan los valores correspondientes a las entradas de MOPG vertical calculados durante los 5 meses a un período anual se obtiene un valor aproximado a los 1081 g m-2 año-1 lo cual corresponde a un valor alto en comparación con otros trabajos. Larned (2000) para Norte América reporta mínimos de 300 g m-2 año-1 y máximos de 1050; Zah & Uehlinger (2001) encontraron valores máximos de 23 g m-2 año-1 en ríos Suizos; Cillero et al. (1999) reportan valores de 730 g m-2 año-1 en Quebradas ibéricas de segundo orden; Benfield et al. (2000) reportan para Quebradas de orden menor en las montañas de los Apalaches un aporte de MOPG cercano a 460 g m-2 año-1.
Estudios en regiones tropicales muestran un patrón diferente al encontrado en este estudio, como sucede con Quebradas de montaña ubicadas en la región amazónica del Perú (McClain et al. 1996) donde se ha reportado que estos bosques montañosos son menos productivos que los de tierras bajas (Tanner 1980), debido a los declives de altitud, con temperaturas y radiación que después afectan el clima, la composición de los suelos, el pH (Marrs et al. 1988, Tanner et al. 1998), la composición de las especies, las tasas de fotosíntesis y la poca caída de hojarasca (Vitousek & Stanford, 1986). Benfield et al., (2000) reportan un patrón similar, quienes demostraron la influencia negativa del gradiente elevacional en la producción de hojarasca en las montañas de los Apalaches.
Para Colombia se destacan los trabajos Vargas (2003) en la Quebrada La Vieja, que encontraron valores máximos de MOPG por la caída de hojarasca de 1300 g m-2 año-1 en relación con el período de lluvias, con Clusia multiflora, Vaccnum sp. y Weinmania tomentosa como las especies dominantes en el aporte de MOPG al mantillo del suelo ripario. León & Quiroga (1982) reportan cantidades de 920 y 475 g m-2 año-1 en Robledales y Bosque Altoandino de la Sabana de Bogotá. Estos autores destacan la asociación existente en la producción de hojarasca con la precipitación de la región, al igual que Arenas (1988) y Hernández & Murcia (1994).
La calidad de esta MOPG alóctona juega un papel relevante en términos de energía para comunidades acuáticas, como sucede en épocas de primavera en regiones holárticas, donde existe una mayor proporción de flores que caen al lecho de las Quebradas, las cuales ofrecen un mayor aporte energético a comunidades de insectos en comparación con las hojas (Fittkau 1964, Winterbourn, 1976 En: Abellho & Graça 1998) y palizadas (Webster et al. 1999).
Transporte de MOP
De acuerdo a los resultados obtenidos por las diez campañas, se presentaron diferencias estadísticas en la cantidad de MOPT transportada en el tramo a escala temporal. La MOPG aumentó 1.5 veces durante el período de alto caudal, así como la MOPF aumentó 1.3 veces en esta fase (Tabla 3), aunque esta cantidad no fue suficiente para mostrar una correspondencia estadísticamente significativa con la intensidad del caudal debido a la estabilidad que presentó el caudal durante las campañas de muestreo. Esto demuestra que el caudal no presentó un efecto significativo en el flujo de MOP hacia aguas abajo para una Quebrada conservada como la Vieja, a pesar de que el período de muestreo correspondió a un año lluvioso.

Cabe resaltar que las campañas de muestreo desarrolladas durante la época de lluvias (M3 y M4), fueron realizadas cinco días después de haber iniciado los eventos máximos de incremento en caudal (Figura 1B). Durante este lapso de tiempo, pudo presentarse un barrido masivo de MOP similar a lo sucedido con las comunidades de macroinvertebrados bentónicos (ver Capítulo V). En tal sentido, sería de esperar que durante el primer día de haber iniciado estos eventos puntuales de incremento en la descarga, se generara un umbral de escorrentía suficiente para movilizar una gran cantidad de MOP, caso en el cual se podría presentar una relación altamente positiva entre la descarga y el transporte de MOPT. Cuffney & Wallace (1989) encontraron en quebradas templadas de los Apalaches del Sur en Norte América que el exporte de MOPG estuvo relacionado con patrones de descarga, particularmente en épocas de creciente posteriores a períodos de sequía. Larned (2000) reporta para quebradas templadas que un período de siete días es apropiado para estimar el transporte representativo de materiales durante eventos de creciente (postsequía); posterior a este tiempo es muy poca la materia disponible para ser movilizada.
Sería relevante conocer la dinámica de estos ecosistemas y su respuesta ante eventos periódicos en el régimen de descarga para saber, a ciencia cierta, si existe un umbral de escorrentía capaz de ocasionar una respuesta significativa en el incremento del transporte de MOP y cuál sería la velocidad de recuperación de éstas en cuanto a los aportes y almacenamiento de materiales al lecho de la quebrada.
O´Hop & Wallace (1983) analizaron el efecto de las tormentas sobre el transporte de detritos fino y grueso y encontraron un incremento significativo en la movilización de estas fracciones de MOP posterior al incremento en el flujo como consecuencia de las tormentas. Bilby & Likens (1979) y Gurtz et al. (1980) reportan que el incremento en el transporte de seston y MOPF se presenta previo a los picos en la descarga.
El transporte de MOPT a escala de tramo no presentó una correspondencia con el patrón de descarga. Es probable que a escalas de mayor tamaño, se presente un comportamiento diferente, como lo reportan Shannon et al. (1996), quienes desarrollaron investigaciones en el río Colorado y encontraron que a escala de cuenca sí se presenta una correlación positiva entre la deriva de MOPG (detritos) con la descarga y la distancia de movilización.
En la Quebrada La Vieja, el tiempo de duración en los picos de incremento y descenso en el transporte MOPT puede ser afectado por otros factores adicionales a la presencia de crecientes, como lo son la geomorfología, pendientes fuertes y el encañonamiento que esta Quebrada exhibe. Minshall et al. (2000) destacan la importancia de factores como la infiltración hiporreica y la retención biótica, los cuales demuestran un efecto en la tasa de transporte de MOPF en adición a los fenómenos hidrodinámicos y gravitacionales.
Según Vannote et al. (1980), tramos de cabecera presentan una relación MOG/MOPF >1. Es muy probable que grandes cantidades de MOPG sean retenidas por el lecho de la Quebrada, a diferencia de la MOPF, la cual tiende a ser movilizada aguas abajo en mayores cantidades, muy superiores a la MOPG, como sucedió en el tramo de la Quebrada La Vieja.
Relación MOPG/MOPF transportada
La MOPF transportada fue 30 y 26 veces superior a la MOPG durante períodos de alto y bajo caudal respectivamente (Tabla 2).
Larned (2000) menciona que la MOPG que ingresa a las quebradas es fragmentada a partículas finas y disueltas durante períodos de sequía, incrementando la disponibilidad de MOPF durante períodos de lluvia, lo cual es reflejado por la mayor densidad en comunidades de insectos trozadores residentes y esporádicos. Allan (1995) y Wallace & Grubaugh (1996) demostraron que la MOPG es movilizada episódicamente por el efecto de tormentas, alternando etapas de almacenamiento y erosión. En consecuencia, sólo una pequeña fracción del ciclo anual de descarga pudo mostrar un transporte masivo de la MOPG. En cambio la MOPF fue movilizada continuamente río abajo en conjunto con las moléculas de agua y su almacenamiento pudo ocurrir en el sedimento del lecho o en la zona hiporreica.
La alta heterogeneridad que presentó el tramo de la Quebrada La Vieja (debido a una gran cantidad de obstáculos como parches de palizadas, sustratos emergentes, presencia de rápidos y remansos) pudo haber influenciado para la poca movilidad de la MOPG. Webster et al. (1999) mencionan que estos factores son relevantes en la dinámica de las entradas, retención y pérdidas por transporte de MOPG en quebradas.
El balance de entradas y salida de MOPG/MOPF en la Quebrada La Vieja, demuestra un dominio en las entradas y retención de MOPG (ver Capítulos II y IV) y la MOPF dominó en las salidas o pérdidas por transporte aguas abajo. Este balance de materiales se destaca si se quieren desarrollar aproximaciones en cuanto al flujo de MOP en la interacción bosque-río (Allan 1995), estudios aplicativos en restauración de quebradas de cabecera, efecto hidroclimático en la dinámica de la MOP y comparaciones con teorías desarrolladas en sistemas lóticos (Vannte et al. 1980).
AGRADECIMIENTOS
Los autores agradecen al Laboratorio de Invertebrados Acuáticos de la Universidad Nacional de Colombia y a todos sus miembros, por su apoyo en el desarrollo exitoso del presente trabajo tanto en la fase de campo como en la de laboratorio.
LITERATURA CITADA
1. ALLAN, J.D. 1995. Stream Ecology, estructure and funtion of running waters. Edit. Chatman & Hall. [ Links ]
2. AMERICAN PUBLIC HEALTH ASSOCIATION (APHA). 1985. Standard Methods for the Examination of Water and Waste Water, 16th ed. American Public Health Association, Washington, DC. [ Links ]
3. ANDERSON, N.H. & J.R. SEDELL. 1979. Detritus processing by macroinvertebrates in stream ecosystems. Ann. Rev. Entomol. 24:351-377. [ Links ]
4. ARENAS, H. 1988. Producción y descomposición de hojarasca en un bosque nativo y un bosque de Eucaliptus en la región de Monserrate. Tesis de Grado. Biología. Universidad Nacional de Colombia, Bogotá. [ Links ]
5. BÄRLOCHER, F. 1983. Seasonal variation of standing crop and digestibility of CPOM in a Swiss Jura stream. Ecology 64:1266–1272. [ Links ]
6. BENFIELD, E.F., J.R. WEBSTER, J.R. HUTCHENS, J.L. TANK & P.A. TURNER. 2000. Organic matter dynamics along a stream-order and elevational gradient in a southern Appalachian stream. Verh. Internat. Verein. Limnol. 27:1-5. [ Links ]
7. BENKE, A.C., C.A. HALL, C.P. HAWKINS, R.H. LOWE-MCCONNEL, J.A. STANFORD, K. SUBBERKROPP & J.V. WARD. 1988. Bioenergetic considerations in the analysis of stream ecosystems. J. N. Am. Benthol. Soc. 7:480–502. [ Links ]
8. BILBY, R.E. & P.A. BISSON. 1992. Relative Contribution of Allochthonous and Autochthonous Organic Matter to the Trophic Support of Fish Populations in Clear-Cut and Old-Growth Forested Headwater Streams. Canadian Journal of Fisheries and Aquatic Sciences 49:540-551. [ Links ]
9. CAMPBELL, I.C., K.R. JAMES, B.T. HART, & A. DEVEREAUX. 1992. Allochthonous coarse particulate organic material in forest and pasture reaches of two southeastern Australian streams. II. Litter processing. Freshwater Biology 27:353-365. [ Links ]
10. CORTÉS, L.A. 1976. Los Suelos de la Sabana de Bogotá y sus Alrededores Subdirección Agrológica Instituto Geográfico Agustín Codazzi. Bogotá. [ Links ]
11. CUFFNEY, T.F. & J.B. WALLACE. 1989. Discharge export relationships in headwater streams: the influence of invertebrate manipulations and drought. Journal of North American Benthological Sosiety 8:331-341. [ Links ]
12. CUMMINS, K.W. 1974. Structure and funtion of strem ecosystem. Ecology. 61:1107-1113. [ Links ]
13. CUMMINS, K.W. & M.J. KLUG. 1979. Feeding ecology of stream invertebrates. Annual Review of Ecology and Systematics 10:147-172. [ Links ]
14. CUMMINS, K.W., J.R. SEDELL, F.J. SWANSON, G.W. MINSHALL, S.G. FISHER, C.E. CUSHING, R.C. PETERSEN, & R.L. VANNOTE. 1983. Organic matter budgets for stream ecoystems, p. 299-353. En: J.R. Branes & G.W. Minshall (Editores). Stream ecology: Application and testing of general ecological theory. Plennum, Nueva York. [ Links ]
15. CUMMINS, K.W., M.A. WILZBACH, D.M. GATES, J.B. PERRY & W.B. TALIAFERRO. 1989. Shredders and riparian vegetation. Bioscience 39:24-30. [ Links ]
16. CUSHING, C.E., G.W. MINSHALL & J.D. NEWBOLD. 1993. Transport dynamics of fine particulate organic matter in two Idaho streams. Limnology and Oceanography 38:1101-1115. [ Links ]
17. EHRMAN, T.P. & G.A. LAMBERTI. 1992. Hydraulic and particulate matter retention in a 3rd-order Indiana stream. Journal of North American Benthological Society 11:341-349. [ Links ]
18. ELLIOT, J.M. 1971. The distances traveled by drifting invertebrates in a lake district stream. Oecologia 6:350-379. [ Links ]
19. ESPINAL, L.S. & E. MONTENEGRO. 1963. Formaciones vegetales de Colombia. Instituto Geográfico Agustín Codazzi. Bogotá. [ Links ]
20. FISHER, S.G. & G.E. LIKENS. 1973. Energy Flow in Bear Brook, New Hampshire: An Integrative Approach to Stream Ecosystem Metabolism. Ecological Monographs 43:421-439. [ Links ]
21. FRISSELL, C.A., W.J. LISS, C.E. WARREN & M.D. HURLEY. 1986. A hierarchical framework for stream classification: viewing streams in a watershed context. Env. Manage. 10:199-214. [ Links ]
22. GOODMAN, B.L. 1964. Processing thickened sludge whit chemical conditioners. En: Sludge Concentration, Filtration and Incineration. Univ. Michigan Continued Education Ser. No. 113, Ann. Arbor. [ Links ]
23. GRATTEAU, J.C. & R.I. DICK. 1968. Activated sludge suspended solids determinations. Water Sewage Works 115:468. [ Links ]
24. GUTIÉRREZ, J.D. 2006. Caracterización del metabolismo y de la oferta de recursos de materia orgánica para la fauna de macroinvertebrados bentónicos en una Quebrada de montaña de orden menor. Tesis de Doctorado. Universidad Nacional de Colombia. Bogotá, D.C. [ Links ]
25. HAUER, F.R. & G.A. LAMBERTI. 1996. Methods in stream ecology. Academy Press. 674 p. [ Links ]
26. HERNÁNDEZ, M.L. & M.A. MURCIA. 1994. Estimación de la productividad primaria de Espeletia grandiflora H & B y Pinus patula Schl & Cham en el páramo “el Granizo”, Cundinamarca Colombia. Tesis de Grado. Biología. Universidad Nacional de Colombia, Bogotá, D.C. [ Links ]
27. INSTITUTO GEOGRÁFICO AGUSTÍN CODAZZI (IGAC). 1968. Estudio General de Clasificación de los suelos de la Cuenca Alta del Río Bogotá para fines Agrícolas. Vol IV No. 1. Departamento Agrológico. Instituto Geográfico Agustín Codazzi. Bogotá. [ Links ]
28. KARLSSON, O.M., J.S. RICHARDSON, & P.M. KIFFNEY. 2005. Modelling Organic Matter Dynamics in Headwater Streams of South-Western British Columbia, Canada. Ecological Modelling 183:463-476. [ Links ]
29. LARNED, S.T. 2000. Dynamics of coarse riparian detritus in a hawaiian stream ecosystem: a comparison of drought and post-drought conditions. J. N. Am. Benthol. Soc. 19(2):215–234. [ Links ]
30. LEÓN, M.R. & R.A. QUIROGA. 1982. Producción de materia orgánica en dos bosques circundantes a la Sabana de Bogotá. Tesis de Grado. Biología. Universidad Nacional de Colombia, Bogotá. [ Links ]
31. MARRS, R.J., J. PROCTOR, A. HEANEY & M.D. MOUNTFORD. 1988. Changes in soil nitrogen - mineralization and nitrification along an altitudinal transect in tropical rainforest in Costa Rica. Journal of Ecology 76:466-482. [ Links ]
32. MCCLAIN, M.E., J.E. RICHEY & R.L. VICTORIA. 1996. Andean contributions to the biogeochemistry of the Amazon River system. Bulletin de l'Institut Fracais d'Etudes Andines. 24:425-437. [ Links ]
33. MINSHALL, G.W., CH.T. ROBINSON, & D.E. LAWRENCE. 1985. Postfire responses of lotic ecosystems in Yellowstone National Park, U.S.A. Can. J. Fish. Aquat. Sci. 54: 2509–2525. [ Links ]
34. MINSHALL, G.W., J.T. BROCK & T.W. LAPOINT. 1982. Characterization and dynamics of benthic organic matter and invertebrate functional feeding group relationships in the Upper Salmon River, Idaho (U.S.A.). Int. Rev. ges. Hydrobiol. 67: 793–820. [ Links ]
35. MINSHALL, G.W., S.A. THOMAS, J.D. NEWBOLD, M.T. MONAGHAN & C.E. CUSHING. 2000. Physical factors influencing fine organic particle transport and deposition in streams. Journal of North American Benthological Sosiety 19(1):1-16. [ Links ]
36. NAIMAN, R.J. 1982. characteristics of sediment and organic carbon export for pristine boreal forest watersheds. Can. Jorn. Fish. Ac. Sc. 39:1699-1718. [ Links ]
37. PAUL, M.J., R.O. HALL. 2002. Particle transport and transient storage along a stream-size gradient in the Hubbard Brook Experimental Forest. Journal of North American Benthological Sosiety 21(2): 195-205. [ Links ]
38. PETERSON, R.C., K.W. CUMMINS & G.M. WARD. 1989. Microbial and animal processing of detritus in a woodland stream. Ecological Monographs. 59:21-39. [ Links ]
39. REEVES, G.H., L.E. BENDA, K.M. BURNETT, P.A. BISSON, & J.R. SEDELL. 1995. A Disturbance-Based Ecosystem Approach to Maintaining and Restoring Freshwater Habitats of Evolutionarily Significant Units of Anadromous Salmonids in the Pacific Northwest. American Fisheries Society Symposium 17:334-349. [ Links ]
40. RICHARDSON, J.S. 1992. Coarse particulate detritus dynamics in small, montane stream of southwertern British Columbia. Can. Jorn. Fish. Ac. Sc. 49:337-346. [ Links ]
41. RICHARDSON, J.S., R.E. BILBY & C.A. BONDAR. 2005. Organic matter dynamics in small streams of the Pacific Northwest. Journal of the American Water Resources Association 921-934. [ Links ]
42. SHANNON, J.P., D.W. BLINN, P.L. BENENATI & K.P. WILSON. 1996. Organic drift in a regulated desert river. Can. J. Fish. Aquat. Sci. 53:1360-1369. [ Links ]
43. TABACCHI E., E. CORREL, R. HAUER, G. PINAY, A. TABACCHI & R. WISSMAR. 1998. Development, maintenance and role of riparian vegetation in the river landscape. Fresh Water Biology 40:497-516. [ Links ]
44. TANNER, E.V. 1980. Studies on the biomass and productivity in a series of montane rain forests in Jamaica. Journal of Ecology 68:573-588. [ Links ]
45. TANNER, E.V.J., P.M. VITOUSEK & E. CUEVAS. 1998. Experimental investigation of nutrient limitation of forest growth on wet tropical mountains. Ecology 79(1):10-22. [ Links ]
46. VARGAS, J.P. 2003. Análisis de un bosque Altoandino con énfasis en la precipitación del mantillo: Quebrada La Vieja, Bogotá Colombia. Tesis en Biología. Universidad Nacional de Colombia, Bogotá. [ Links ]
47. VANNOTE, R.L., G.W. MINSHALL, K.W. CUMMINS, J.R. SEDELL, C.E. CUSHING. 1980. The river continuum concept. Can. J. Fish. Aquat. Sci. 37:130-137. [ Links ]
48. VARGAS, J.P. 2003. Análisis de un bosque Altoandino con énfasis en la precipitación del mantillo: Quebrada La Vieja, Bogotá, Colombia. Tesis en Biología. Universidad Nacional de Colombia, Bogotá. [ Links ]
49. VITOUSEK, P.M. & R.L. STANFORD, JR. 1986. Nutrient cycling in moist tropical forest. Annual Review of Ecology and Systematics 17:137-167. [ Links ]
50. WALLACE, J.B., M.R. WHILES, S. EGGERT, T.F. CUFFNEY, G.J. LUGTHART & K. CHUNG. 1995. Long term dynamics of coarse particulate organic matter in three Appalachian Mountain streams. J. N. Am. Benthol. Soc. 14:217-232. [ Links ]
51. WALLACE, J.B., J.W. GRUBAUGH & M.R. WHILES. 1996. Biotic indices and stream ecosystem processes: Results from an experimental study. Ecological Applications 6:140-151. [ Links ]
52. WARD, G.M. & N.G. AUMEN. 1986. Woody Debris as a Source of Fine Particulate Organic Matter in Coniferous Forest Stream Ecosystems. Canadian Journal of Fisheries and Aquatic Sciences 43:1635-1642. [ Links ]
53. WEBSTER, J.R. & B.C. PATTEN. 1979. effects of watershed perturbation on strean potassium and calcium dynamics. Ecological Monogrphs 49:51-72. [ Links ]
54. WEBSTER, J.R., E.F. BENFIELD. 1986. vascular plant breakdown in freshwater ecosystems. Annual Review of Eology and Syatematics 17:567-594. [ Links ]
55. WEBSTER, J.R., E.F. BENFIELD, T.P. EHRMAN, M.A. SCHAEFFER, J.L. TANK, J.J. HUTCHENS & D.J. D'ANGELO. 1999. What happens to allochthonous material that falls into streams? A synthesis of new and published information from Coweeta. Freshwater Biology 41: 687-705. [ Links ]
56. ZAH, R. & U. UEHLINGER. 2001. Particulate organic matter inputs to a glacial stream ecosystem in the Swiss Alps. Fresh Water Biology 46:1597-1608. [ Links ]
Recibido: 11/08/2007
Aceptado: 08/07/2008

