










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.32 no.1 Bogotá Jan./June 2010
Floristic diversity and structure of Andean forests remnants near to Los Nevados National Park, Central Colombian Andes
MARCELA ALVEAR
JULIO BETANCUR
PILAR FRANCO-ROSSELLI+
Department of Botany, California Academy of Sciences, 55 Music Concourse Drive, San Francisco, California 94118, U.S.A. malvear@calacademy.org
Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Apartado 7495, Bogotá D.C. Colombia. jcbetancurb@unal.edu.co
RESUMEN
Se caracterizó la diversidad, la composición florística y la estructura de los bosques de la Reserva Torre Cuatro, la cual hace parte de la zona amortiguadora del Parque Nacional Natural Los Nevados, Cordillera Central de los Andes colombianos. Los remanentes de bosque se subdividieron en tres zonas de acuerdo a la altitud: baja (2650-2900 m), media (3000-3300 m) y alta (3400-3750 m), en cada una de las cuales se muestreó 0.1 ha y se censaron todos los individuos con DAP = 2.5 cm. La riqueza de especies encontrada para las tres zonas (62 para la baja, 69 para la media y 48 para la baja) fue superior al promedio registrado para otros bosques andinos situados a altitudes similares. En todas las zonas la mayor proporción de individuos tuvieron alturas inferiores a 7.2 m. La composición florística y la fisonomía de los bosques muestra que los de las zonas baja y media corresponden a los descritos para la franja andina propiamente dicha, mientras que los de la alta a la franja alto andina. En todas las zonas las familias con mayor importancia ecológica fueron Melastomataceae y Asteraceae, pero también se destacaron Cunoniaceae en la baja, Betulaceae en la media y Ericaceae en la alta. En todas las zonas unas pocas especies sobresalieron por su mayor importancia ecológica en la comunidad (IVI); en las zonas baja y media las cinco especies con más IVI acumularon aproximadamente el 30% del mismo, mientras que en la zona alta cerca del 47%. Las especies con más IVI fueron Weinmannia pubescens (32.7) en la zona baja, Alnus acuminata (29.9) en la media y Tibouchina andreana (33.4) en la alta.
Palabras clave. Andes, Colombia, Bosque montano, Bosque nublado, Diversidad florística.
ABSTRACT
The diversity, structure, and floristic composition of the Andean forests remnants at Torre Cuatro Reserve were characterized. This reserve is part of the buffer zone of Los Nevados National Park, Central Cordillera of the Andes in Colombia. The forest remnants were grouped into three zones according to the altitude: low (between 2650 and 2900 m), medium (between 3000 and 3300 m), and high (between 3400 and 3750 m). In each zone, 0.1 ha was sampled and all individuals with DBH = 2.5 cm were surveyed. The species richness found in the zones (62 in low, 69 in medium, 48 in high) was higher compared to that found in other forests with similar altitudes. In all zones the largest proportion of individuals had heights less than 7.2 m. Floristic composition and physiognomy of the zones shows that the forest of low and middle zones correspond to those described as the characteristic Andean forest, while the high zone corresponds to the high Andean forest. In all areas the most ecologically important families were Melastomataceae and Asteraceae, but some other families were also important, such as Cunoniaceae in the low zone, Betulaceae in the middle and Ericaceae in the high zone. In all altitudes a few species were most ecologically significant in the community (IVI); in the medium and low zones only five species accumulated approximately 30% of the IVI, while in the high zone five species accumulated 47%. The species with the highest IVI were Weinmannia pubescens (32.7) in the low zone, Alnus acuminata (29.9) in the medium zone and Tibouchina andreana (33.4) in the high zone.
Key words. Andes, Cloud forest, Colombia, Floristic diversity, Montane forest.
Recibido: 26/01/2009
Aceptado: 17/09/2009
INTRODUCCIÓN
Colombia es uno de los países con mayor biodiversidad a nivel mundial, buena parte de la cual está concentrada en los biomas de montaña (Gentry 1991 a, b, 1995, van der Hammen & Rangel-Ch. 1997). La gran riqueza de la flora y de la vegetación de la zona andina colombiana y de las altas montañas tropicales es el resultado de múltiples fenómenos ocurridos en el pasado, los que contribuyeron a conformar un escenario con alta heterogeneidad ambiental y, por ende, con alta diversidad regional (van der Hammen 1995, Gentry 1982).
Los Andes tropicales han sido considerados como uno de los principales centros de diversidad y especiación en el mundo, además de que concentran gran cantidad de endemismos (Churchill et al. 1995, Brown & Kappelle 2001, Hamilton 2001, Kappelle & Brown 2001, Ulloa & Jørgensen 2005). Así pues, los Andes se convierten en un área crítica para la conservación a nivel mundial, pues se estima que el 50% de las especies de plantas presentes en esta región no se encuentran en ningún otro lugar del planeta (Mittermeier et al. 1999, Rodríguez-Mahecha et al. 2004).
Por otra parte, la región andina en Colombia ha sido el soporte del desarrollo económico y cultural del país, lo que ha generado en ella un gran incremento demográfico, acompañado de una evidente falta de planificación y destrucción de sus ecosistemas naturales. Se aprecia una acelerada expansión de la frontera agrícola y ganadera con los consecuentes procesos de agotamiento del recurso hídrico, empobrecimiento de los suelos, aumento de la erosión y desaparición de muchos elementos de la biota (Hernández-Camacho 1990, Castaño-Uribe 1991, Cavelier 1991, Kattan & Alvarez-López 1995, Vis 1995, van der Hammen & Rangel-Ch. 1997). Se estima que en Colombia perduran menos del 10% de los bosques andinos originales y menos del 5% de los bosques altoandinos, encontrándose principalmente restringidos a fragmentos de diferentes tamaños y grados de aislamiento (Henderson et al. 1991, Carrizosa-U. 1990, Cavelier et al. 2001).
En esta contribución se presenta la caracterización florística y fisionómica de los remanentes de bosque andino de una reserva natural que hace parte de la zona de amortiguación del Parque Nacional Natural Los Nevados (PNNN), región que ha sido reconocida como uno de los centros con mayor diversidad de plantas en América (Davis et al. 1997).
ÁREA DE ESTUDIO
El estudio se realizó en la Reserva Torre Cuatro, localizada en la vertiente occidental de la Cordillera Central colombiana, aproximadamente a 5º01' Norte y 75º22' Oeste. Está ubicada entre las veredas La Esperanza y Maltería, a 19 km del municipio de Manizales (departamento de Caldas) y hace parte de la zona de amortiguación del PNNN. Esta reserva fue adquirida por la Corporación Autónoma Regional de Caldas con el fin de reforestar y proteger la cuenca alta del río Chinchiná, la cual surte de agua al acueducto de la ciudad de Manizales y es fuente de generación eléctrica en la región. El predio posee una extensión aproximada de 310 ha, tiene forma irregular y alargada y posee altitudes entre 2650 y 3750 m. Tiene temperatura media anual de 9 ºC y precipitación promedio anual entre 1600 y 1800 mm, presentando dos períodos de lluvias (marzo-mayo y septiembre-noviembre) y dos períodos secos (diciembre-febrero y junio-agosto). Los suelos son derivados de cenizas volcánicas, son superficiales a moderadamente profundos, bien drenados, ligeramente ácidos, con textura gruesa y contenido medio de materia orgánica. Por otra parte, presenta un relieve fuertemente ondulado y con altas pendientes (70-80% en promedio), con evidencias de erosión hídrica, deslizamientos y escurrimientos difusos, principalmente en las áreas de pendiente más fuerte, caminos y sitios donde originalmente el ganado abrevaba (Vélez-Palacio 1996, Anónimo 1999, 2004, Restrepo 1999).
Estos predios estaban sometidos a la explotación agrícola intensiva desde finales del siglo XIX, especialmente a la ganadería lechera y al cultivo de papa y forrajes. Actualmente, la vegetación es un mosaico de pastizales y remanentes de bosque secundario en diferentes estados de regeneración natural, los cuales están restringidos principalmente a los bordes de los cursos de agua y a las zonas con mayores pendientes. Estos remanentes de bosque han sido objeto de perturbación a través del tiempo, especialmente por la entresaca selectiva de maderas, pero presentan un dosel arbóreo conformado por especies que son elementos florísticos característicos de los bosques maduros de la región (Vélez-Palacio 1996, Restrepo 1999, Anónimo 1999, 2004). La región corresponde a la formación de selva andina, en el sentido de Cuatrecasas (1958), o al bosque muy húmedo montano bajo (bmh-MB) y transición al montano (bmh-M), según el sistema de zonas de vida de Holdridge (Espinal 1977).
MÉTODOS
Muestreo. El muestreo se realizó entre abril de 1999 y febrero de 2000. El área de estudio fue dividida en tres zonas con base en el análisis de fotografías aéreas, la altitud y el reconocimiento de campo, así: la zona baja (entre 2600 y 2900 m), la zona media (entre 3000 y 3300 m) y la zona alta (entre 3400 m y 3750 m). En todos los casos, el muestreo se realizó en áreas boscosas que no tuvieran impacto humano muy reciente y que representaran la vegetación arbórea del lugar.
Se utilizó el método de "transectos de 0.1 ha de Gentry" (Phillips & Miller 2002), para lo que en cada una de las zonas se realizaron diez transectos de 50 x 2 m (100 m2). Los transectos se dirigieron al azar dentro de los parches de bosque natural, evitando las áreas alteradas -como caminos y claros dentro del bosque- y que se traslaparan. En cada transecto se censaron todos los individuos con diámetro a la altura del pecho (DAP) > 2.5 cm, medido 1.3 m por encima del suelo. A cada individuo se le registró la altura (m), el perímetro a la altura del pecho (CAP) y el hábito o forma de crecimiento, en las categorías de altura de árbol ( > 10 m), arbolito (< 10 m), arbusto erecto, arbusto escandente, hierba y liana, de acuerdo con los criterios de Font-Quer (1982) y Moreno (1984). En el caso de individuos con varios vástagos, brotes basales o ramificaciones por debajo de 1.3 m, se midió independientemente el perímetro a cada uno de ellos. Las familias de angiospermas se organizaron de acuerdo al sistema de Cronquist (1981) y los pteridofitos de acuerdo al de Tryon & Tryon (Kramer & Green 1990). Los ejemplares testigo fueron depositados en los herbarios COL, FAUC, HUA y MO.
Tratamiento de los datos. El perímetro medido a cada individuo (CAP) se transformó a DAP mediante la ecuación DAP = CAP/π. En el caso de las plantas ramifi cadas por debajo de la altura del pecho, o con varios vástagos o brotes basales, el DAP total se calculó según la ecuación DAPt = (4 At/π)1/2, en donde At (área total) = ΣAi, y Ai (área individual) = πDAP1/2 (Franco- Rosselli et al. 1997). Posteriormente, los DAP se transformaron a área basal mediante la ecuación AB = π/4(DAP)2 (Mueller- Dombois & Ellenberg 1974).
Para cada especie y en cada zona de muestreo se calculó la densidad (DeR), la frecuencia (FR) y la dominancia (DoR) relativas, cuya sumatoria representa el índice de valor de importancia (IVI) (Finol 1976), así: DeR = (# individuos de la especie/# total de individuos) x 100; FR = # transectos en los que aparece la especie/Σfrecuencia de todas las especies) x 100; DoR = (ΣAB de los individuos de la especie/ΣAB del total individuos) x 100. Para analizar la distribución de cada uno de los parámetros ecológicos en la comunidad se construyeron histogramas de frecuencia estableciendo intervalos, según la ecuación C = (Xmax-Xmin)/m, donde C = amplitud del intervalo, Xmax = valor máximo, Xmin = valor mínimo, m = 1 + 3.3 (log N) y N = # total de individuos (Rangel-Ch. & Velásquez 1997).
Se calculó el índice de importancia de las familias (VIF) en cada zona como la sumatoria de la densidad, la dominancia y la riqueza relativas de cada familia (Mori & Boom 1983). Para estimar el número de especies esperadas de acuerdo con la altitud se utilizó la ecuación No. especies = 260.1 - (0.073) altitud (Gentry 1995). Para comparar la similitud florística cualitativa entre las zonas de estudio se calcularon los índices de similitud de Jaccard y Sørensen, teniendo en cuenta la presencia/ausencia de las especies (Magurran 1989).
RESULTADOS
Riqueza y composición florística. La mayor riqueza de especies se presentó en la zona media (69), seguida de la baja (61) y la alta (48). Sin embargo la zona alta presentó el mayor promedio de individuos por especie, siendo éste muy similar en las zonas baja y media. Por otra parte, el número de especies esperadas en la zona baja estuvo cercano al encontrado (60 vs. 61), mientras que estuvo muy por debajo de lo encontrado en las zonas media y alta (Tabla 1).

Las familias Asteraceae, Melastomataceae y Solanaceae fueron las que presentaron mayor número de géneros y especies en todas las zonas, especialmente Asteraceae que siempre ocupó el primer lugar en todas. No obstante, Lauraceae y Ericaceae también se destacaron por su riqueza en las zonas media y alta, respectivamente (Tabla 2).
Más de las tres cuartas partes de las familias estuvieron representadas en las tres zonas sólo por una o dos especies (92%, 81% y 76 % en la baja, media y alta, respectivamente), mientras que ningún género estuvo representado por más de tres especies, siendo estos Cestrum en la baja, Cestrum, Cyathea, Meliosma y Palicourea en la media y Gaultheria, Oreopanax y Weinmannia en la alta (Anexo 1, Anexo 2, Anexo 3).
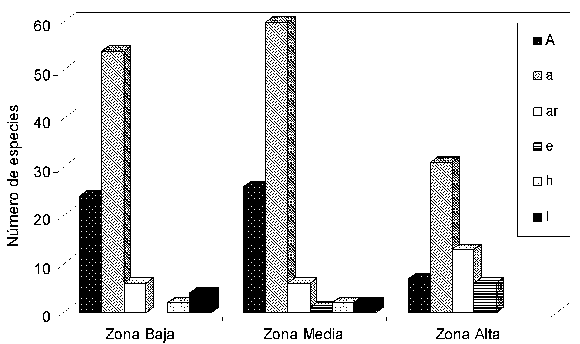
Formas de crecimiento. En las tres zonas estudiadas el mayor número de especies correspondió a las que crecían como arbolitos. Por otra parte, la proporción de especies de árboles y arbolitos fue mayor en las zonas baja y media, mientras que la de arbustos fue mayor en la alta. Los arbustos escandentes se presentaron sólo en las zonas media y alta, siendo mayor el número de especies en esta última. Así mismo, las hierbas y las lianas estuvieron ausentes en la zona alta (Figura 1).
Estructura. En todas las zonas la mayor proporción de individuos se presentó en los primeros intervalos de altura, hasta alcanzar 7.2 m, punto a partir del cual hay una disminución gradual en el número de individuos, la cual es más drástica en la zona alta. Por otra parte, según la distribución de alturas de los individuos se podrían considerar tres estratos para los bosques de las zonas baja y media, encontrándose el inferior hasta 2.6 m de altura, el intermedio hasta 7.2 m y el superior hasta 16.4 m (Figura 2).
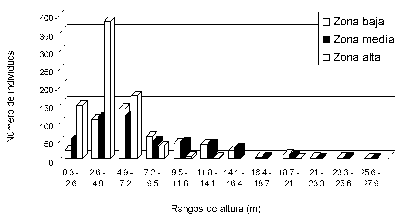
Figura 2. Distribución vertical de los individuos con DAP > 2.5 cm en los remanentes de bosque de la reserva Torre Cuatro (Manizales, Colombia).
La zona alta presentó bosques de menor porte y también con tres estratos: el inferior hasta 2.6 m de altura, el intermedio hasta 4.9 m (el cual presenta el mayor número de individuos) y el superior hasta 9.5 m. A partir de este punto se encuentran algunos individuos emergentes que pueden alcanzar hasta 14 m de altura y que corresponden a Axinaea colombiana, Escallonia myrtilloides, Hesperomeles ferruginea, Oreopanax discolor y Tibouchina aff. lepidota. Sólo en uno de los transectos se presentaron dos individuos de Escallonia myrtilloides con alturas cercanas a 20 m (Figura 2, Anexo 3).
Densidad. Los bosques de la zona alta presentaron el mayor número de individuos (759), mientras que éste valor fue muy similar en las zonas media y baja (470 y 480, respectivamente; Tabla 1). En general, en las tres zonas la mayoría de los individuos pertenecen a unas pocas especies, siendo Weinmannia pubescens y Leandra subseriata las especies con más individuos en la zonas baja y media, mientras que en la alta fueron Tibouchina andreana, Baccharis latifolia y Gaultheria foliolosa (Anexo 1, Anexo 2, Anexo 3).
Frecuencia. En todas las zonas la mayoría de las especies presentaron valores bajos de frecuencia. Sin embargo, los bosques de la zona alta presentaron las especies más frecuentes, pues Tibouchina andreana, Gaultheria foliolosa, Baccharis latifolia y Escallonia myrtilloides se encontraron en los diez transectos realizados. Por el contrario, las especies más frecuentes en las zonas baja y media se presentaron sólo en seis de los diez transectos realizados (Anexo 1, Anexo 2, Anexo 3).
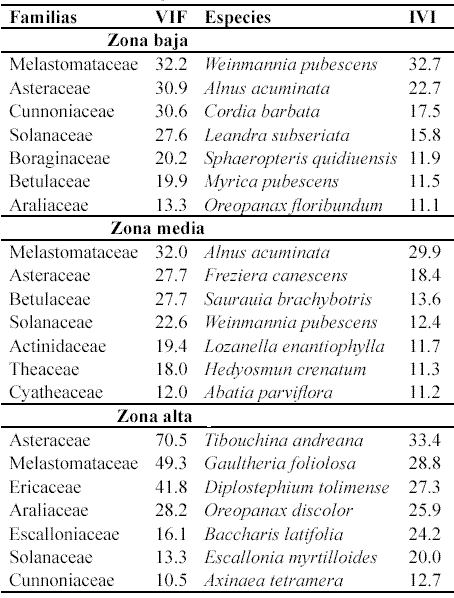
Dominancia. El mayor valor acumulado de área basal se presentó en los bosques de la zona media (9.09 m2), seguido de los de las zonas alta (8.23 m2) y baja (5.51 m2). En las tres zonas la mayoría de las especies presentaron valores bajos de dominancia relativa, mientras que sólo unas pocas sobresalieron por la alta concentración de la misma. Así, Weinmannia pubescens, Alnus acuminata y Cordia barbata acumularon el 42% de dominancia en la zona baja, Alnus acuminata y Freziera canescens cerca del 36% en la zona media y Diplostephium tolimense, Oreopanax discolor, Gaultheria foliolosa y Tibouchina andreana más del 50% en la zona alta (Anexo 1, Anexo 2, Anexo 3).
Índice de Valor de Importancia (IVI). La mayoría de las especies en las tres zonas presentaron índices de valor de importancia bajos (Anexo 1, Anexo 2, Anexo 3). Sin embargo, en todas las zonas hay unas pocas especies que sobresalen por su mayor importancia ecológica en la comunidad y que logran acumular una alta proporción del IVI. Así, en las zonas baja y media las cinco especies con más IVI acumulan aproximadamente el 30% del mismo, mientras que en la zona alta acumulan cerca del 47%. Las especies con más IVI acumulado fueron Weinmannia pubescens (32.7) en la zona baja, Alnus acuminata (29.9) en la media y Tibouchina andreana (33.4) en la alta.
Por otra parte, los bosques de las zonas baja y media compartieron dos especies dentro de las cuatro con más importancia ecológica en la comunidad (Weinmannia pubescens y Alnus acuminata), mientras que la zona alta no compartió ninguna con las anteriores (Tabla 3).
Además, los índices de Jaccard y Sørensen también muestran que la mayor similaridad se presentó entre las zonas baja y media, con 0.36 y 0.8 respectivamente (Tabla 4).
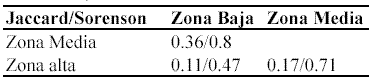
Valor de importancia de familias (VIF). En las tres zonas la mayoría de las familias presentaron índices de importancia bajos (Anexo 1, Anexo 2, Anexo 3). Sin embargo, en todas las zonas hay unas pocas familias que sobresalen por su mayor importancia ecológica en la comunidad y que acumulan una alta proporción del VIF, especialmente en la zona alta. Así, en las zonas baja y media las primeras cinco familias con más VIF acumularon entre el 42% y 47% del mismo, mientras que en la zona alta acumulan cerca del 70% (Tabla 3).
En las tres zonas Melastomataceae y Asteraceae son las dos familias con mayores valores de VIF, aunque en distinto orden de importancia. Por otra parte, es de resaltar que en los bosques de las zonas baja y media el tercer lugar de predominio ecológico estuvo ocupado por familias que estuvieron representadas por sólo una especie: Cunoniaceae con Weinmannia pubescens en la baja y Betulaceae con Alnus acuminata en la media (Tabla 3).
DISCUSIÓN
Aspectos florísticos y estructurales. Teniendo en cuenta la composición florística de los bosques andinos registrada por Cuatrecasas (1958) y la encontrada en el PNNN (Cleef et al. 1983, Rangel-Ch. 1995c, Rangel-Ch. et al. 1995), se puede afirmar que en la Reserva Torre Cuatro están presentes aún los elementos típicos de la flora nativa de la región, a pesar de la fuerte presión humana a la que ha estado sometida durante el último siglo. No obstante, es importante señalar que algunos de estos elementos, como en el caso de especies arbóreas maderables, se encuentran representados por pocos individuos (Anexo 1, Anexo 2, Anexo 3) y que generalmente están aislados en medio de los potreros.
Los resultados referidos a la fisonomía, la estratificación, las formas de crecimiento, la composición florística, el predominio ecológico y la similaridad señalan que los remanentes de bosque natural de la Reserva Torre Cuatro se agrupan en dos tipos principales: el bosque de la franja andina propiamente dicha, correspondiente a los remantes de las zonas baja y media entre 2600 y 3400 m de altitud, y el bosque de la franja alto-andina, correspondiente a los remanentes de la zona alta entre 3400 y 3750 m de altitud. Diversos autores han diferenciado a la selva o bosque alto-andino de la selva o bosque andino propiamente dicho, en el sentido que originalmente lo consideraba Cuatrecasas (1958), para referirse a los bosques ubicados en las partes más altas de las montañas. Así, el bosque alto-andino se caracteriza por tener menor riqueza de especies, por el reemplazo de algunos de los grupos florísticos dominantes y por algunos cambios fisonómicos en la vegetación, como son la disminución en el porte de los árboles, la reducción de la superficie foliar y el incremento del indumento en diferentes órganos de las plantas, entres otros (Mora-O. & Sturm 1994, Rangel-Ch. 1995b, van der Hammen 1998, Gradstein et al. 1995, Salamanca 1988).
El cambio entre los bosques andino y alto-andino se efectúa muy gradualmente y se ha relacionado principalmente con las diferencias en la temperatura ambiental, la humedad relativa, la precipitación y la exposición a la radiación solar (Mora-O. & Sturm 1994, Rangel-Ch. 1995b, van der Hammen 1998, Gradstein et al. 1995, Salamanca 1988). A su vez, en Colombia los límites altitudinales para estos dos tipos de bosque presentan algunas variaciones locales y entre las vertientes de las cordilleras. Para las cordilleras Central y Occidental se ha propuesto que la franja de bosque alto-andino se ubica entre 3300 y 3800 (-3900) m de altitud, mientras que para la Sierra Nevada de Santa Marta y la Cordillera Oriental entre 3000 y 3400 (-3500) m de altitud (Cleef & Rangel-Ch. 1984, Rangel-Ch., 1995b, Salamanca 1988). Los resultados obtenidos en este estudio para los remanentes de bosque de la reserva Torre Cuatro concuerdan con los límites altitudinales propuestos para la Cordillera Central y especialmente en lo referente al bosque alto-andino en el macizo volcánico Ruiz-Tolima, en donde los parches de este tipo de bosque alcanzan las mayores altitudes y no hay una franja de subpáramo definida (Cleef et al. 1983, Salamanca 1991, Salamanca et al. 2003).
Las familias Melastomataceae y Asteraceae, que correspondieron a las más importantes en las tres zonas de estudio, también han sido detectadas dentro de las más importantes en los bosques andinos del Ecuador (Valencia & Jørgensen 1992). Algunas familias de plantas en la reserva Torre Cuatro, aunque están representadas en todas las zonas de estudio, tienen mayor riqueza en alguna de ellas, como ocurre con Asteraceae y Ericaceae en la zona alta. También, se encontró que algunas especies están restringidas a sólo una de las zonas estudiadas, lo que corresponde a una segregación y reemplazo florístico que ya ha sido registrado para la región andina en general (Cuatrecasas 1958, Rangel-Ch. 1995a, Bach et al. 2007). Algunos de los principales grupos de plantas registrados para la zona alta de la Reserva Torre Cuatro, pero no registrados para las zonas media y baja, fueron las familias Berberidaceae, Escalloniaceae y Grossulariaceae, la asterácea Diplostephium tolimense, la clusiácea Hypericum laricifolium, las ericáceas Gaultheria anostomosans, G. erecta, G. foliosa y Macleania rupestris, las melastomatáceas Miconia salicifolia y Tibouchina andreana, la mirsinácea Myrsine dependens, la rosácea Hesperomeles ferruginea y varias especies de solanáceas de los géneros Saracha y Sessea (Anexo 1, Anexo 2, Anexo 3).
Por otra parte, en la zona alta no se encontró una franja definida de subpáramo, mientras que algunos elementos típicos de éste aparecen mezclados con los propios del bosque, como en el caso de Hypericum laricifolium. Esta es una especie caracteríctica de subpáramo que se registró en el bosque alto-andino de la reserva Torre Cuatro (Anexo 3) y, también, ha sido encontrada por otros autores en los parches de bosque alto-andino del macizo volcánico Ruíz-Tolima entre 3600 y 3900 m de altitud (Cleef et al. 1983, Salamanca 1991).
Algunas de las especies registradas como las más importantes para las zonas andina y alto-andina en el PNNN (Cleef et al. 1983, 2003) también presentaron alto predominio ecológico en la Reserva Torre Cuatro, como Weinmannia pubescens y Diplostephium tolimense. Por otra parte, otras especies registradas como importantes en el PNNN no lo fueron en la Reserva Torre Cuatro, como Hesperomeles ferruginea, H. obtusifolia var. microphylla, Miconia salicifolia y Brunellia goudotii. Otras especies, registradas simplemente como asociadas a algunas comunidades vegetales en el PNNN, presentaron alto predominio ecológico en Torre Cuatro, como Gaiadendron punctatum y Oreopanax discolor. Así mismo, especies como Tibouchina andreana y Gaultheria foliolosa son bastante importantes en Torre Cuatro y, sin embargo, no habían sido registradas como tal para ningún otro bosque alto-andino (Tabla 3, Anexo 1, Anexo 2, Anexo 3).
Es importante señalar la ausencia del roble (Quercus humboldtii) en los bosques de la Reserva Torre Cuatro, así como el alto predominio de otras especies como Weinmannia pubescens o Alnus acuminata, por ejemplo (Tabla 3). Algunos autores (Cleef et al. 1983, Rangel-Ch. & Franco-Rosselli 1985) han relacionado la ausencia del roble en los bosques nativos con una mayor humedad y la presencia de capas humíferas más gruesas. De hecho, van der Hammen et al. (1995) atribuyeron la ausencia del roble en la vertiente occidental del PNNN, la misma en la que están los bosques estudiados, a su mayor humedad ambiental debida a la baja radiación solar que recibe (Witte 1995).
Así mismo, los trabajos florísticos realizados en el PNNN (Cleef et al. 1983, Salamanca 1991, Rangel-Ch. et al. 1995) no han registrado la presencia del aliso (Alnus acuminata), especie que ocupó el primer o segundo lugar de predominio ecológico en los bosques andinos de la reserva Torre Cuatro (Tabla 3). Según Rangel-Ch. (com. pers.) el aliso no se presenta en poblaciones naturales dentro de la vegetación zonal del PNNN, como si ocurre en los bosques andinos del Quindío y del nevado del Tolima, o en áreas inundables de la sabana de Bogotá (van der Hammen 1998). Creemos que la abundancia de A. acuminata en la Reserva Torre Cuatro, así como en otros bosques cercanos a la ciudad de Manizales, podría deberse a su dispersión a partir de plantaciones cercanas que fueron establecidas desde los años cincuenta del siglo veinte, a raíz del auge de la reforestación con esa especie (Pérez-Arbeláez 1996, Cavelier 1993, Murcia 1997). Sin emabargo, se sabe que en inmediaciones de la Reserva Torre Cuatro nunca existieron plantaciones de esta especie. La rápida expansión y colonización de esta especie se ve favorecida por su dispersión a través del viento y por su rápida germinación y crecimiento (Carlson & Dawson, en Murcia 1997). Igualmente, la regeneración del aliso se ve favorecida en áreas con pendientes pronunciadas (hasta del 80%), suelos pedregosos y con alta pluviosidad y nubosidad (Suárez-Navarro et al. 1984), condiciones que se presentan en algunos parches de bosque de las zonas baja y media de la Reserva Torre Cuatro, especialmente en sitios cercanos a donde ocurrieron deslizamientos de tierra.
Al igual que para otros bosques montanos colombianos, tal como se mostró atrás, la estructura y composición florística conocida para otros bosques andinos ecuatorianos es diferente, como se ha registrado para Pasochoa, un bosque húmedo montano del Ecuador situado a 3260-3310 m de altitud (Valencia & Jørgensen 1992). Aunque utilizando métodos diferentes (muestreo de 1 ha de individuos con dbh > 5 cm), en Pasochoa la especie ecológicamente más importante -Miconia theaezans- tiene un alta dominancia en la comunidad (IVI de 85), valor que supera por mucho a la primera especie con más IVI en cualquiera de las tres zonas estudiadas en Torre Cuatro (Tabla 3). Por otra parte, entre Pasochoa y Torre Cuatro hay muchas otras diferencias florísticas y estructurales, como las especies con más importancia ecológica y los géneros con más especies, pero las familias Melastomataceae y Asteraceae se destacan entre las más importantes en ambas localidades.
Teniendo en cuenta el número de individuos y especies, se encontró que la mayor diversidad florística se concentró en las clases de menor altura (figuras 1, 2), lo que corresponde con lo encontrado por algunos autores para otros bosques andinos (Jørgensen et al. 1995, Marín-C. & Betancur 1997, Galindo-T. et al. 2003). Por otra parte, el número de individuos con DAP > 2.5 cm en 0.1 ha aumentó con relación a la altitud de los bosques (Tabla 1), condición que ha sido registrada como uno de los principales cambios fisonómicos de la vegetación relacionados con el incremento en la altitud (Cavelier 1991).
Respecto a las formas de crecimiento también se observaron diferencias estructurales entre los bosques de las diferentes zonas muestreadas. Los arbustos estuvieron pobremente representados en las zonas baja y media, forma de crecimiento que presentó mayor importancia en la alta. Por otra parte, la disminución en el número de especies arbóreas en la zona alta se esperaba, teniendo en cuenta que la fisonomía típica de la vegetación alto-andina se caracteriza por la disminución en el porte de los árboles (Cuatrecasas 1958, Rangel-Ch. 1995b, Jørgensen et al. 1995). Las hierbas solamente estuvieron representadas en las zonas baja y media por especies del género Chusquea, las cuales adquieren gran importancia en las zonas de borde de bosque, así como en la regeneración y recuperación de la vegetación después de los derrumbes y deslizamientos de tierra (Cleef et al. 1983, Gentry 1991 a, b).
Riqueza de especies. La zona alta presentó el menor número de especies (Tabla 1), lo cual concuerda con el gradiente de disminución de especies respecto al aumento en la altitud (Gentry 1995). Por otra parte, el hecho de que la zona media presentara más especies que la zona baja, contrario a lo esperado, puede deberse a que la zona baja tiene mayor grado de intervención humana, debido a que hace parte de la zona de transito por la que los campesinos conducen sus productos, es atravesada por la carretera principal que comunica a Manizales con Bogotá y sus bosques han estado sometidos a la entresaca y la tala desde tiempo atrás. Además, los parches de bosque presentes en la zona media están más aislados entre sí que en la zona baja, lo que acentúa el efecto de heterogeneidad ambiental y, posiblemente, el incremento en la riqueza de especies. Algunos autores afirman que los disturbios en intensidades intermedias pueden aumentar la riqueza al disminuir la dominancia de una o pocas especies, con lo que se liberan recursos para otras menos competitivas, y/o incrementan la heterogeneidad ambiental (Denslow 1985, White & Pickett 1985, Tilman & Pacala 1993). De hecho, Vis (1995) encontró que en algunas áreas del PNNN con intensidades moderadas de pastoreo la riqueza de especies aumentaba, debido principalmente a la mayor heterogeneidad ambiental. Sin embargo, si los disturbios son muy intensos aumenta la proporción de especies exóticas y disminuye la diversidad (Denslow 1985, Kessler 2001, Hofstede 1995, Vis 1995). De la misma forma, se ha registrado que en bosques montanos de Bolivia con intensidades intermedias de disturbio el endemismo de cuatro grupos de plantas (Araceae, Bromeliaceae, Palmae y Pteridophyta) es mayor que en áreas de bosque maduro, y que disminuye en zonas fuertemente perturbadas (Kessler 2001).
Por otra parte, teniendo en cuenta la regresión propuesta por Gentry (1995) para bosques andinos por encima de 1500 m de altitud, el número de especies esperadas en las zonas media y alta fue bastante inferior al encontrado (Tabla 1). Una de las principales razones que explican esta diferencia es que cuando fue formulada la ecuación solo se disponía de datos de riqueza para bosques por debajo de 2900 m de altitud. Nosotros recalculamos la ecuación teniendo en cuenta datos nuevos provenientes de muestreos con métodos similares. En total usamos los datos provenientes de 21 localidades andinas de Gentry (1995), los de una localidad en el Santuario de Fauna y Flora de Iguaque (Boyacá, Colombia) de Marín-C. & Betancur (1997), los de cuatro localidades del Santuario de Fauna y Flora Guanentá Alto Río Fonce (Santander y Boyacá, Colombia) de Galindo-T. et al. (2003), y los de las tres localidades de la Reserva Torre Cuatro (Caldas, Colombia) que hacen parte de este estudio.
La nueva ecuación propuesta es: y (no. de especies) = 217.98 - (0.055 x altitud), en donde r2 = 0.77, n = 29 y p < 0.0001 (figura 3).
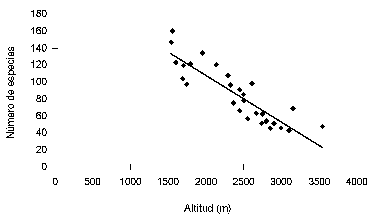
De todas formas, el número de especies esperadas obtenido a través de esta nueva ecuación es inferior al encontrado para cada una de las zonas de estudio. Además, el nuevo coeficiente de regresión obtenido es inferior al proporcionado originalmente por Gentry en 1995 (vs. r2 = 0,87). Lo anterior puede deberse a que el número de muestras para alturas superiores a 2900 m es aún muy reducido (Galindo-T. et al. 2003) y a que el modelo de regresión pueda ser diferente al lineal.
Tratando de buscar una mayor cercanía entre el número de especies obtenidas y las encontradas se realizaron otros análisis de regresión, utilizando sólo datos correspondientes a los muestreos por encima de 2370 m de altitud. Se obtuvo así más cercanía entre el número de especies observadas y esperadas, aunque el coeficiente de regresión disminuyó aún más (r2 < 0.2). De todas formas, estos resultados muestran la necesidad de explorar más este tipo de relaciones para describir su verdadero comportamiento, de realizar más muestreos a altitudes superiores a 2500 m y, además, señalan la importancia de relacionar los patrones de diversidad y riqueza de especies con otros parámetros distintos a la altitud, como los son la humedad, la precipitación y la temperatura, entre otros.
En la Tabla 5 se compara la riqueza de especies en las tres zonas estudiadas con la de otros bosques situados a altitudes similares y muestreados con métodos similares.
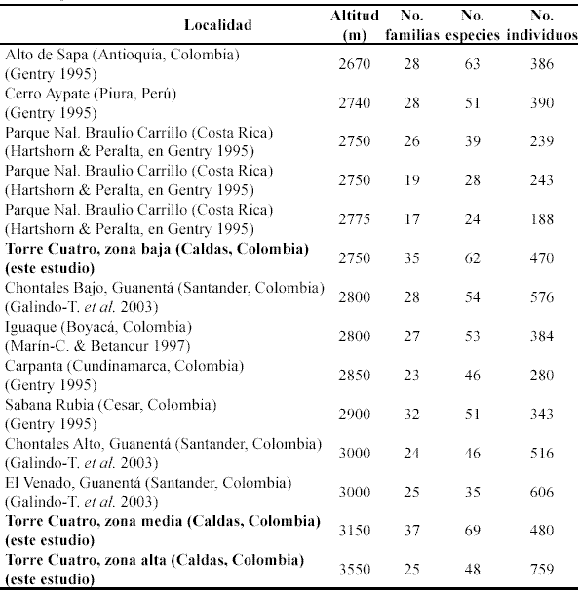
Se observa, como tendencia general, que en los bosques montanos situados entre 2620 y 3000 m la riqueza de especies disminuye a medida que aumenta la altitud. Sin embargo, las localidades centroamericanas (Parque Nacional Braulio Carrillo, Costa Rica, Gentry 2005) presentan valores inferiores comparados con las localidades colombianas, tanto en riqueza de especies como en número de individuos. Estos datos corresponden con la afirmación de que los bosques de montaña costarricenses, situados a alturas similares, son menos diversos que los de los Andes de la porción nor occidental de Suramérica (Gentry 1995, Galindo-T. et al. 2003).
Por otra parte, los bosques de las zonas media y alta de la reserva Torre Cuatro, a pesar de ser los que están a mayor altitud, presentan mayor riqueza de especies que los que les preceden con altitudes similares. La mayor riqueza presentada por los bosques de Torre Cuatro puede explicarse porque corresponden a un mosaico de parches de vegetación natural en diferentes estados de regeneración, producto de los disturbios ocasionados por la intervención humana, situación que incrementa la heterogeneidad ambiental y la diversidad. No obstante, también es de resaltar que esta mayor diversidad se relaciona con el hecho de que los bosques de Torre Cuatro no corresponden a bosques monoespecíficos, como ocurre en los robledales (Marín-C. & Betancur 1997). Así mismo, en relación con esta gran riqueza de los bosques de la reserva Torre Cuatro, se resalta el hecho de que algunas de las especies encontradas han sido registradas como endémicas para el centro de diversidad del PNNN (Davis et al. 1997), e incluso especies nuevas para la ciencia (Alvear 2000, Lozano-C. & Alvear 2001).
AGRADECIMIENTOS
Al Instituto de Ciencias Naturales y al Herbario Nacional Colombiano (COL), de la Universidad Nacional de Colombia, por las facilidades proporcionadas para el desarrollo del trabajo. A la Corporación Autónoma Regional de Caldas (CORPOCALDAS) por la financiación del trabajo de campo, especialmente a Mariela Londoño y Herbert Soto. A Diego Alvear, José D. García, Alexander Sabogal, Francisco Sánchez, Jairo y Melba, por su invaluable apoyo en el trabajo de campo. A Laura Clavijo, por sus sugerencias y ayuda en la edición final del manuscrito. A los botánicos que colaboraron con la determinación y/o confirmación de los especimenes: Ricardo Callejas (Piperaceae), Santiago Díaz (Asteraceae), José L. Fernández (varios grupos), Gustavo Lozano (Melastomataceae), José C. Murillo (Pteridophyta), Clara I. Orozco (Brunelliaceae, Solanaceae), Orlando Rivera (Araliaceae) y Charlotte M. Taylor (Rubiaceae). A los dos revisores anónimos por su concepto crítico sobre al manuscrito.
LITERATURA CITADA
1. ALVEAR, M. 2000. Flora y vegetación de la Reserva Torre Cuatro (Manizales, Colombia). Trabajo de grado. Departamento de Biología, Universidad Nacional de Colombia, Bogotá. [ Links ]
2. ANÓNIMO. 1999. Plan de ordenamiento ambiental del territorio de la cuenca del río Chinchiná. Síntesis del diagnóstico. Tomo I. Corpocaldas & Aguas de Manizales, Manizales. [ Links ]
3. ANÓNIMO. 2004. Acuerdo No. 16 (noviembre 4 de 2004) por el cual se declara Zona de Reserva Forestal Protectora el predio Torre Cuatro, ubicado en jurisdicción del municipio de Manizales, departamento de Caldas. Consejo Dircetivo, Corporación Autónoma Regional de Caldas (Corpocaldas), Manizales. [ Links ]
4. BACH K., M. KESSLER & S.R. GRADSTEIN. 2007. A simulation approach to determine statistical significance of species turnover peaks in a species-rich tropical cloud forest. Diversity and Distributions 13: 863-870. [ Links ]
5. BROWN, A.D. & M. KAPPELLE. 2001. Introducción a los bosques nublados del neotrópico: una síntesis regional. Págs. 25-40 en: M. Kappelle & A.D. Brown (eds.), Bosques nublados del neotrópico. Instituto Nacional de Biodiversidad (INBio), Santo Domingo de Heredia. [ Links ]
6. CARRIZOSA-U., J. 1990. La selva andina. Págs 151-184 en: J. Carrizosa-U. & J.I. Hernández-Camacho (eds.), Selva y Futuro. El Sello Editorial, Bogotá. [ Links ]
7. CASTAÑO-URIBE, C. 1991. Oasis de niebla. Págs 53-68 en: C. Uribe (ed.), Bosques de niebla de Colombia. Banco de Occidente, Cali. [ Links ]
8. CAVELIER, J. 1991. El ciclo del agua en bosques montanos. Págs. 69-83 en: C. Uribe (ed.), Bosques de niebla de Colombia. Banco de Occidente, Cali. [ Links ]
9. CAVELIER, J. 1993. Reforestation with the native tree Alnus acuminata: effects on phytodiversity and species richness in an upper montane rain forest area of Colombia. Págs. 78-85 en L. Hamilton & F. Scatena (eds.), Tropical montane cloud forest. East-West Center, UNESCO, International Institute of Tropical Forestry, U.S.A. [ Links ]
10. CAVELIER, J., D. LIZCAINO & M.T. PULIDO. 2001. Colombia. Págs. 443-496 en: M. Kappelle & A.D. Brown (eds.), Bosques nublados del neotrópico. Instituto Nacional de Biodiversidad (INBio), Santo Domingo de Heredia. [ Links ]
11. CHURCHILL, S. P., H. BALSLEV, E. FORERO & J.L. LUTEYN (Eds.). 1995. Biodiversity and conservation of Neotropical montane forests. The New York Botanical Garden, New York. [ Links ]
12. CLEEF, A. M., J.O. RANGEL-CH. & S. SALAMANCA. 1983. Reconocimiento de la vegetación de la parte alta del transecto Parque Nacional Natural Los Nevados. Págs. 150-173 en: T. van der Hammen, T., A. Pérez & P. Pinto (eds.), La Cordillera Central de Colombia, transecto Parque Nacional Natural Los Nevados. Estudios de Ecosistemas Tropandinos vol. 1. J. Cramer, Berlín. [ Links ]
13. CLEEF, A.M. & J.O. RANGEL-CH. 1984. Las selvas del transecto Buritaca. Págs. 267-406 en: T. van der Hammen & P. Ruiz-C. (eds.), La Sierra Nevada de Santa Marta (Colombia), transecto Buritaca-La Cumbre. Estudios de Ecosistemas Tropandinos vol. 2. J. Cramer, Berlín. [ Links ]
14. CLEEF, A.M., J.O. RANGEL-CH. & S. SALAMANCA. 2003. The Andean rain forests of the Parque Los Nevados, Codillera Central, Colombia. Págs. 79-142 en: van der Hammen, T., A. G. Dos Santos (eds.), La Cordillera Central Colombiana, transecto Parque Los Nevados. Estudios de Ecosistemas Tropandinos vol. 5. J. Cramer, Berlín. [ Links ]
15. CRONQUIST, A. 1981. An integrated system of classification of flowering plants. Columbia Universtity Press, New York. [ Links ]
16. CUATRECASAS, J. 1958. Aspectos de la vegetación natural en Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 10 (40): 221-268. [ Links ]
17. DAVIS, S.D., V.H. HEYWOOD, O. HERRERA-MACBRIDE, J. VILLA-LOBOS & A.C. HAMILTON (Eds.). 1997. Centres of plant diversity. A guide and strategy for their conservation. Vol. 3. The Americas. The World Wide Fund for Nature (WWF) & The World Conservation Union (IUCN). IUCN Publications Unit, Cambridge. [ Links ]
18. DENSLOW, J.S. 1985. Disturbance-mediated coexistence of species. Págs. 307-321 en: S. Pickett & P. White (eds.), The ecology of natural disturbance and patch dynamics. Academic Press, San Diego. [ Links ]
19. ESPINAL-T., L.S. 1977. Zonas de vida o formaciones vegetales de Colombia. Memoria explicativa sobre el mapa ecológico de Colombia. Instituto Geográfico Agustín Codazzi (IGAC), Bogotá. [ Links ]
20. FINOL, H. 1976. Estudio fitosociológico de las unidades 2 y 3 de la Reserva Forestal de Carapo, estado de Barinas. Acta Botánica Venezuelica 10 (1-4): 15-103. [ Links ]
21. FONT-QUER, P. 1982. Diccionario de botánica. Editorial Labor, Barcelona. [ Links ]
22. FRANCO-ROSSELLI, P., J. BETANCUR & J.L. FERNÁNDEZ-A. 1997. Diversidad florística en dos bosques subandinos del sur de Colombia. Caldasia 19 (1-2): 205-234. [ Links ]
23. GALINDO-T., R., J. BETANCUR & J.J. CADENA-M. 2003. Estructura y composición florística de cuatro bosques andinos del Santuario de Flora y Fauna Guanentá-Alto Río Fonce, Cordillera Oriental colombiana. Caldasia 25 (2): 313-335. [ Links ]
24. GENTRY, A. 1982. Neotropical floristic diversity: phytogeographical connections between Central and South America, pleistocene climatic fluctuation or an ancient of the Andean orogeny? Annals of the Missouri Botanical Garden 69 (3): 557-593. [ Links ]
25. GENTRY, A. 1991a. Vegetación del bosque de niebla. Págs 23-52 en: C. Uribe (ed.), Bosques de niebla de Colombia. Banco de Occidente, Cali. [ Links ]
26. GENTRY, A. 1991b. El bosque nublado de Colombia. Págs. 14-20 en: C. Uribe (ed.), Bosques de niebla de Colombia. Banco de Occidente, Cali. [ Links ]
27. GENTRY, A. 1995. Patterns of diversity and floristic composition in Neotropical montane forests. Págs. 103-133 en: S. P. Churchill, H. Balslev, E. Forero & J. L. Luteyn (eds), Biodiversity and conservation of Neotropical montane forests. The New York Botanical Garden, New York. [ Links ]
28. GRADSTEIN, S., G. VAN REENEN & D. GRIFFIN. 1995. Origen de la flora de briófitos en el transecto Parque Los Nevados (Cordillera Central, Colombia). Págs. 377-384 en: T. van der Hammen & A. G. Dos Santos (eds.), La Cordillera Central Colombiana, transecto Parque de los Nevados (tercera parte). Estudios de Ecosistemas Tropandinos vol 4. J. Cramer, Berlín. [ Links ]
29. HAMILTON, L.S. 2001. Una campaña por los bosques nublados: ecosistemas únicos y valiosos en peligro. Págs. 41-49 en: M. Kappelle & A.D. Brown (eds.), Bosques nublados del neotrópico. Instituto Nacional de Biodiversidad (INBio), Santo Domingo de Heredia. [ Links ]
30. HERNÁNDEZ-CAMACHO, J.I. 1990. La selva en Colombia. Págs. 13-47 en: J. Carrizosa-U. & J.I. Hernández-Camacho (eds.), Selva y Futuro. El Sello Editorial, Bogotá. [ Links ]
31. HENDERSON, A., S.P. CHURCHILL & J.L. LUTEYN. 1991. Neotropical plant diversity. Nature 351: 21-22. [ Links ]
32. HOFSTEDE, R. 1995. Effects of burning and grazing on a Colombian páramo ecosystem. Ph. D. Dissertation, University of Amsterdam, Amsterdam. [ Links ]
33. JØRGENSEN, P.M., C. ULLOA-ULLOA, J.E. MADSEN & R. VALENCIA-R. 1995. A floristic analysis of the high Andes of Ecuador. Págs. 221-237 en: S.P. Churchill, H. Balslev, E. Forero & J.L. Luteyn (eds.), Biodiversity and conservation of Neotropical montane forests. The New York Botanical Garden, New York. [ Links ]
34. KAPPELLE, M. & A.D. BROWN (Eds). 2001. Bosques nublados del neotrópico. Instituto Nacional de Biodiversidad (INBio), Santo Domingo de Heredia. [ Links ]
35. KATTAN, G. & G. ALVAREZ-LÓPEZ. 1995. Preservation and management of biodiversity in fragmented landscapes in the Colombian Andes. Págs. 3-18 en: J. Schelhas & R. Greenberg (eds.), Forest patches in tropical landscapes. Island Press, Washington D.C. [ Links ]
36. KESSLER, M. 2001. Maximum plant-community endemism at intermediate intensities of anthropogenic disturbance in Bolivian montane forests. Conservation Biology 15 (3): 634-641. [ Links ]
37. KRAMER, K.U. & P.S. GREEN. 1990. Pteridophytes and Gymnosperms. Págs. 284-391 en: K. Kubitzki (ed.), The families and genera of vascular plants. Vol. 1. Springer-Verlag, Berlín. [ Links ]
38. LOZANO-C, G. & M. ALVEAR. 2001. Novedades en Axinaea y Meriania (Melastomataceae) de Colombia. Caldasia 23 (1): 147-152. [ Links ]
39. MAGURRAN, A.E. 1989. Diversidad ecológica y su medición. Ediciones Vedrá, Barcelona. [ Links ]
40. MARÍN-C., C. & J. BETANCUR. 1997. Estudio florístico en un robledal del Santuario de Flora y Fauna de Iguaque (Boyacá, Colombia). Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 21 (80): 249-259. [ Links ]
41. MITTERMEIER, R.A., N. MYERS, P. ROBLES-GIL, & C.G. MITTERMEIER. 1999. Hotspots. CEMEX, México. [ Links ]
42. MORA-O., L.E. & H. STURM. 1994. Resumen y conclusiones. Págs. 697-715 en: L.E. Mora-O. & H. Sturm (eds.), Estudios ecológicos del páramo y del bosque altoandino, Cordillera Oriental de Colombia. Tomo II. Academia Colombiana de Ciencias Exactas, Físicas y Naturales. Colección Jorge Alvarez Lleras No. 6. [ Links ]
43. MORENO, N.P. 1984. Glosario botánico ilustrado. Instituto Nacional de Investigaciones sobre Recursos Bióticos. Compañía Editorial Continental S. A, México. [ Links ]
44. MORI, S. & B. BOOM. 1983. Ecological importance of Myrtaceae in an Esatern Brazilian forest. Biotropica 15 (1): 68-70. [ Links ]
45. MUELLER-DOMBOIS, D. & H. ELLENBERG. 1974. Aims and methods of vegetation ecology. John Willey & Sons, Inc., New York. [ Links ]
46. MURCIA, C. 1997. Evaluation of Andean alder as a catalyst for the recovery of tropical cloud forest in Colombia. Forest Ecology 99: 163-170. [ Links ]
47. PÉREZ-ARBELÁEZ, E. 1996. (reimpresión). Cuencas hidrográficas y conservación de recursos naturales. Segunda edición. Fondo FEN Colombia, Bogotá. [ Links ]
48. PHILLIPS, O. & J.S. MILLER. 2002. Global patterns of plant diversity: Alwyn H. Gentry´s forest transect data set. Monographs in Systematic Botany from the Missouri Botanical Garden 89: 1-319. [ Links ]
49. RANGEL-CH., J.O. 1995a. La diversidad florística en el espacio andino en Colombia. Págs 187-205 en: S.P. Churchill, H. Balslev, E. Forero & J.L. Luteyn (eds.), Biodiversity and conservation of Neotropical montane forests. The New York Botanical Garden, New York. [ Links ]
50. RANGEL-CH., J.O. 1995b. Consideraciones sobre la diversidad y la vegetación de alta montaña en Colombia. Págs. 33-60 en: J.A. Lozano & J.D. Pabón (eds.), Memorias del Seminario Taller sobre Alta Montaña Colombiana (Santafé de Bogotá, 13-15 de octubre de 1993). Academia Colombiana de Ciencias Exactas, Físicas y Naturales. Colección Memorias No. 3. Bogotá. [ Links ]
51. RANGEL-CH., J.O. 1995c. Diversidad y frecuencia de las familias, géneros y especies de plantas vasculares en el transecto Parque de los Nevados. Págs 419-429 en: T. van der Hammen & A. Dos Santos (eds.), La Cordillera Central Colombiana, transecto Parque de los Nevados (tercera parte). Estudios de Ecosistemas Tropandinos vol 4. J. Cramer, Berlín. [ Links ]
52. RANGEL-CH., J.O. & P. FRANCO-ROSSELLI. 1985. Observaciones fitoecológicas en varias regiones de vida de la Cordillera Central de Colombia. Caldasia (14) 67: 211-249. [ Links ]
53. RANGEL-CH., J.O. & A. VELÁZQUEZ. 1997. Métodos de estudio de la vegetación. Págs 59-88 en: J.O. Rangel-Ch., P.D. Lowy-C. & M. Aguilar-P. (eds.), Colombia Diversidad Biótica II, tipos de vegetación en Colombia. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá. [ Links ]
54. RANGEL-CH., J.O., J.M. IDROBO, A.M. CLEEF & T. VAN DER HAMMEN. 1995. Segunda lista del material herborizado en el transecto Parque de los Nevados. Págs 385-418 en: T. van der Hammen & A.G. Dos Santos (eds.), La Cordillera Central Colombiana, transecto Parque de los Nevados (tercera parte). Estudios de Ecosistemas Tropandinos vol 4. J. Cramer, Berlín. [ Links ]
55. RESTREPO, D. 1999. Diagnóstico del impacto pecuario y de cultivos en la Finca Torre Cuatro, con propuesta de sistemas sostenibles agroforestales. Informe Técnico, CORPOCALDAS, Manizales. [ Links ]
56. RODRÍGUEZ-MAHECHA, J.V., P. SALAMAN, P. JØRGENSEN, T. CONSIGLIO, L. SUÁREZ, F. ARJONA, & R. BENSTED-SMITH. 2004. Tropical Andes. Págs. 73-79 en: R.A. Mittermeier, P. Robles-Gil, M. Hoffmann, J.D. Pilgrim, T.M. Brooks, C.G. Mittermeier, & G. Fonseca (eds.), HOTSPOTS REVISITED: EARTH'S BIOLOGICALLY RICHEST AND MOST ENDANGERED ECOREGIONS. SECOND EDITION. CEMEX, MÉXICO. [ Links ]
57. SALAMANCA, S. 1988. Los bosques altoandinos. Colombia, sus Gentes y sus Regiones 9: 19-29. [ Links ]
58. SALAMANCA, S. 1991. La vegetación de páramo y su dinámica en el macizo volcánico Ruiz-Tolima. Análisis Geográficos 21: 1-155. [ Links ]
59. SALAMANCA, S, A.M. CLEEF & J.O. RANGEL-CH. 2003. The paramo vegetation of the volcanic Ruiz-Tolima massif. Págs 1-78 en: T. van der Hammen & A.G. Dos Santos (eds.), La Cordillera Central Colombiana, transecto Parque de los Nevados (úlitma parte). Estudios de Ecosistemas Tropandinos vol 5. J. Cramer, Berlín [ Links ]
60. SUÁREZ-NAVARRO, A.E., G. HURTADO-PEÑA, F.J. CARVAJAL-LEMUS, J.E. RODRÍGUEZ-BAUQERO & R. RODRÍGUEZ-SOTO. 1984. Mapa de bosques de Colombia, memoria explicativa. IGAC, INDERENA & CONIF, Bogotá. [ Links ]
61. TILMAN, D. & S. PACALA. 1993. The maintenance of species richness in plant communities. Págs. 13-25 en: D. Shulter & R. Ricklefs (eds.), Species diversity in ecological communities, historical and geographical perspectives. The University of Chicago Press, Chicago & London. [ Links ]
62. ULLOA ULLOA, C. & P.M. JØRGENSEN. 2005. Endemic plant genera of the Tropical Andes. Database v. Jan 2005. Missouri Botanical Garden, St. Louis. [ Links ]
63. VALENCIA, R. & P.M. JØRGENSEN. 1992. Composition and structure of a humid montane forest on the Pasachoa volcano, Ecuador. Nordic Journal of Botany 12: 239-247. [ Links ]
64. VAN DER HAMMEN, T. 1995. Global change, biodiversity, and conservation of Neotropical montane forest. Págs. 603-607 en: S.P. Churchill, H. Balslev, E. Forero & J.L. Luteyn (eds.). Biodiversity and conservation of Neotropical montane forests. The New York Botanical Garden, New York. [ Links ]
65. VAN DER HAMMEN, T. 1998. Plan ambiental de la cuenca alta del río Bogotá, análisis y orientaciones para el ordenamiento territorial. Corporación Autónoma Regional de Cundinamarca (CAR), Bogotá. [ Links ]
66. VAN DER HAMMEN, T., H.J.L. WITTE & G.B.A. VAN REENEN. 1995. Aspectos ecoclimatológicos del área del transecto Parque Los Nevados. Págs. 329-364 en: T. van der Hammen & A.G. Dos Santos (eds.), La Cordillera Central Colombiana, transecto Parque de los Nevados (tercera parte). Estudios de Ecosistemas Tropandinos vol 4. J. Cramer, Berlín. [ Links ]
67. VAN DER HAMMEN, T. & J.O. RANGEL-CH. 1997. El estudio de la vegetación en Colombia. Págs. 17-57 en: J.O. Rangel-Ch., P.D. Lowy-C. & M. Aguilar-P. (eds.), Colombia Diversidad Biótica II, tipos de vegetación en Colombia. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá. [ Links ]
68. VÉLEZ-PALACIO, J. 1996. Avalúo del inmueble Torre Cuatro. En: Escrituras de propiedad del predio Torre Cuatro. CORPOCALDAS, Manizales. [ Links ]
69. VIS, M. 1995. Proceses and patterns of erosion in natural and disturbed Andean forest ecosystems. Págs. 17-182 en: T. van der Hammen & A.G. Dos Santos (eds.), La Cordillera Central Colombiana, transecto Parque los Nevados (tercera parte). Estudios de Ecosistemas Tropandinos vol 4. J. Cramer, Berlín. [ Links ]
70. WITTE, H.J.L. 1995. Distribución estacional y altitudinal de la precipitación, la temperatura y la humedad relativa en el transecto Parque de Los Nevados. Análisis Geográficos 25. [ Links ]
71. WHITE, P. & S. PICKETT. 1985. Natural disturbance and patch dynamics: An introduction. Págs. 3-13 en: S. Pickett & P. White (eds.), The ecology of natural disturbance and patch dynamics. Academic Press, San Diego. [ Links ]

