










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.31 no.1 Bogotá Jan./June 2009
Short-term temporal changes in the spatial patterns of nine species of common plants in an Andean cloud forest in southwestern Colombia
MARTHA ISABEL VALLEJO
GLORIA GALEANO
Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Apartado 7495, Bogotá D. C., Colombia. mivallejoj@unal.edu.co; gagaleanog@unal.edu.co
RESUMEN
Se analizaron los cambios en los patrones de distribución espacial de nueve especies de plantas comunes en un bosque pluvial premontano, entre dos censos realizados en una parcela permanente de 25 ha. Durante el primer censo efectuado en 1997 se marcaron, se midieron y se ubicaron en un mapa, todos los tallos leñosos con un DAP = 1 cm; transcurridos cinco años se realizó un segundo censo y se incluyeron datos de mortalidad y reclutamiento. Mediante pruebas de estadística espacial usando la función K(t) de Ripley, se detectaron los patrones de distribución de las especies estudiadas y las relaciones de dependencia de dos tipos de eventos: árboles grandes (DAP = 10 cm) vs. árboles pequeños (DAP < 10 cm), e individuos vivos vs. individuos muertos. Se puso a prueba la hipótesis de la mortalidad aleatoria, con el fin de determinar si la tendencia a la sobre-dispersión de los árboles grandes sobrevivientes podía ser atribuida a la mortalidad selectiva de los árboles pequeños. Los resultados corroboran hallazgos de otras investigaciones sobre la tendencia al agrupamiento en las clases de tamaño más pequeñas, pero no en las clases de DAP = 10 cm, las cuales tuvieron un patrón aleatorio a escalas pequeñas (t < 10 m de distancia). Dos especies, Otoba lehmanii (Myristicaceae) y Matisia bolivarii (Bombacaceae), apoyan claramente las hipótesis de la densidad y/o de la distancia propuestas por Janzen & Connell, pero se infiere que los procesos que llevaron a la sobre-dispersión de los árboles grandes en estas especies fueron diferentes. En O. lehmanii se atribuyó a procesos dependientes de la distancia por requerimientos de luz, y en M. bolivarii se atribuyó a procesos denso-dependientes debido al ataque de patógenos en estadios tempranos. Al final, se menciona el aporte de esta investigación a la discusión sobre la teoría neutral de la biodiversidad y biogeografía.
Palabras clave. Ecología de comunidades, ecología de plantas, hipótesis de la mortalidad aleatoria, K de Ripley, patrones espaciales.
ABSTRACT
The changes in the spatial distribution of nine common species of an Andean cloud forest were analyzed between two censuses carried out in a 25-ha plot. In the first census in 1997, all woody stems with DBH =1 cm were tagged, measured and plotted. After five years, a second census was made and data of mortality and recruitment were recorded. Through spatial statistical tests using Ripley's K (t) function, the patterns of distribution were detected in each species, as well as the relationship between two types of events: big trees (DBH = 10 cm) vs. small trees (DBH < 10 cm) and surviving individuals vs. dead individuals. The random mortality hypothesis was tested to determine if the tendency to over dispersion of the big trees can be attributed to the selective mortality of the small trees. The results corroborate findings of other studies about the aggregation tendency for the smallest size classes, but not for those with DBH = 10 cm, which had a random pattern at small scales. Otoba lehmanii (Myristicaceae) and Matisia bolivarii (Bombacaceae) clearly supported the density and/or distance hypothesis proposed separately by Janzen and Connell, but it was inferred that the processes that led to the over dispersion of the big trees were different. The pattern in O. lehmanii was attributed to distance-dependent processes related to light requirements, and in M. bolivarii to density-dependent processes due to pathogen attack in early stages. Finally, the contribution of this research to the discussion on the neutral theory of biodiversity and biogeography is discussed.
Key words. Community ecology, plant ecology, random mortality hypothesis, Ripley's K-function, spatial patterns.
INTRODUCCIÓN
A comienzos del presente siglo el ecólogo Stephen Hubbell (2001) dio a conocer su teoría neutral unificada de la biodiversidad y la biogeografía. Esta teoría pretende explicar la diversidad y la abundancia relativa de las especies en las comunidades ecológicas, bajo la presunción de que las diferencias entre miembros de una comunidad ecológica de especies tróficamente similares son "neutrales" o irrelevantes a sus procesos. De acuerdo con la definición de la palabra "neutral", descrita como la equivalencia ecológica per capita de todos los individuos de todas las especies de una comunidad tróficamente definida, su modelo está concebido prácticamente a nivel de individuos, y asume que todos los individuos de cada especie obedecen exactamente las mismas reglas de compromiso ecológico, por lo que si todas las especies de un mismo nivel trófico se comportan de una manera similar (e.g. reproducción y muerte), los individuos también lo hacen en el mismo sentido.
Visto en un contexto espacial, se puede decir que los planteamientos de Hubbell (2001) sugieren que la distribución actual de los organismos no son el resultado de procesos determinísticos sino estocásticos, es decir, que su establecimiento estaría determinado por la entrada y la salida de los individuos en una comunidad, producto de eventos aleatorios que ocurren durante la historia de vida de las especies que permanecen en un sitio en particular. Así, procesos como la competencia, la especialización de hábitat, la dispersión limitada o la mortalidad aleatoria, no serían los que determinan que el patrón de distribución de las poblaciones sea uniforme, agrupado o aleatorio, sino la suma de todos los efectos que se derivan de dichos procesos; lo cual no es otra cosa distinta que la entrada y la salida por nacimiento, migración, muerte o extinción.
Si los planteamientos de la teoría neutral son ciertos, uno esperaría encontrar al interior de una comunidad de plantas que habitan un área determinada, poblaciones de especies distribuidas de una manera aleatoria, siempre y cuando el ambiente local sea homogéneo, es decir, sin variaciones físicas y climáticas de gran impacto que pudiesen haber direccionado los procesos de evolución de algunas especies hacia la ocupación de hábitat especializados. O bien, un mosaico de patrones espaciales donde priman los aleatorios por encima de los uniformes y los agrupados.
Para mayor claridad, los principales mecanismos que se han planteado para explicar los patrones de distribución actual de los árboles al interior de una comunidad pueden resumirse en tres: 1-procesos denso-dependientes (e.g. competencia) asociados a patrones de distribución uniforme; 2- estocasticidad demográfica por mortalidad aleatoria, asociada a patrones de distribución aleatoria; y 3- dispersión limitada y/o especialización de hábitat, asociada a patrones agrupados o distribución por parches. Estos mecanismos hacen parte de teorías ampliamente conocidas que han intentado explicar la alta riqueza de especies en los bosques tropicales (Ashton 1969, Janzen 1970, Connell 1971, King & Pimm 1983, Hubbell & Foster 1983), así como los procesos que han llevado a la distribución espacial actual de las poblaciones de plantas (Hubbell 1979, Armesto et al. 1986, Lieberman & Lieberman 1994).
La denso-dependencia o la dependencia de la densidad, son mecanismos inicialmente planteados por Janzen (1970) y Connell (1971) como procesos que derivan en una distribución uniforme de árboles adultos debido a la mortalidad de semillas y plántulas cercanas a adultos de la misma especie o en sitios con una alta densidad de juveniles co-específicos. Dichos procesos han sido corroborados en varios estudios con especies particulares, pero también han sido refutados por diversos autores que evidenciaron una tendencia al agrupamiento en árboles adultos. Dentro de los mecanismos propuestos para explicar los patrones agrupados, se encuentran la especialización en hábitats disponibles para sobrevivir o crecer, la regeneración en claros de bosque y la dispersión limitada (Hubbell 1980, Brokaw & Busing 2000, Condit et al. 2000, Lawes & Obiri 2003). Sin embargo, la distribución per se de los árboles adultos no necesariamente prueba que las hipótesis de Janzen & Connell estén operando. Por esta razón, otros investigadores, como Sterner y sus colegas (1986), decidieron observar los patrones de distribución por clases de edad simulando la mortalidad aleatoria de los tallos más jóvenes, con el fin de explorar si la distribución en el espacio estaba asociada a la supervivencia, crecimiento o dispersión de las poblaciones. Ellos encontraron una clara evidencia de sobre-dispersión o distribución uniforme de los árboles adultos por procesos diferentes a la mortalidad aleatoria.
Por otra parte, la competencia es uno de los procesos más difíciles de probar en organismos sésiles como las plantas, debido a lo complicado que resulta detectar interacciones entre los individuos; por eso, la observación de los cambios en los patrones de distribución espacial, se constituye en una buena herramienta para evidenciarla. Por ejemplo, en un bosque maduro formado por individuos de diversas edades y tamaños, Oliver & Larson (1996) encontraron que a medida que un árbol se hace dominante en el dosel, aumenta el espacio que ocupa y su capacidad para seguir creciendo, mientras que cuando un árbol alcanza la categoría de subordinado se encuentra confinado a un espacio constante o incluso cada vez más reducido. Los patrones espaciales de los árboles dominantes y subordinados son divergentes en este sentido, predominando un patrón aleatorio o uno regular para árboles dominantes, usualmente asociados con grandes diámetros, y un patrón agregado para los subordinados, pertenecientes a clases de diámetro menores (Ward et al. 1996).
Teniendo en cuenta lo anterior, el presente artículo pretende analizar los cambios en los patrones de distribución de las especies seleccionadas al interior de una parcela de 25 ha, con el propósito de determinar si alguno de los mecanismos antes enunciados puede ser inferido. Para hacerlo, se analizaron los eventos de mortalidad de los individuos pertenecientes a nueve especies de plantas comunes en un bosque nublado andino, separados en dos clases de tamaño: árboles pequeños, definidos como aquellos tallos con un diámetro a la altura del pecho (DAP) entre 1 y 10 cm y árboles grandes, aquellos con un DAP = 10 cm. Esta diferenciación por clases diamétricas es, en términos generales, equivalente a los juveniles y a los adultos mencionados en las hipótesis de densidad-dependencia (Janzen 1970, Connell 1971), sin que esto implique que el tamaño de los individuos de las especies seleccionadas esté estrictamente asociado a la edad reproductiva de los mismos.
Con base en los eventos de mortalidad registrados en dos censos realizados en la parcela, se pone a prueba una predicción que relaciona los patrones de distribución de las dos clases de tamaño definidas y la hipótesis de la mortalidad aleatoria propuesta por Sterner y sus colegas (1986). Esta última plantea que la distribución de los árboles adultos, generalmente asociados con clases diamétricas grandes, no difiere significativamente de la esperada por procesos de mortalidad aleatoria de individuos en estadios de temprana edad, tales como semillas, plántulas o juveniles; es decir, que si la mortalidad en los estadios más tempranos ocurre al azar, la distribución de los tallos grandes también debería ser azarosa. En caso contrario, alguno de los mecanismos alternativos propuestos podría ser el causante del patrón espacial observado en las clases diamétricas mayores, para lo cual el análisis de los resultados obtenidos se soporta en la información disponible sobre la biología y la ecología de las especies estudiadas. Si múltiples mecanismos están ocurriendo (incluidos procesos estocásticos debido a mortalidad aleatoria), producirían patrones espaciales diversos, y probablemente los planteamientos de Hubbell tienen sentido y ayuden a comprender cómo y por qué las comunidades de plantas se encuentran organizadas al interior del bosque.
Para reducir el efecto de factores enmascarados en la heterogeneidad ambiental sobre los patrones de distribución espacial esperados, se tuvo en cuenta la escala de observación y se siguieron las recomendaciones de Sterner et al. (1986), al considerar varios criterios de selección de las especies y del sitio de estudio, como por ejemplo la escogencia de especies de larga vida, no clonales, con una estructura de edades evidente, que estuvieran en pie y que sus semillas fueran dispersadas en altas densidades iniciales sobre un sustrato homogéneo (Sterner et al. 1986, Kenkel 1988).
Área de estudio
El estudio se llevó a cabo en una parcela permanente de 25 ha establecida en 1996 por el Instituto de Investigación de Recursos Biológicos "Alexander von Humboldt", en cooperación con la Fundación FES y COLCIENCIAS. La parcela, que cuenta con un rango de elevación entre 1796 y 1891 msnm, hace parte de la Reserva Natural La Planada, una reserva privada ubicada en la vertiente occidental de la Cordillera Occidental de los Andes, al sur occidente de Colombia (1º17'Norte, 78º15'Oeste). La Planada se extiende sobre una depresión alargada siguiendo la dirección de la cordillera, con una variación en altitud que va desde 1300 hasta 2100 msnm (Vallejo et al. 2004). La vegetación de la región corresponde a un bosque pluvial premontano (bp-PM) de acuerdo con la clasificación de Holdridge (1982).
La estructura del bosque la conforman cinco capas definidas por el tipo de vegetación predominante: árboles emergentes (20-25 m de altura), árboles de dosel (12-15 m), arbolitos (5-8 m), arbustos (2-4 m), y plantas bajas del sotobosque incluyendo hierbas (0-2 m). La altura promedio del dosel es de 15 m. Los árboles emergentes pueden exceder los 25 m en altura y 1.5 m en DAP. Las hemiepífitas, que incluyen árboles estranguladores y lianas, son muy comunes en La Planada y colonizan casi siempre los árboles emergentes. Una de las características estructurales más importantes de la reserva es la presencia de epífitas (Gentry 1995), mientras que la capa densa del sotobosque está dominada por especies de la familia Rubiaceae, principalmente de los géneros Faramea y Palicourea.
Los suelos de La Planada, derivados de materia orgánica y cenizas volcánicas, se encuentran saturados de aluminio en el horizonte superficial (>60 %); esto, sumado a los altos niveles de precipitación registrados por más de 15 años (~ 4.400 mm/año) (Vallejo et al. 2004) y a las bajas concentraciones de fósforo (21,6 mg/Kg de suelo), hacen que los suelos de La Planada sean pobres en nutrientes, por lo que se descarta la posibilidad de que hayan tenido algún tipo de aprovechamiento agrícola en épocas anteriores. Sin embargo, se tiene conocimiento de que antes de ser declarada Reserva Natural, los primeros predios que conformaron La Planada fueron utilizados para el aprovechamiento selectivo de especies maderables y la explotación de especies para producir carbón de leña (Samper & Vallejo 2007).
Materiales y Métodos
Recolección de datos. El establecimiento de la parcela consistió, en primera instancia, en hacer el levantamiento topográfico de las 25 ha con curvas de nivel cada metro. Luego el terrero se dividió en 625 cuadrantes de 20 m x 20 m, los cuales a su vez fueron divididos en sub-cuadrantes de 5 m x 5 m. Una vez establecida la parcela, se inició el primer censo de plantas en 1997. Esta fase consistió en marcar con una placa de aluminio numerada cada tallo de DAP = 1 cm, luego medir su diámetro, y finalmente ubicarlo en un mapa a nivel de cuadrante (Condit 1998, Vallejo et al. 2005). El segundo censo, realizado entre los años 2002 y 2003, consistió en registrar los eventos de mortalidad, reclutamiento y crecimiento de cada uno de los individuos censados.
Selección de las especies. La escogencia de las especies se hizo teniendo en cuenta las características sugeridas por Sterner et al. (1986): 1- que las especies fueran comunes en la parcela (N>50 individuos, 2 indiv/ha), 2- que sus frutos fueran medianos (2-5 cm en diámetro o longitud) o grandes (> 5 cm), asumiendo que el tamaño es una variable directamente asociada con la estrategia de dispersión de las semillas; es decir, que frutos grandes probablemente caen por su propio peso muy cerca del árbol "madre" formando parches de semillas bajo la copa del mismo, 3- que no fueran especies clonales, 4- que tuvieran suficiente representatividad de individuos en diversas clases de tamaño, y, 5- que estuvieran distribuidas sobre un sustrato homogéneo, asumiendo poca variabilidad en los componentes del suelo y en la topografía del sitio. Las características de las especies seleccionadas se detallan en la Tabla 1.
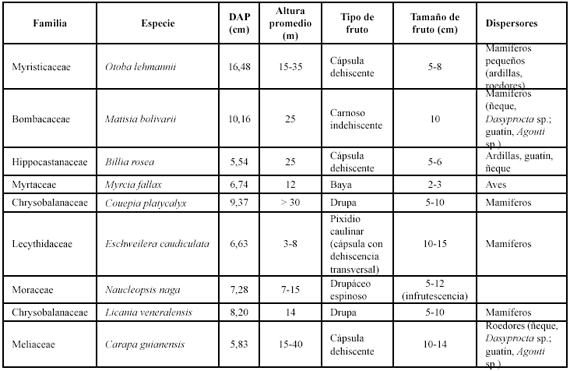
Procesamiento de la información y análisis de datos. Los aspectos estructurales como la abundancia, la diversidad y el área basal, así como la información demográfica determinada por las tasas de mortalidad, reclutamiento y crecimiento promedio anual de las poblaciones fueron obtenidas mediante la metodología descrita en Samper & Vallejo (2007).
Los patrones de distribución de las poblaciones estudiadas se efectuaron mediante la función Ripley K(t), una herramienta ampliamente conocida para analizar patrones de puntos espacialmente referenciados en un mapa de un área de estudio predeterminada y usualmente registrados en dos dimensiones (x,y). Esta función se emplea para medir la distancia de un punto a todos sus vecinos en un área específica y resumirla en un patrón de distribución (agrupado, aleatorio o uniforme). También se pueden usar análisis bivariados o multivariados para describir las relaciones entre dos o más patrones de puntos o eventos (Dixon 2002).
Para determinar la distribución espacial de los individuos de cada una de las especies se empleó el análisis univariado K (t) definido por la ecuación:
en donde A es el área de estudio (m2) representada por el área de la parcela; Wj es un factor de peso usado para reducir el problema del efecto de borde; It es un contador que toma el valor de 1 cuando la distancia entre i y j es menor que t y 0 en el caso contrario y n es el número de eventos (puntos) en A, es decir, la densidad.
El resultado es presentado como una transformación de la raíz cuadrada de K(t) conocido como L(t) y definido por la ecuación:
L(t) permite interpretar K(t) de una manera más fácil porque estabiliza la varianza y es una función lineal de t. Así, una desviación negativa de L(t) indica un patrón agrupado de los puntos, mientras que una desviación positiva indica un patrón regular. Los intervalos de confianza se calcularon mediante simulaciones de Monte Carlo (99 en total) para asegurar un nivel de significancia de 0.01.
Para comparar la distribución entre clases de tamaño y entre estatus de vida se empleó un análisis bivariado K12(t), que es análogo a K(t) pero permite observar la dependencia espacial de dos eventos diferentes a varias distancias (t) del vecindario inmediato. Igual que con el análisis univariado se construyeron los intervalos de confianza mediante simulaciones de Monte Carlo, que se emplean para analizar las desviaciones de L12(t). Si la desviación es negativa es porque existe "atracción" entre los dos eventos y si es positiva se habla de "repulsión". De acuerdo con Peterson & Squiers (1995), la repulsión se define como una tendencia de los puntos (árboles) de dos eventos diferentes a estar más separados de lo que estarían si ellos estuvieran distribuidos independientemente uno del otro.
La hipótesis de la mortalidad aleatoria fue evaluada como se describe a continuación: 1- se separaron los tallos pequeños de los grandes y se calculó la probabilidad de supervivencia de los tallos pequeños para cada especie; 2- mediante un proceso conocido como dispersión aleatoria (Random Thinning, Baddeley & Turner 2005), cada individuo de los tallos pequeños fue sacado o incluido de la población inicial, resultando en un patrón de puntos que fueron retenidos o que sobrevivieron; 3- se calculó L(t) para los sobrevivientes; 4-se repitió 99 veces el proceso de dispersión aleatoria y se construyeron los intervalos de confianza del 99%; y 5- se graficó la distribución de los tallos grandes sobrevivientes con los límites de confianza generados a partir de las simulaciones. Si la función empírica cae por fuera de los límites de confianza significa que existe una desviación significativa de la hipótesis de la mortalidad aleatoria, es decir, determina si la distribución de los tallos grandes sobrevivientes difiere o no significativamente de la esperada por procesos de mortalidad aleatoria de los tallos pequeños.
Los análisis fueron realizados usando funciones de estadística espacial del software de programación y estadística R. El paquete que se usó fue spatstat versión 1.12-1 (Baddeley & Turner 2005) y las funciones que se corrieron fueron Kest, Kcross, envelope y rthin.
Resultados
Estructura y dinámica poblacional. De las nueve especies estudiadas Otoba lehmanii fue la más representativa en términos de área basal (80.606 m2). Esta misma especie fue también la más representativa en área basal para toda la parcela con un 11.12% (ab total parcela = 724.826 m2). Le siguen en orden decreciente Matisia bolivarii (3.37%), Billia rosea (2.38%), Myrcia fallax (1.84%), Couepia platycalyx (1.10%), Eschweilera caudiculata (1.08%), Naucleopsis naga (0.98%), Licania veneralensis (0.79%) y Carapa guianensis (0.63%). En cuanto a abundancia relativa, Otoba lehmanii también fue la más abundante de las nueve especies, pero en toda la parcela ocupó el doceavo lugar con un 1.5% de representatividad de un total de 133 110 individuos censados en 25 ha.
En términos dinámicos, la tasa de mortalidad promedio anual para la parcela fue 3.65%, ligeramente por encima de la tasa de reclutamiento que fue 3.52%. De las nueve especies, la que presentó la mayor tasa de mortalidad fue B. rosea (2.26%); tres tuvieron tasas entre 1% y 1.66% y el resto tuvieron tasas menores al 1%. Visto por categorías de tamaño (Tabla 2), tanto el número de individuos como el área basal para cada clase se conserva entre un censo y otro, aunque en la mayoría de las especies el número de tallos pequeños es proporcionalmente mayor que el número de árboles grandes, en una razón que va desde 2:1 hasta 8:1. Las únicas especies que tuvieron una cantidad similar de tallos pequeños y grandes fueron O. lehmanii (911 vs. 1082) y M. bolivarii (984 vs. 727).
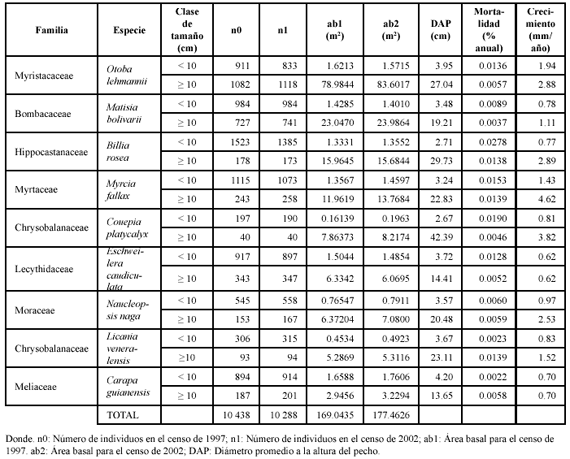
En cuanto a la tasa de mortalidad, en cinco especies ésta fue mayor en la categoría de los árboles pequeños, en dos fue muy similar (M. fallax y N. naga), y en otras dos (L. veneralensis y C. guianensis) fue mayor en los árboles grandes que en los pequeños. Con respecto a la tasa de crecimiento, en casi todas las especies fue mayor en los árboles grandes que en los pequeños. Solo dos especies tuvieron una tasa de crecimiento igual para las dos clases de tamaño consideradas en el estudio (E. caudiculata y C. guianensis).
Patrones de distribución espacial. Los patrones de distribución espacial indican una desviación significativa de la esperada de Poisson para la mayoría de las especies, con tendencia al agrupamiento. La única especie que presentó una desviación significativa hacia un patrón de distribución uniforme fue O. lehmanii entre 1 y 6 m de distancia (Figura 1a). B. rosea y C. guianensis mostraron una distribución agrupada independiente de la distancia (Figura 1b y 1c). L. veneralensis, M. bolivarii, N. naga y E. caudiculata presentaron desviaciones hacia el agrupamiento por encima de 5, 6, 7 y 7.8 m, respectivamente (Figuras 1d-1g), y C. platycalyx y M. fallax tuvieron distribuciones agrupadas en rangos de distancia entre 2.5 y 83 m y entre 1 y 87 m, respectivamente (Figuras 1h y 1i).

Patrones de distribución espacial por clases de tamaño. Los tallos pequeños de la mayoría de las especies tuvieron una tendencia igual o similar a la de la población completa en cuanto a la escala de agrupamiento, mientras que los grandes marcaron la diferencia con respecto al patrón general, pero principalmente a escalas pequeñas. Esto se debe en gran parte al alto número de tallos pequeños con respecto al número de tallos grandes.
En el caso de O. lehmanii se aprecia que los tallos pequeños están distribuidos de manera aleatoria a pequeñas escalas (0-10 m) y agrupados el resto de las distancias, mientras que los grandes, a la misma escala, mostraron una distribución uniforme y solo por encima de 20 m tuvieron una distribución agrupada (Figura 2). Esto indica que la tendencia a la uniformidad a escalas pequeñas que se observó cuando se tuvo en cuenta toda la población, es un efecto de la distribución de los árboles grandes y no de los pequeños.
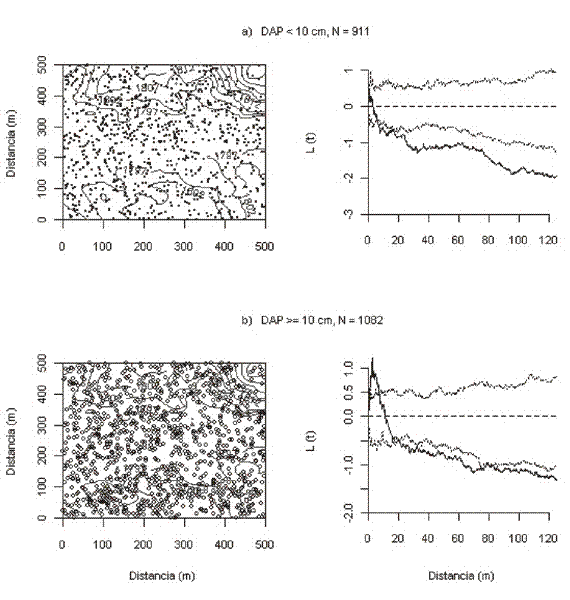
Para B. rosea y C. guianensis el patrón de agrupamiento de los tallos pequeños fue igual al de toda la población, es decir, que se mantuvo a cualquier escala, pero el de los grandes fue diferente; en B. rosea el patrón observado fue aleatorio a escalas pequeñas (entre 0-17 m) y se fue haciendo cada vez más agrupado con el incremento de t, y en C. guianensis el patrón fue en su mayoría aleatorio pero tuvo una desviación significativa hacia un patrón uniforme entre 5-6 m y entre 8-9 m de distancia.
Las demás especies también presentaron patrones agrupados para los tallos pequeños similares a los de la población completa, con una tendencia a la aleatoriedad a distancias menores a 10 m y más agrupada a medida que t incrementaba. Sin embargo, en el caso de los árboles grandes tuvieron patrones distintos. Algunas especies, como E. caudiculata, C. platycalyx y N. naga, tuvieron un patrón aleatorio a cualquier escala, otras como M. bolivarii mostraron una desviación significativa hacia un patrón uniforme entre 2-3 m y agrupado por encima de 9 m, y otras como L. veneralesis y M. fallax presentaron desviaciones hacia un patrón agrupado pero en rangos de distancia específicos.
Para el censo del 2002, donde además de registrar los tallos muertos se incluyeron los nuevos, los patrones de distribución de los árboles grandes y pequeños se mantuvieron igual que en el censo de 1997 en todas las especies. Cuando se analizó el patrón de distribución espacial de los árboles grandes que sobrevivieron, bajo la hipótesis de la mortalidad aleatoria de los tallos pequeños, se encontraron desviaciones significativas de los intervalos de confianza únicamente para las especies que tuvieron un patrón uniforme a escalas pequeñas (Tabla 3).
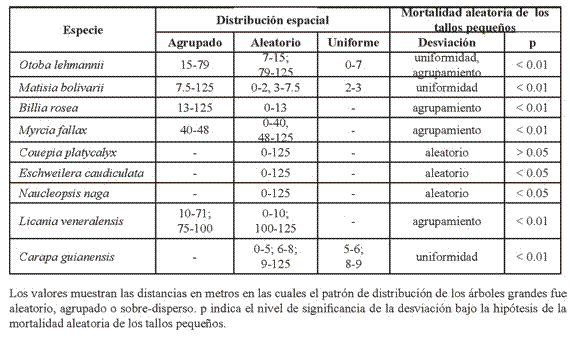
Así, para O. lehamnii la desviación de los intervalos de confianza indica que la tendencia hacia la regularidad es altamente significativa entre 0-12 m (Figura 3a), para M. bolivarii entre 0-5 m (Figura 3b) y para C. guianensis entre 5-6 m y 8-9 m de distancia (Figura 3c), lo que significa que la tendencia a la sobre-dispersión en las escalas señaladas no es explicada por la mortalidad aleatoria de los tallos pequeños.
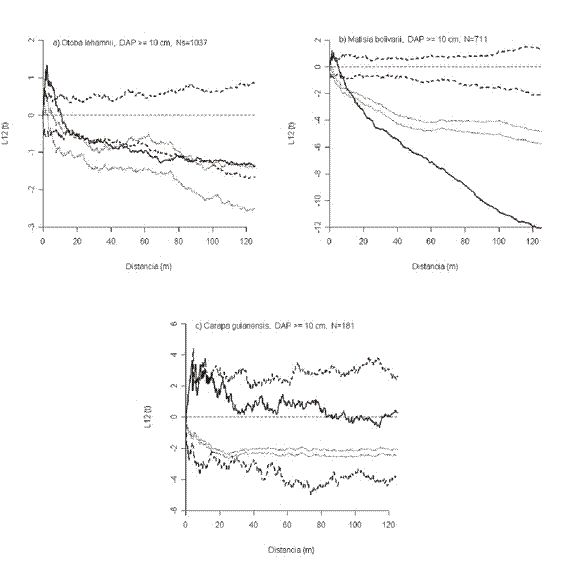
Interacción entre árboles grandes y pequeños. La Tabla 4 resume las interacciones que se presentaron entre los árboles grandes y los pequeños de las especies estudiadas.
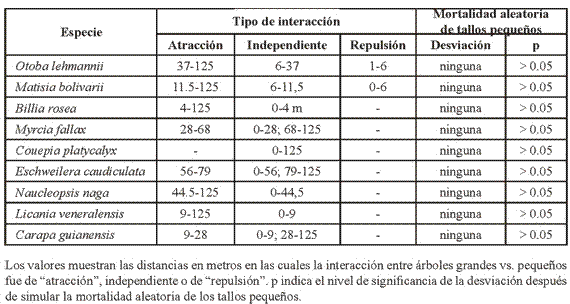
Las interacciones fueron en su mayoría de "atracción". B. rosea tuvo una interacción de "atracción" por encima de 4 m, L. veneralensis por encima de 9 m y N. naga por encima de 44.5 m. C. guianensis tuvo una interacción de "atracción" únicamente entre 9-28 m, E. caudiculata entre 56-79 m y M. fallax entre 28-68 m. Por su parte, O. lehmanii y M. bolivarii mostraron una interacción de "atracción" por encima de 37 m y 11.5 m, respectivamente y de "repulsión" a distancias menores a 6 m en ambos casos. La única especie que no mostró ningún tipo de interacción fue C. platycalyx.
Distribución espacial de los individuos muertos. Cinco de las nueve especies tuvieron algún grado de agrupamiento de los individuos muertos. B. rosea por encima de 2 m de distancia (Figura 4a), C. platycalyx desde 0 hasta 77 m (Figura 4b), M. fallax entre 60-65 m y entre 76-101 m (Figura 4c), M. bolivarii entre 1-5 m, 10-16 m, 24-34 m y por encima de 39 m (Figura 4d); y O. lehmanii entre 2-12 m (Figura 4e).
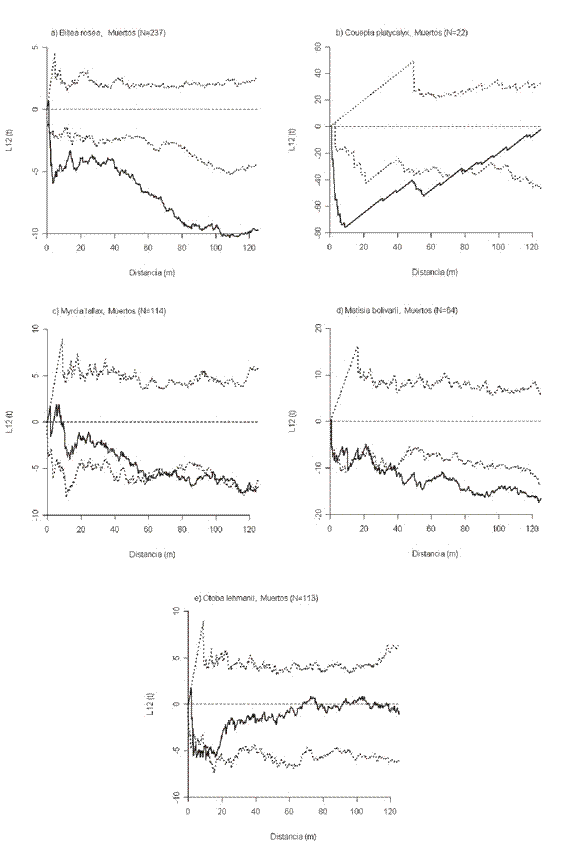
Interacción entre árboles vivos y muertos. Ocho de las nueve especies presentaron algún grado de interacción de "atracción". Para B. rosea ocurrió en todo el rango de distancias analizado, mientras que para M. bolivarii empezó a partir de 6.5 m. En el caso de C. guianensis se dio únicamente entre 6 y 43 m, y para C. platycalyx se presentó entre 11.5-15 m y entre 26-77 m. E. caudiculata tuvo una interacción de "atracción" entre 17-74.5 m y L. veneralensis a distancias mayores a 62 m. Para M. fallax la "atracción" se presentó en tres rangos de distancia (2.5-5 m, 9-66 m y 98-106 m), mientras que N. naga fue la que tuvo mayores interacciones de "atracción" pese a que se dieron de manera intermitente a lo largo de todo el rango de distancias analizado. Finalmente, para O. lehmanii la interacción de los dos eventos comparados no tuvo desviaciones significativas de la esperada de Poisson.
Los intervalos de confianza obtenidos a partir del modelo de mortalidad aleatoria indican que la interacción entre árboles vivos y muertos para todas las especies no presenta desviaciones significativas de lo esperado bajo dicho modelo para un rango total de 125 m, es decir, que la interacción de "atracción" entre los individuos vivos y muertos que se presentó en la mayoría de las especies no tiene nada que ver con los procesos de mortalidad de los individuos.
DISCUSIÓN
En general, los resultados indican que sí existen diferencias en los patrones de distribución de los tallos pequeños y los grandes para las nueve especies de árboles estudiadas. Sin embargo, estas diferencias no pueden ser atribuidas exclusivamente a la mortalidad selectiva de los tallos pequeños, es decir, a la mortalidad ocasionada por agentes bióticos o abióticos asociados a factores denso-dependientes como los planteados por Janzen y Connell. En efecto, la predicción inicial que suponía un patrón tendiente a la uniformidad de los árboles grandes debido a la mortalidad selectiva de los tallos pequeños, se cumplió sólo para dos de las nueve especies estudiadas y a escalas pequeñas, que es donde ocurren la mayoría de las interacciones intra e inter específicas en organismos sésiles como las plantas.
El caso más evidente fue el de O. lehmanii, la cual tuvo una marcada y significativa desviación hacia un patrón uniforme en la distribución de los árboles grandes a pequeña escala, lo que sugiere el efecto de agentes diferentes al azar como los causantes de la mortalidad de los tallos más pequeños. Apoyan este indicio, el hecho de que la distribución de los individuos que murieron entre el primero y segundo censo tuvieron un patrón agrupado a la misma escala en que los árboles grandes presentaron una distribución uniforme. No obstante, la ausencia de interacción entre los individuos vivos y muertos indica que en principio la mortalidad agrupada no puede explicar en su totalidad el patrón de distribución regular de los árboles grandes sobrevivientes. Por otra parte, la interacción de "repulsión" que se presentó en la misma escala entre los árboles grandes y los pequeños, no tuvo desviaciones significativas bajo la hipótesis de la mortalidad aleatoria, lo que apunta a que el distanciamiento entre árboles grandes y pequeños tampoco puede ser explicado en su totalidad por la mortalidad agrupada de los tallos pequeños.
El segundo caso que apoya las predicciones iniciales fue el patrón encontrado para M. bolivarii, que fue similar al de O. lehmanii pero en un rango de distancia más reducido. También para esta especie se presentó una desviación significativa del patrón observado de los árboles grandes sobrevivientes con respecto a los intervalos generados a partir de la hipótesis de la mortalidad aleatoria de los tallos pequeños; pero a diferencia de O. lehmanii, el patrón agrupado de los individuos que murieron fue muy débil y además no se registró ningún tipo de interacción repulsiva entre los individuos vivos vs. muertos que pudiera explicar el patrón regular de los árboles grandes a causa de la mortalidad agrupada de los tallos pequeños.
A pesar de las similitudes encontradas en los patrones de distribución de estas dos especies, no es posible afirmar que los procesos que llevaron a la distribución regular de los árboles grandes a escalas pequeñas sean los mismos, pero sí se pueden hacer algunas inferencias a partir de los datos de dinámica poblacional y del conocimiento de sus historias de vida. Por un lado, el patrón agrupado observado en los tallos pequeños de la mayoría de las especies estudiadas, atribuido al tamaño y/o peso de los frutos que caen por gravedad debajo o muy cerca del árbol "madre", sugiere que un alto porcentaje de semillas germina a manera de parches bajo la copa de un árbol adulto. Además, se sabe que depredadores y dispersores de frutos y semillas grandes (Dasyprocta sp. y Agouti sp.), tienden a consumirlas parcial o totalmente en el mismo sitio en el que son encontradas, o son transportadas a distancias muy cortas (50 m), donde suelen enterrarlas cuando no alcanzan a consumirlas en una sola jornada de forrajeo (e.g. Dasyprocta punctata) (Smythe 1982). De ser así, no es raro encontrar un patrón espacial agrupado de semillas y plántulas de dichas especies, a menos que la oferta de frutos no sea muy alta o que exista un agente dispersor bastante efectivo como para alejar las semillas lo suficiente de su árbol "madre". Observaciones de campo en la parcela mostraron que es frecuente encontrar semillas de especies como el zapote (M. bolivarii) y el cuángare (O. lehmanii) bajo adultos de la misma especie; también, a menudo se observaron frutos de zapote mordidos por guatín (Dasyprocta sp.), pero siempre muy cerca del árbol adulto cargado con frutos. Bajo este supuesto y teniendo en cuenta las predicciones hechas bajo el modelo de Janzen y Connell, las semillas y las plántulas más agrupadas o menos distanciadas entre sí, serían más susceptibles de ser atacadas por patógenos o herbívoros, lo que derivaría en un distanciamiento progresivo entre los individuos que sobreviven.
Los patrones de distribución espacial encontrados para O. lehmanii y M. bolivarii apoyan parcialmente dichos planteamientos si consideramos que, a pequeñas escalas, los tallos pequeños tuvieron una distribución aleatoria; es decir, que probablemente la mortalidad de las semillas o de las plántulas provocaron un distanciamiento que se reflejó en un patrón menos agrupado de los tallos pequeños de lo que se esperaría si todas las semillas o plántulas se hubiesen desarrollado con éxito.
Por otra parte, existen cualidades inherentes a cada una de las especies que sugieren la ocurrencia de procesos diferentes. O. lehmanii, por ejemplo, es una especie bastante común, ampliamente distribuida y con una notable oferta de frutos en la parcela. De acuerdo con información disponible de otra especie relacionada (O. parviflora, Pringle et al. 2007), se presume que un alto porcentaje de las semillas de esta especie germina de manera exitosa, lo que indica que probablemente la mortalidad no ocurre drásticamente a este nivel, sino en estadios posteriores como plántulas y/o juveniles; así que es factible que las semillas germinen en altas densidades cerca de los adultos o a distancias muy cortas entre sí.
De otro lado, al observar los patrones de distribución de los tallos pequeños separados de los grandes, fue evidente una tendencia hacia un patrón uniforme causada por los árboles grandes, principalmente aquellos con DAP entre 30 y 40 cm. Dado que la uniformidad fue más evidente en esta categoría, se presume que la tendencia hacia la sobre-dispersión en una clase de tamaño específica, es el producto de un proceso de mortalidad gradual de los tallos más pequeños por requerimientos de luz. Esto podría sustentarse con los hallazgos de Pringle y sus colegas (2007) en otra especie de Otoba a la que catalogaron como una especie intolerante a la sombra. Dichas evidencias, sumadas al hecho de que el análisis bivarido entre tallos pequeños y grandes mostró una clara "repulsión" a la misma escala espacial en que los árboles grandes tuvieron un patrón de distribución uniforme, indican que los tallos pequeños están más separados de sus padres potenciales que lo esperado si su distribución fuera independiente; es decir, que no están creciendo con éxito cerca de un adulto por falta de luz.
Con base en lo anterior, se propone una estructura espacial derivada de un proceso como el que se describe a continuación: 1- frutos que caen por su tamaño y peso bajo el árbol "madre" formando parches; 2- semillas que germinan y permanecen como plántulas por un largo período de tiempo; 3- plántulas que crecen paulatinamente y se convierten en tallos pequeños (DAP < 10 cm) que también se distribuyen de manera agrupada pero van muriendo gradualmente en la medida en que los requerimientos de luz se hacen más necesarios para su desarrollo. La mortalidad ocurre de manera agrupada y afecta a aquellos que están más cerca del árbol "madre"; por ende, sobreviven los que están más lejos o por fuera del área de la copa del árbol parental ("hipótesis de la distancia"); 4- tallos pequeños que logran desarrollarse y alcanzar diámetros por encima de los 10 cm y hasta unos 40 cm aproximadamente, comenzando a exhibir un patrón espacial uniforme a escalas pequeñas, la cual se acentúa cuando alcanzan un tamaño entre los 30-40 cm de DAP. Aunque también se presentan casos de mortalidad, ya no es tan marcada como en las clases diamétricas más pequeñas; lo que sí ocurre es un desarrollo notable de los individuos, ya que en la categoría diamétrica entre 10-20 cm fue donde se registró la tasa promedio de crecimiento más alta de la especie (3.23 mm/año). Es probable que al sobrepasar este tamaño, aumenten las probabilidades de sobrevivencia, lo cual explicaría la mayor abundancia de árboles grandes con respecto a la de los pequeños; 5- por último, los individuos que se convirtieron en árboles grandes muestran un patrón de distribución agrupado a manera de parches localizados en distintos sitios de la parcela, apreciables dependiendo de la escala de observación.
Un proceso muy distinto se puede inferir para M. bolivarii, cuyos frutos tienen una corteza relativamente fácil de romper, raspar o roer por los mamíferos que los consumen (Dasyprocta sp. y Agouti sp.), y según Pringle et al. 2007, quienes también incluyeron en su estudio tres especies del género Matisia, las plántulas y los juveniles son más tolerantes a la sombra comparados con los de O. lehmanii, y son susceptibles de ser atacados por patógenos en la medida en que la proporción de juveniles/adulto es mayor. Estas características explican hasta cierto punto el patrón aleatorio de los tallos pequeños a distancias muy cortas, provocado por la mortalidad de semillas o plántulas, y la desviación hacia un patrón uniforme de los árboles grandes. Esto también indica que a pesar de que el patrón predominante en las dos clases de tamaño analizadas es el agrupado, algunos procesos dependientes de la distancia o la densidad están ocurriendo a pequeñas escalas y serían los causantes de las desviaciones hacia patrones uniformes en las clases de diámetro más grandes.
Adicionalmente, la "repulsión" observada entre tallos pequeños y grandes soportaría el supuesto de que, a distancias muy cortas, hay un efecto de mortalidad (probablemente de plántulas) denso-dependiente, que va ampliando la distancia entre individuos y por tanto genera una distribución aleatoria de los tallos pequeños y un patrón posterior tendiente a la uniformidad de las clases diamétricas mayores. No obstante, al no presentarse una desviación significativa bajo la hipótesis de la mortalidad aleatoria de los tallos pequeños, no es posible atribuir el distanciamiento entre los dos grupos de tamaño a un proceso de mortalidad selectiva de las clases diamétricas más pequeñas. Se presume por tanto, que para M. bolivarii podría estar operando uno de los modelos propuestos por Janzen y Connell, ya sea como un efecto de denso-dependencia o debido a la distancia y/o la densidad a escalas pequeñas. La mortalidad estaría provocada por agentes patógenos o enfermedades que atacan principalmente las semillas, o por herbívoros que atacan las plántulas.
Los resultados encontrados en las demás especies estudiadas no ofrecen indicios significativos que rechacen la predicción inicial de este estudio, ya que no mostraron una tendencia hacia un patrón regular de los árboles grandes sobrevivientes, o simplemente las desviaciones fueron muy débiles y no tuvieron soporte en los análisis realizados para los individuos muertos, o en las interacciones entre árboles grandes vs. pequeños, o entre vivos vs. muertos. Por el contrario, algunos de los patrones observados sugieren una tendencia a ser "indiferentes" respecto a los procesos de mortalidad que ocurren al interior de sus poblaciones. Así, encontramos especies como B. rosea, que pese a compartir varias características similares con O. lehmanii en cuanto a sus frutos y semillas (tamaño, textura de la corteza, dureza), tuvo patrones de distribución que no sugieren la ocurrencia de interacciones intra-específicas que afecten la distribución de los árboles sobrevivientes. En especies como C. platycalyx y L. veneralensis se infiere que los patrones encontrados para las clases de tamaño analizadas pueden deberse a que la oferta de frutos y semillas de estas especies no es muy alta, a los procesos extremadamente lentos de germinación de sus semillas (Fosters 1986, Vargas 2002), y a la baja tasa de mortalidad que se registró en cada una de ellas. Además, el patrón de distribución aleatorio de los pocos individuos que murieron sugiere otros factores como los causantes de la mortalidad en estas especies, tales como la caída de ramas y/o epífitas, característica del interior de bosques nublados como el de La Planada.
De acuerdo con la discusión planteada en relación a los procesos que pudieron haber ocasionado los actuales patrones de distribución espacial de las especies estudiadas, se puede decir que el aporte a la dinámica temporal y espacial de este tipo de bosque no es apreciable en las especies de estratos más altos, es decir, dosel y sub-dosel, considerando que las especies incluidas en el presente trabajo representan el 15% de las especies de dosel que conforman la parcela (~ 60 spp). Por un lado, porque las tasas de mortalidad y reclutamiento fueron muy bajas y no tuvieron mayor efecto sobre el número total de individuos, y por otro, porque fueron muy pocos los casos en los que se presentaron desviaciones hacia un patrón de distribución uniforme en los árboles grandes apoyadas por procesos de mortalidad agrupada de los tallos pequeños (O. lehmanii y M. bolivarii). Esto significa que procesos más dinámicos (con mayores tasas de mortalidad) probablemente ocurren a nivel de estadios más tempranos como plántulas o semillas, o en especies de estrato arbustivo que son las que tuvieron las tasas de mortalidad más altas en toda la parcela (> 5 % promedio anual) (Samper & Vallejo 2007). Se presume entonces que la dinámica espacial de las especies de sotobosque puede jugar un papel importante en la determinación de la distribución espacial de toda la comunidad.
Por otra parte, el bajo nivel de interacciones de "repulsión" que se presentó entre los eventos analizados, sugiere que procesos como la competencia por recursos tal vez no sea un factor relevante entre individuos de la misma especie, como lo puede ser entre individuos de especies diferentes. En las especies donde se encontró cierto nivel de "repulsión" entre árboles grandes y pequeños, se sugiere que la luz y el ataque de patógenos o herbívoros determinan en gran medida el patrón espacial de categorías de tamaño específicas (e.g. Otoba lehmanii y Matisia bolivarii).
Al comparar los resultados de este estudio con los encontrados en otras parcelas en bosques tropicales de zonas bajas (Condit et al. 2000), se puede deducir que los bosques pre-montanos presentan tendencias similares en cuanto al patrón de distribución espacial de los individuos cuando se observa toda la población; es decir, que el patrón fue agrupado en casi todas las escalas, aunque el análisis de Condit et al. (2000) se centró principalmente en los patrones a escalas pequeñas. Pero, al separarla por clases de tamaño, se pudo apreciar que el patrón de distribución fue diferente en los árboles mayores a 10 cm de DAP. Mientras que la mayoría de las especies analizadas por Condit y sus colaboradores tuvieron patrones agrupados a escalas pequeñas (0-10 m) para las clases diamétricas mayores a 10 cm, en la Planada los individuos de esta misma categoría tuvieron una distribución aleatoria en el mismo rango de distancia e incluso a mayor escala; es decir, que aunque sí exhibieron tendencia al agrupamiento, ésta ocurrió a distancias mayores a 10 m y en algunos casos en rangos de distancia específicos que podrían estar asociados con sus estrategias de dispersión. Cabe aclarar que Condit et al. (2000) usaron un índice de agrupamiento diferente y afirman que su método permite aislar clases de distancia específicas para determinar un patrón, mientras que el estadístico K(t) de Ripley confunde efectos a grandes distancias con efectos a cortas distancias ya que es un índice de distribución acumulada. No obstante, sería interesante comparar los hallazgos encontrados para las especies de este estudio (ocho de dosel y una de sub-dosel) con las demás especies pertenecientes a los diferentes estratos del bosque de La Planada, con el fin de saber si se asemejan o no a los patrones encontrados por Condit et al. (2000), quienes no encontraron diferencias en los patrones de distribución (agrupada) entre especies de dosel y especies de sotobosque en la mayoría de las parcelas incluidas en el estudio que realizaron, excepto en Barro Colorado, en Panamá.
A la luz de la teoría neutral de la biodiversidad y la biogeografía, se puede afirmar que los hallazgos de este estudio apoyan los planteamientos de Hubbell en el sentido en que apuntan hacia una combinación de diferentes estrategias que desarrollan los individuos dentro de la población, que se traducen en mayor abundancia cuando se es susceptible de alta mortalidad en estadios tempranos (e.g. M. fallax, N. naga), o una mayor resistencia a los agentes de mortalidad pero pagando el precio de una población reducida (e.g. C. platycalyx, L. veneralensis).
En la carrera evolutiva de las especies por ocupar un lugar en espacios donde los recursos son aparentemente reducidos al interior de una comunidad, tienen lugar muchos procesos que apoyan de manera no excluyente diversas hipótesis. Por ejemplo, especies de plantas tróficamente similares como las incluidas aquí, indican que su permanencia a nivel de la comunidad ha sido el producto de distintos procesos donde el azar juega un papel importante. En este sentido, así como la interacción entre los individuos de la misma especie fue evidente a pequeñas escalas, lo cual se traduce en un mayor o menor distanciamiento relacionado con la edad y asociado a eventos de denso-dependencia, existen también evidencias sobre interacciones inter-específicas fundamentadas en la competencia por los recursos, la mayoría de ellos también a pequeñas escalas (Kenkel 1988, Duncan 1991, Haase et al. 1996). En consecuencia, el arreglo espacial de una comunidad sería la suma de los arreglos espaciales de muchas especies que se han establecido a través de procesos distintos como la competencia, la especialización de hábitat (teoría de nichos), los procesos denso-dependientes ocasionados por patógenos o por herbívoros, e inclusive la plasticidad genética que en términos evolutivos y ecológicos les han permitido ser indiferentes a procesos estocásticos. Probablemente la combinación de todos estos eventos es lo que permite, en tiempo evolutivo, la coexistencia de un gran número de especies en áreas reducidas, en donde las estrategias de supervivencia y desarrollo van a determinar su grado de dominancia (densidad), que no necesariamente se relaciona con su capacidad competitiva.
AGRADECIMIENTOS
Al Instituto "Alexander von Humboldt" por permitir el uso de los datos de la parcela. A la Fundación FES y a la Reserva Natural La Planada, fundamentales para el acceso y protección del área de estudio. El establecimiento de la parcela y el primer censo fue financiado por el Instituto "Alexander von Humboldt" y el segundo por la Fundación Mellon a través del Instituto Smithsoniano de Investigaciones Tropicales. Muchos colaboradores de campo y de laboratorio han participado en el estudio de esta parcela, pero queremos agradecer especialmente a las personas de las comunidades de San Isidro y del Resguardo Indígena A'wa por su esfuerzo y dedicación durante el trabajo de campo. Agradecemos a R. Condit, E. Losos, S. Loo de Lao y P. Hall, del Centro para las Ciencias Forestales del Trópico del Instituto Smithsoniano, por su apoyo y orientación durante el manejo del programa R, y a A. Baddeley de la University of Western Australia y a J. Mateu de la Universitat Jaume I (Castellón, España), por su colaboración con el manejo del paquete de estadística utilizado. Los profesores O. Vargas, A. Duque, A. Bonilla y A. Rudas de la Universidad Nacional de Colombia, y J.I. Barrera de la Pontificia Universidad Javeriana, hicieron valiosos aportes en diferentes fases de desarrollo de la investigación, A. Ramírez y A. Repizo de la Pontificia Universidad Javeriana apoyaron de manera importante temas específicos del proyecto. Gracias también al profesor K. Harms de la University of Louisiana y a N. Norden de la University of Connecticut, y a dos evaluadores anónimos, por la revisión crítica de una versión anterior del manuscrito. Este trabajo hizo parte de la tesis de Maestría en Ciencias-Biología (Línea Ecología) de la Universidad Nacional de Colombia, Sede Bogotá, de la primera autora.
LITERATURA CITADA
1. ARMESTO, J.J, J.D. MITCHELL & C. VILLAGRAN.1986. A comparison of spatial patterns of trees in some tropical and temperate forests. Biotropica 18(1): 1-11. [ Links ]
2. ASHTON, P.S. 1969. Speciation among tropical forest trees: some deductions in the light of recent evidence. Biological Journal of the Linnean Society 1: 155-196. [ Links ]
3. BADDELEY, A. & R. TURNER. 2005. Spatsat: An R package for analyzing spatial point patterns. Journal of Statistical Software 12 (6): 1-42. [ Links ]
4. BROKAW, N. & R. T. BUSING. 2000. Niche versus chance and tree diversity in forest gaps. Trends in Ecology and Evolution 15:183-188. [ Links ]
5. CONDIT, R. 1998. Tropical forest census plots: Methods and results from BCI, Panamá and a comparison with other plots. Springer-Verlag, Berlín. [ Links ]
6. CONDIT, R., P.S. ASHTON, P. BAKER, S. BUNYAVEJCHEWIN, S. GUNATILLEKE, N. GUNATILLEKE, S.P. HUBBELL, R.B. FOSTER, A. ITOH, J.V. LAFRANKIE, H.S. LEE, E. LOSOS, N. MANOKARAN, R. SUKUMAR & T. YAMAKURA. 2000. Spatial patterns in the distribution of tropical tree species. Science 288: 1414-1418. [ Links ]
7. CONNELL, J.H. 1971. On the role of the natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. Págs. 298-310 en: P.J. den Boer & G.R. Gradwell (eds), Dynamics of Populations. Proceeding of the Advanced Study Institute, Centre for Agricultural Publishing and Documentation, Wageningeng. [ Links ]
8. DIXON, P.M. 2002. Ripley's K function. Págs. 1796-1803 en: A.H. El-Shaarawi & W.W. Piegorsch (eds), Encyclopedia of Environmetrics. John Wiley & Sons, Ltd, Chichester. [ Links ]
9. DUNCAN, R.P. 1991. Competition and the coexistence of species in a mixed podocarp stand. Journal of Ecology 79: 1073-1084. [ Links ]
10. FOSTERS, S.A. 1986. On the adaptative value of large seeds for tropical moist forest trees: a review and synthesis. Botanical Review 52: 260-299. [ Links ]
11. GENTRY, A.H.1995. Patterns of diversity and floristic composition in neotropical montane forests. Págs. 103-126. en: S.P. Churchill, H. Balslev, E. Forero & J. Lutein (eds), Biodiversity and Conservation of Neotropical Montane Forests. New York Botanical Garden, Nueva York. [ Links ]
12. HAASE, P., F.L. PUGNAIRE, S.C. CLARK & L.D. INCOLL. 1996. Spatial pattern in two-tired semi-arid shrub land in southeastern Spain. Journal of Vegetation Science 7: 527-534. [ Links ]
13. HOLDRIDGE, L. R. 1982. Ecología basada en zonas de vida. Instituto Interamericano de Cooperación para la Agricultura, IICA, San José, Costa Rica. (Serie Libros y Materiales Educativos; no. 34). [ Links ]
14. HUBBELL, S.P. 1979. Tree dispersion, abundance, and diversity in a tropical dry forest. Science 203: 1299-1309. [ Links ]
15. HUBBELL, S.P. 1980. Seed predation and the coexistence of tree species in tropical forests. Oikos 35: 214-229. [ Links ]
16. HUBBELL, S.P. 2001. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton University Press, Princeton. [ Links ]
17. HUBBELL, S. P. & R. B. FOSTER.1983. Diversity of canopy trees in a neotropical forest and implications for conservation. Págs. 25-41 en: S.L. Sutton, T.C. Whitmore & A.C. Chadwick (eds), Tropical Rain Forest: Ecology and Management. Blackwell Scientific Publications, Oxford. [ Links ]
18. JANZEN, D.H. 1970. Herbivores and the number of tree species in tropical forests. The American Naturalist 104: 501-528. [ Links ]
19. KENKEL, N.C. 1988. Patterns of self-thinning in Jack-pine: testing the random mortality hypothesis. Ecology 69(4): 1017-1024. [ Links ]
20. KING, A.W. & S.L. PIMM.1983. Complexity, diversity, and stability: A reconciliation of theoretical and empirical results. The American Naturalist 122(2): 229-239. [ Links ]
21. LAWES, M.J. & J.A.F. OBIRI. 2003. Canopy gaps in subtropical forest in South Africa: size of the species pool and not the number of available niches limits species richness. Journal of Tropical Ecology 19:594-556. [ Links ]
22. LIEBERMAN, M. & D. LIEBERMAN.1994. Patterns of density and dispersion of forest trees. Págs.106-119 en: L.A. McDade, H.A. Henspenheide, G.S. Hartshorn (eds), La Selva: Ecology and Natural History of a Neotropical Rain Forest. University of Chicago Press, Chicago. [ Links ]
23. OLIVER, C.D. & B.C. LARSON.1996. Forest stands dynamics. Wiley and Sons. Nueva York. [ Links ]
24. PETERSON, C.J. & E.R. SQUIERS.1995. An unexpected change in spatial pattern across 10 years in an aspen-white-pine forest. Journal of Ecology 83: 847-855. [ Links ]
25. PRINGLE, E.G., P.ÁLVAREZ-LOAYZA & J. TERBORGH. 2007. Seed characteristics and susceptibility to pathogen attack in tree seeds of Peruvian Amazon. Plant Ecology 193: 211-222. [ Links ]
26. SAMPER, C. & M. I. VALLEJO. 2007. Estructura y dinámica de poblaciones de plantas en un bosque andino. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 31(118): 57-68. [ Links ]
27. SMYTHE, N. 1982. Relaciones entre las épocas de abundancia de frutos y los métodos de dispersión de las semillas en un bosque neotropical. Págs 77-83 en: G. De Alba, & R.W. Rubinoff (eds), Evolución en los Trópicos. Smithsonian Tropical Researh Institute, Editorial Universitaria, Panamá. [ Links ]
28. STERNER, R.W., C.A. RIBIC & G.E. SHATZ. 1986. Testing for life historical changes in spatial patterns of four tropical tree species. Journal of Ecology 74: 621-633. [ Links ]
29. VALLEJO, M.I., C. SAMPER, H. MENDOZA & J.T. OTERO. 2004. La Planada Forest Dynamics Plot, Colombia. Págs. 517-526 en: E. Losos & E.G. Leigh, Jr. (eds), Tropical Forest Diversity and Dynamism: Findings from a large scale plot network. University of Chicago Press, Chicago. [ Links ]
30. VALLEJO, M.I., A.C. LONDOÑO, R. LÓPEZ, G. GALEANO, E. ÁLVAREZ& W. DEVIA. 2005. Establecimiento de Parcelas Permanentes en Bosques de Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá D.C. (Serie: Métodos para Estudios Ecológicos a Largo Plazo). [ Links ]
31. VARGAS, W.G. 2002. Guía ilustrada de las plantas de las montañas del Quindío y los Andes Centrales. Universidad de Caldas (ed). Colección Ciencias Agropecuarias, Manizales. [ Links ]
32. WARD, J.S., G.R. PARKER & F.J. FERRANDINO. 1996. Long-term spatial dynamics in an old-growth deciduous forest. Forest Ecology and Management 83: 189-202. [ Links ]
Recibido: 17/10/2008
Aceptado: 27/01/2009

