










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.31 no.1 Bogotá Jan./June 2009
Relationships between biomass and morphological characters of phytotelmata bromeliads in a Colombian upper Andean forest
CAROLINA ISAZA
JULIO BETANCUR
Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Apartado 7495, Bogotá, Colombia. isazacarolina@hotmail.com; jcbetancurb@unal.edu.co
RESUMEN
Se evaluaron algunas características morfológicas para seis especies de bromelias fitotelmata (Guzmania gloriosa, Racinaea tetrantha, Tillandsia biflora, T. complanata, T. fendleri y T. turneri) con el fin de establecer cuales características podrían utilizarse como estimadores indirectos de la biomasa total de la planta. El estudio se realizó en un bosque alto andino de la Sabana de Bogotá, Colombia, ubicado a 2900 m de altitud. Para cada especie se recolectaron entre 32 y 64 individuos en diferentes estadios de crecimiento, a cada uno de los cuales se le registró: 1) el número de hojas por roseta; 2) el perímetro de la base de la roseta; 3) la cobertura de la planta; 4) el largo de la lámina foliar; 5) la altura de la roseta; y 6) la biomasa total. Los resultados de la prueba Wilcoxon muestran que la cobertura fue la variable que más se relacionó con la biomasa. Una prueba de ajuste de curva mostró que la línea recta es la que más explica la relación entre la cobertura y la biomasa, permitiendo hacer estimaciones a partir de la ecuación sin ser necesario un muestreo destructivo.
Palabras clave. Andes, Biomasa, Bromeliaceae, Colombia, Epífitas, Fitotelmata, Morfometría.
ABSTRACT
Some morphological characteristics were evaluated for six bromeliad phytotelmic species (Guzmania gloriosa, Racinaea tetrantha, Tillandsia biflora, T. complanata, T. fendleri and T. turneri) to establish which of those can be used as indirect estimators of total biomass. The present study was performed in a high Andean forest of the Savanna of Bogotá, Colombia, at 2900 m. We collected between 32 and 64 individuals per species in different developmental stages. For each one we registered 1) number of leaves, 2) the basal rosette perimeter, 3) coverage, 4), leaf length 5) the height of the rosette; and 6) total biomass. According to the results, coverage was the variable most related with biomass. A curve fitting analysis showed that a straight line best explained the relationship between coverage and biomass. Thus, the equation allows estimating biomass from coverage without the destructive sampling effect.
Key words. Andes, Biomass, Bromeliaceae, Colombia, Epiphytes, Morphometry, Phytotelmata.
INTRODUCCIÓN
Las plantas epífitas son componentes importantes de los bosques tropicales, tanto por la riqueza como por la biomasa que aportan (Golley 1971, Nadkarni 1984, Gentry & Dodson 1987, Benzing 1990, Nieder et al. 2001, Merwin et al. 2003, Arévalo & Betancur 2004, 2006). En los bosques neotropicales la familia Bromeliaceae es uno de los grupos de epífitas vasculares más característicos (Kress 1986, Benzing 1990, Isaza et al. 2004). Las estrategias de localización de los nutrientes aseguran la supervivencia y el crecimiento de los individuos, algunas de las cuales son la alta acumulación de biomasa en el tejido foliar, la reducción de las raíces, la obtención de agua y nutrientes a través de la microcharca formada por sus hojas y la ruta metabólica CAM (Benzing 2000). Estas adaptaciones permiten a las bromelias sobrevivir en condiciones de estrés hídrico, mantener una alta tasa fotosintética, aún en condiciones de sombra, y una lenta acumulación de nutrientes (Skillman et al. 1999, Zotz et al. 2001).
Por otra parte, la biomasa y el área foliar en las bromelias están influenciadas por las adaptaciones ecofisiológicas de las especies (Pfitsch & Smith 1988, Skillman et al. 1999, Benzing 2000, Dunn 2000, Zotz et al. 2001, Hietz et al. 2002, Pett-Ridge & Silver 2002, Freitas et al. 2003, Merwin et al. 2003). Por ejemplo, en los individuos juveniles de bromelias fitotelmata (tipo tanque) algunas de las adaptaciones se manifiestan en bajas tasas de crecimiento y poca acumulación de agua y nutrientes. Este comportamiento se acentúa en las bromelias de los bosques montanos, producto de las fluctuaciones extremas en la temperatura, de los períodos de intensidad lumínica cortos, del estrés hídrico durante la estación seca y del área del forofito disponible para su crecimiento (Pfitsch & Smith 1988, Zotz et al. 2001).
En la ecología de comunidades se utilizan diferentes índices que estiman la diversidad y el recambio entre ellas (diversidad beta), para lo que se requiren datos de abundancia relativa de las especies dentro de la comunidad (Magurran 1988). La abundancia de las especies vegetales puede ser medida a través de su cobertura proyectada, el número de individuos o su peso (biomasa). En comunidades de epífitas no vasculares la abundancia se ha estimado a través del área de la superficie proyectada (Nadkarni 1984, Hofstede et al. 1993, Wolf 1994, Freiberg & Freiberg 2000, Nadkarni 2000, Ingram & Nadkarni 1993, Merwin et al. 2003), mientras que en epífìtas vasculares este procedimiento es poco práctico ya que el tamaño de las plantas muchas veces excede el diámetro de la rama sobre la que crecen.
Por lo tanto, en muchos estudios la abundacia de epífitas se expresa como el número de individuos, pero en bromelias clonales densamente agrupadas es difícil discernir los individuos (genets) de los vástagos laterales de un mismo individuo (ramets), aunque para estimar la biomasa este aspecto no es relevante. Por otra parte, la estimación directa de la biomasa puede implicar métodos destructivos que afectan la conservación de las especies. Para minimizar los efectos destructivos del muestreo, usualmente se hacen extrapolaciones del peso seco derivadas de las características morfológicas de las plantas (Isaza et al. 2004, Serna-Isaza 1994, Zotz et al. 2001, Wolf et al. 2009).
En bromelias fitotelmatas la altura de la roseta, la longitud de la lámina foliar, el área de la superficie de la planta y el diámetro del tanque parecen estar correlacionadas con el peso de la planta (Zotz & Thomas 1999, Merwin et al. 2003). Sin embargo, hasta el momento no se conocen pruebas formales que permitan establecer cual de estos parámetros morfométricos estima mejor la biomasa. Así, el objetivo principal de este estudio es explorar la relación entre la biomasa y algunas características morfológicas de las bromelias, con la intensión de poder estimar indirectamente la biomasa sin necesidad de utilizar métodos destructivos.
ÁREA DE ESTUDIO
El estudio se desarrolló en un bosque alto andino localizado en la Hacienda "El Santuario", municipio de Chocontá, departamento de Cundinamarca (Colombia), ubicado a 2900 m de altitud, a 5º 01' N y 73º 42' W. La precipitación media anual es de 924.7 mm, con un régimen unimodal que presenta una época de lluvias entre los meses de mayo y agosto y una época seca entre los meses de septiembre y abril. La temperatura media anual es de 11.9º C y, según el sistema de zonas de vida, el sitio se clasifica como bosque húmedo montano (bh-M) (IGAC 1977).
El bosque estudiado posee cerca de 3500 árboles/ha con DAP = 1 cm y un área basal promedio de 51.35 m2/ha. El dosel alcanza una altura promedio de 15 m, una apertura promedio de 5.67 % y presenta algunos árboles emergentes de hasta 25 m de altura. Las especies arbóreas ecológicamente más importantes son Weinmannia tomentosa, Drymis granadesis y Myrsine cf. ferruginea (observaciones personales).
MÉTODOS
El muestreo se realizó entre los meses de agosto y diciembre de 2001. Se muestrearon seis especies de bromelias, pertenecientes a la subfamilia Tillandsioideae: Guzmania gloriosa (André) André ex Mez, Racinaea tetrantha (Ruiz & Pav.) M. A. Spencer & L. B. Sm., Tillandsia biflora Ruiz & Pav., T. complanata Benth., T. fendleri Griseb. y T. turneri Baker.
Durante la fase inicial del estudio se identificaron los diferentes estados de desarrollo para cada una de las especies, de forma tal que se pudieran identificar éstas sin necesidad de que estuvieran florecidas. Para cada especie se recolectaron de 32 a 64 individuos que representaran el rango total de tamaño para cada una de ellas, sin distinguir su procedencia (genets o ramets). A cada individuo se le midió las siguientes variables morfométricas: 1) el perímetro de la base de la roseta, medido en el sector donde terminan las vainas e inician las láminas foliares; 2) la cobertura superior de la planta, estimada idealmente como el área de la elipse, para lo cual se midió los radios mayor (a) y menor (b), y se transformó según la ecuación A = pab; 3) la altura de la roseta; 4) el número de hojas vivas; 5) el promedio aritmético de la longitud de las cinco láminas foliares vivas más periféricas, sin incluir la vaina. Los valores de las variables morfométricas (media y error estándar de la media) pueden consultarse en un estudio anterior realizado en el mismo sitio (Isaza et al. 2004).
Para determinar la biomasa de las rosetas (hojas completas y eje de inserción), definida como el peso en seco de la materia viva (Nadkarni 1984), se removieron las raíces, la materia orgánica acumulada dentro del tanque, las hojas muertas y las inflorescencias. Posteriormente, se secaron en un horno eléctrico a 70ºC, durante 36 horas continuas.
La relación entre las variables morfométricas mencionadas y la biomasa, se determinó a través de una prueba Wilcoxon, puesto que la mayor parte de los grupos de datos no mostraron distribución normal (P > 0.05) (Zar 1999). Posteriormente, realizamos un análisis que nos permitiera describir la relación entre la biomasa y las variables que presentaran una correlación significativa. Para ello utilizamos la prueba de ajuste de la curva ("curve fitting") con el paquete estadístico XL-plot (http://www.bram.org/xlplot/xlplot.php).
RESULTADOS
La prueba Wilcoxon muestra que la cobertura fue la única variable morfométrica que presentó correlación significativa con la biomasa para todas las especies (P< 0.000). Así mismo, la altura de la roseta también mostró fuerte correlación con la biomasa en todas las especies, excepto en T. fendleri (Tabla 1).
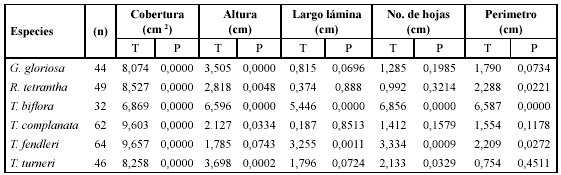
El análisis de ajuste de la curva mostró que la relación entre la cobertura y la biomasa se ajusta a una correlación lineal positiva, en donde para todas las especies el coeficiente de correlación estuvo entre 0.530 y 0.791, con P< 0.0005 (Tabla 2).
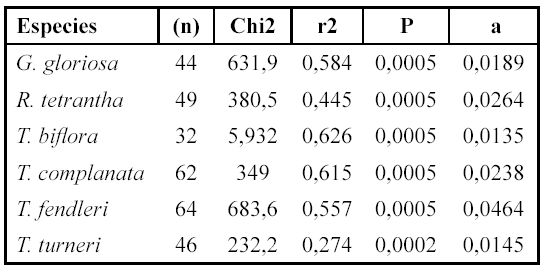
Realizamos regresiones lineales para cada especie con el fin de obtener el valor de la pendiente y poder estimar la biomasa a partir de la cobertura. Para ello usamos la ecuación de la línea recta y = ax + b, donde y = biomasa, x = cobertura, a = pendiente y b = intercepto sobre el eje y (Zar 1999) (Tabla 2). Además, comparamos los valores de la biomasa encontrada con los esperados, utilizando la ecuación de la línea recta. Para ello escogimos cuatro individuos al azar por especie y comparamos los valores de biomasa esperada y estimada (Tabla 3).
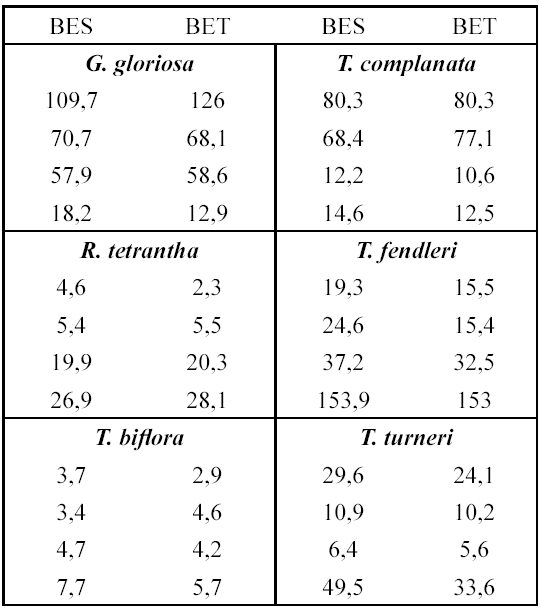
En general estos valores fueron bastante cercanos entre sí, lo que refuerza los resultados obtenidos en este estudio. Así, la cobertura nos permite hacer estimaciones indirectas de la biomasa en las poblaciones silvestres de las bromelias censadas.
DISCUSIÓN
Aunque se asuma que cualquier medida de alguna parte u órgano de la planta, como el área foliar, el número de hojas y el largo de las mismas, entre muchas otras, podrían estar correlacionadas directamente con la biomasa, no se cuenta con estudios que permitan comparar directamente esta relación. Los resultados de este estudio indican que sólo algunas de las características morfométricas estudiadas se relacionan significativamente con la biomasa y que, además, cada especie muestra comportamientos específicos (Tablas 1). Algunos estudios indican que ciertas partes de una bromelia pueden tender a crecer asincrónicamente respecto al resto de la planta, dependiendo de la cantidad de luz a la que está expuesta, siempre y cuando no haya restricciones de humedad (Cervantes et al. 2005). Además, muchas bromelias, especialmente las fitotelmata, aletargan su crecimiento en condiciones de sequía (Laube & Zotz 2003).
Por otra parte, el hecho de que la cobertura fue la única variable que se relacionó con la biomasa en todas las especies, puede deberse a que indica el tamaño de la planta. La cobertura está involucrada en la acumulación de agua y detritus (elementos que le proporcionan a la planta los nutrientes necesarios para su crecimiento) y, por ende, en el incremento en biomasa (Zotz & Thomas 1999). De hecho, este estudio muestra como la cobertura permite hacer estimaciones indirectas de la biomasa.
Cervantes et al. (2005) registraron que existe una relación directa entre el número de hojas y la cantidad de luz recibida por una bromelia epífita, pero que el número de hojas no depende necesariamente del estado de desarrollo de la planta. Lo anterior puede explicar la poca relación encontrada entre el número de hojas y la biomasa en la mayoría de las especies (Tabla 1). Así por ejemplo, una especie de gran tamaño como G. gloriosa puede tener menor número de hojas que una de tamaño reducido como T. biflora (Isaza et al. 2004).
La mayor parte de los estudios realizados con epífitas en el Neotrópico se han concentrado en cuantificar la riqueza de especies y la abundancia (Nieder et al. 2000, Merwin et al. 2003, Arévalo & Betancur 2004). Sin embargo, pocos estudios relacionan la estimación de la biomasa a través del empleo de variables morfométricas, como tampoco su participación en la sucesión vegetal y en la regeneración de los bosques. Los datos presentados por este estudio apuntan a conocer la dinámica de la biomasa de las epífitas en los bosques montanos, los cuales son especialmente frágiles y en los que las bromelias son especies clave y albergan mayor biomasa que en los de tierras bajas (Coxson & Nadkarni 1995).
Los resultados aquí presentados pueden servir de base para desarrollar métodos alternativos que permitan estimar la biomasa de las bromelias sin que sea necesario extraer las plantas de su medio natural. Estas consideraciones son bastante importantes para su conservación, pues se ha observado que la epífitas pueden incrementar rápidamente su abundancia en corto tiempo después de la colonización inicial, mientras que su incremento en biomasa es un proceso mucho más lento. Además, requieren de la presencia de forófitos maduros para que se establezcan comunidades epífitas complejas (Merwin et al 2003).
LITERATURA CITADA
1. ARÉVALO R. & J. BETANCUR. 2004. Diversidad de epífitas vasculares en cuatro en cuatro bosques del sector suroriental de la serranía de Chiribiquete, Guayana Colombiana. Caldasia 26(2): 359-380. [ Links ]
2. ARÉVALO R. & J. BETANCUR. 2006. Vertical distribution of vascular epiphytes in four forests types of the Serranía de Chiribiquete, Colombian Guayana colombiana. Selbyana 27 (2): 175-185. [ Links ]
3. BENZING, D. H. 1990. Vascular Epiphytes: General Biology and Related Biota, 1st ed. Cambridge University Press, Nueva York. [ Links ]
4. BENZING, D. H. 2000. Bromeliaceae. Profile of an Adaptive Radiation. Cambridge University Press, Cambridge. [ Links ]
5. CERVANTES, S.E., E. A. GRAHAM & J. L. ANDRADE. 2005. Light microhabitats, growth and photosynthesis of an epiphyte bromeliad in a tropical dry forest. Plant Ecology 179: 107-118. [ Links ]
6. COXSON, D. S. & N. M. NADKARNI. 1995. Ecological roles of epiphytes in nutritent cycles of forest ecossystems pp. 27-44. In M. D. Lowman & N. M. Nadkarni (eds.). Forest canopies. Academic Press, San Diego. [ Links ]
7. DUNN, R. R. 2000. Bromeliad communities in isolated trees and three succesional stages of an Andean cloud forest in Ecuador. Selbyana 21: 137-143. [ Links ]
8. FREIBERG, M. & E. FREIBERG. 2000. Epiphyte diversity and biomass in the canopy of lowland and montane forest in Ecuador. Journal of Tropical Ecology 16: 673-688. [ Links ]
9. FREITAS DE, C. A., F. R. SCARANO & D. D. BIESBOER. 2003. Morphological variation in two facultative epiphytic bromeliads growing on the floor of a swamp forest. Biotropica 35(4): 546-550. [ Links ]
10. GENTRY, A. H & C. H. DODSON. 1987. Diversity and biogeography of neotropical vascular epiphytes. Ann. Missouri Bot. Gard. 74: 205- 233. [ Links ]
11. GOLLEY, F., J. MCGINNIS & R. CLEMENTS. 1971. La biomasa y la estructura de algunos bosques de Darién, Panamá. Turrialba 21: 189-196. [ Links ]
12. HIETZ P., J. AUSSERER & G. SCHINDLER. 2002. Growth, maturation and survival of epiphytic bromeliads in a Mexican cloud forest. Journal of Tropical Ecology 18: 177-191. [ Links ]
13. HOFSTEDE, R.G. M., J. D. H. WOLF & D. H. BENZING. 1993. Epiphytic biomass and nutrient status of a Colombian upper Montane rain forest. Selbyana 14: 37-45. [ Links ]
14. IGAC. 1977. Zonas de vida o formaciones vegetales de Colombia . Memoria explicativa sobre el mapa ecológico. Vol. XIII, No. 11. Instituto Geográfico Agustín Codazzi, Bogotá. [ Links ]
15. INGRAM, S. W. & N. M. NADKARNI.1993. Composition and distribution of epiphytic organic matter in a Neotropical cloud forest, Costa Rica. Biotropica 24(4): 370-383. [ Links ]
16. ISAZA, C., J. BETANCUR & J. V. ESTÉVEZ-VARÓN. 2004. Vertical distribution of bromeliads in a Montane forest in the eastern cordillera of the Colombian Andes. Selbyana 25(1): 126-137. [ Links ]
17. LAUBE, S. & G. ZOTZ. 2003. Which abiotic factors limit vegetative growth in a vascular epiphyte? Functional Ecology 17: 598-604. [ Links ]
18. KRESS, W. J. 1986. The systematic distribution of vascular epiphytes: an update. Selbyana 9: 2-22. [ Links ]
19. MAGURRAN, A.E. (1988) Ecological diversity and its measurement Princeton University Press, Princeton, Nueva Jersey. [ Links ]
20. MERWIN, M. C., S. A. RENTMEESTER & N. M. NADKARNI. 2003. The influence of host tress species on the distribution of epiphytic Bromeliads in experimental monospecific plantations, La Selva, Costa Rica. Biotropica 35(1): 37-47. [ Links ]
21. NADKARNI, N. M. 1984. Epiphyte biomass and nutrient capital of a Neotropical elfin forest. Biotropica 16: 249-256. [ Links ]
22. NADKARNI, N. M. 2000. Colonization od stripped branch surfaces by epiphytes in a lower Montane cloud forest, Monteverde, Costa Rica. Biotropica 32(2): 358-363. [ Links ]
23. NIEDER, J., S. ENGWALD, M. KLAWUN, & W. BARTHLOTT. 2000. Spatial distribution of vascular epiphytes (including hemiepiphytes) in a lowland Amazonian rain forest (Surumoni crane plot) of Southern Venezuela. Biotropica 32: 385-396 [ Links ]
24. NIEDER, J., J. PROPERI & G. MICHALOUD. 2001. Epiphytes and their contribution to canopy diversity. Plant Ecology 153: 51- 63. [ Links ]
25. PFITSCH, W. A., & A. P. SMITH. 1988. Growth and photosynthesis of Aechmea magdalenae, a terrestrial CAM plant in a moist tropical forest, Panama. Journal of Tropical Ecology 4:199-207. [ Links ]
26. PETT-RIDGE, J. & W. L. SILVER. 2002. Survival, growth and ecosystems dynamics of displaced Bromeliads in a montane tropical forest. Biotropica 32(2): 211-224. [ Links ]
27. SERNA-ISAZA., R. A.1994. Distribución vertical de epifitas vasculares en un relicto de bosque de Weinmannia tomentosa y Dryimis granadensis en la región de Monserrate, Cundinamarca-Colombia. Pages 521-543. En: Mora-OSEJO, L. E. & H. STURM (eds.), Estudios Ecológicos del Páramo y del Bosque Altoandino Cordillera Oriental de Colombia. Tomo II. Academia Colombiana de Ciencias Exactas, Físicas y Naturales. Colección Jorge Álvarez Lleras No. 6. [ Links ]
28. SKILLMAN, J.B., M. GARCIA, & K. WINTER. 1999. Whole-plant consequences of Crassulacean acid metabolism in an understory terrestrial bromeliad. Ecology 80:1584-1593. [ Links ]
29. WOLF, J.H.D. 1994. Factors controlling the distribution of vascular and nonvascular epiphytes in the northern Andes. Vegetatio 112: 15-28. [ Links ]
30. WOLF, J.H.D., GRADSTEIN, S.R., & NADKARNI, N.M. 2009. A protocol for sampling of vascular epiphyte richness and abundance. Journal of Tropical Ecology, 25, 107-121. [ Links ]
31. ZAR, J. H. 1999. Biostatistical Analysis. Fourth edition. Prentice Hall. Upper Saddle River, Nueva Jersey. [ Links ]
32. ZOTZ, G. & V. THOMAS. 1999. How much is in the tank? Model for two epiphytic bromeliads. Annals of Botany 83: 183-192. [ Links ]
33. ZOTZ, G, P. HIETZ & G. SCHMIDT. 2001. Small plants, large plants - the importance of plant size for the physiological ecology of vascular epiphytes. Journal of Experimental Botany 52:2051-56. [ Links ]
Recibido: 04/09/2006
Aceptado: 14/04/2009

