










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.32 no.1 Bogotá Jan./June 2010
Bats from a forest in the Central Andes of Colombia with notes on their taxonomy and distribution
MIGUEL E. RODRÍGUEZ-POSADA
Grupo de investigación en conservación y manejo de vida silvestre, Universidad Nacional de Colombia. Bogotá, D.C. Colombia. migrodriguezp@yahoo.com
RESUMEN
Se presenta la diversidad de murciélagos registrados en un inventario rápido en tres estaciones entre los 2500 y los 3500 m de altitud, sobre la vertiente occidental de la cordillera Central de los Andes colombianos, en el departamento de Caldas. El gradiente de distribución de Anoura aequatoris es ampliado hacia el norte y se sugiere su presencia en otras localidades más septentrionales de la cordillera Central y la Cordillera Oriental. Sturnira aratathomasi, S. erythromos y S. ludovici, fueron capturadas en simpatría mientras que Histiotus montanus e H. humboldti se registraron en estaciones diferentes. Myotis keaysi presentó el gradiente altitudinal más amplio, mientras Carollia brevicauda estuvo restringida a la estación de menor altitud. Los géneros Anoura, Sturnira e Histiotus presentan un patrón de sucesión o reemplazo altitudinal de especies. Considerando la confusión en la identificación de las especies registradas, se presentan comentarios sobre la diferenciación entre ellas y recomendaciones para estudios futuros sobre su taxonomía y distribución. La certeza taxonómica es importante para conocer el estado real de las poblaciones colombianas de murciélagos e identificar patrones básicos para interpretar procesos importantes dentro de los ecosistemas y su estado de conservación. Por el contrario, la información errada, puede implicar consecuencias inmensurables cuando es utilizada y los errores replicados.
Palabras clave. Anoura aequatoris, Andes Centrales de Colombia, Histiotus, Inventario rápido, murciélagos.
ABSTRACT
This paper reports the bat diversity found in a rapid assessment inventory in three stations between 2500 and 3500 masl, along the western slopes of the cordillera Central of Colombia (Departmento of Caldas). The distributional range of Anoura aequatoris is expanded northwards and its presence in more western localities of the the cordillera Central and the Oriental is suggested. Sturnira aratathomasi, S. erythromos and S. ludovici were captured in sympatry while Histiotus montanus and H. humboldti were recorded in different stations. Myotis keaysi showed the broadest altitudinal range, while Carollia brevicauda was only found in the station at lowest elevation. The genera Anoura, Sturnira and Histiotus have a pattern of altitudinal succession or replacement of species. Considering the difficulty to identify the recorded species, comments on their taxonomy and recommendations for future studies are included. Taxonomic accuracy is important to know the real status of the Colombian populations of bats and to identify basic biological patterns that allow us to interpret processes within ecosystems and their conservation status; using wrong information and repeating mistakes might bring immeasurable consequences.
Key words. Anoura aequatoris, Bats, Colombian Andes, Histiotus, rapid assessment inventory.
Recibido: 20/05/2009
Aceptado: 21/08/2009
INTRODUCCIÓN
Los Andes tropicales son considerados áreas prioritarias de conservación a nivel global, por su riqueza biológica, alto nivel de endemismos y por la pérdida acelerada de hábitat (Myers et al. 2000, Armenteras et al. 2003). Debido a su complejidad estructural, historia geológica y climática y su posición tropical, los Andes colombianos son megadiversos, tanto en riqueza de ecosistemas y hábitats, como en grado de endemismo y riqueza de especies. Lamentablemente han sido sometidos a una gran transformación de sus áreas naturales, y se estima que prevalece del 18 al 25% del bosque andino original (Armenteras et al. 2003, Kattan et al. 2004); por lo tanto, es necesario realizar inventarios y documentar sus resultados con el fin de aumentar el conocimiento actual sobre la magnitud y la distribución geográfica de los componentes de su diversidad biológica, estructurar el conocimiento base para la evaluación de prioridades de investigación y conservación, y posteriormente, desarrollar estrategias de manejo de los recursos biológicos, esenciales para el establecimiento de legislaciones y otras acciones para preservar dichos recursos para el futuro (Voss & Emmons 1996, Wilson et al. 1996, Stiles & Roselli 1998).
La fauna quiróptera de los Andes colombianos, comprende cerca del 59% de las especies de murciélagos registradas para el país (Castaño et al. 2004). La mayoría de estudios sobre murciélagos en ecosistemas andinos colombianos ha sido enfocada a nivel de ecología de comunidades, inventarios o listas regionales (e.g., Muñoz 1990, Alfonso-B & Cadena 1994, Cadena et al. 1998, Castaño et al. 2003, Castaño et al. 2004, Cuartas-Calle & Muñoz-Arango 2003, Numa et al. 2005, Bejarano-Bonilla et al. 2007). Sin embargo, muchos estudios sobre diversidad carecen de información sobre criterios para identificación de las especies y en algunos casos sobre la metodología e intensidad del muestreo, limitando la certeza taxonómica requerida para analizar la información y la posibilidad de comparar entre estudios de diferentes localidades. Especialmente, si se tiene en cuenta que muchas especies de murciélagos tropicales, si no la mayoría, son difíciles de identificar acertadamente sin examinar la morfología cráneo-dental, ya que presentan diferencias sutiles, que al no ser revisadas detalladamente, producen identificaciones erróneas (Simmons & Voss 1998).
En este artículo se presentan los resultados de un inventario rápido de murciélagos realizado en tres localidades de la vertiente occidental de la Cordillera Central de los Andes colombianos. Con el fin de contribuir a futuros estudios sobre biodiversidad, se presentan comentarios sobre la identificación, la taxonomía, la distribución y la historia natural de las especies registradas.
MATERIALES Y MÉTODOS
Área de estudio. Los muestreos se realizaron en la Reserva Hidrográfica Río Blanco, ubicada en la vertiente occidental de la Cordillera Central, tres km al noroeste de la ciudad de Manizales, en el departamento de Caldas, Colombia (5º 04' Norte; 75º 32' Oeste). La reserva tiene un área de 43.43 km2 sobre la cuenca del Río Blanco, en un gradiente altitudinal entre 2240 y 3750 m. La precipitación anual es cercana a los 2500 mm, con un régimen de lluvias bimodal-tetraestacional, con dos picos de lluvias, el primero entre marzo y mayo, y el segundo entre septiembre y noviembre. La temperatura promedio anual es de 11ºC.
La vegetación original de la Reserva fue fuertemente intervenida hasta su protección. Actualmente prevalecen cinco unidades de vegetación: bosque nativo, reforestación con aliso (Alnus acuminata), potreros, bosque en regeneración y reforestación con especies exóticas (Sánchez et al. 2004). Los muestreos se realizaron en tres localidades: 1- Caldas, Manizales, vereda Las Palomas, Reserva Hidrográfica Río Blanco, Fundación Ecológica Gabriel Arango Restrepo, estación Viveros, altitud: 2500 m; vegetación compuesta por bosques subandinos y andinos en diferentes grados de regeneración y reforestaciones mixtas de eucalipto y aliso, con sotobosques en diferentes etapas de sucesión (5º04´ Norte, 75º27´ Oeste). 2- Caldas, Manizales, vereda El Desquite, corregimiento 8, Reserva hidrográfica Río Blanco, finca La Navarra, estación La Navarra, altitud: 3000 m; vegetación compuesta por reforestación de aliso con sotobosque en diferentes grados de sucesión (5º05´ Norte, 75º25´ Oeste). 3- Caldas, Manizales, vereda El Desquite, corregimiento 7, Reserva hidrográfica Río Blanco, finca Martinica, estación Martinica, altitud: 3500 m; vegetación compuesta por pastizales abiertos con vegetación arbustiva y remanentes de bosque Andino y Altoandino (5º04´ Norte, 75º23´ Oeste). Información más detallada sobre el área de estudio se puede encontrar en los trabajos de Sánchez et al. (2004, 2008).
Muestreo. El muestreo se realizó entre el 26 de enero y el 7 de febrero del año 2004; los murciélagos se capturaron utilizando redes de niebla estándar con ojo de malla de 38 mm, 2.5 m de alto y 6 o 12 m de ancho. En promedio se utilizaron 64 m de red por jornada de muestreo con un gradiente entre 42 y 96 m de red. Las redes se cambiaron de sitio en cada jornada de muestreo para evitar que los murciélagos las detectaran. En total, se usaron 762 m de red, durante 59 horas (en promedio cinco horas por jornada de muestreo), resultando un esfuerzo total de 3746.5 metros-red-hora (mrh., medido como el producto de los metros lineales de red utilizados por las horas de muestreo promedio, Simmons & Voss 1998). El esfuerzo de muestreo en la estación Viveros fue de 1033.2 mrh, en La Navarra 1505.5 mrh y en Martinica de 1210.3 mrh.
Los murciélagos capturados se identificaron preliminarmente en campo y les fueron tomadas las medidas morfológicas externas estándar (longitud total, longitud cola, longitud pie, longitud oreja y longitud antebrazo), siguiendo a Simmons & Voss (1998). Se determinó el sexo y el estado reproductivo de cada individuo, los machos se clasificaron como reproductivos si tenían los testículos en posición escrotal, o no reproductivos con los testículos en posición inguinal; las hembras se clasificaron como gestantes, lactantes o no reproductivas (Tirira 1998). Las clases de edad se asignaron de acuerdo con la osificación de las epífisis de las falanges de las alas y el tipo de dentición (Kunz & Kurta 1988): se consideraron infantes aquellos murciélagos que todavía estaban lactando y presentaban dentición de leche (dientes en forma de gancho); los individuos jóvenes presentaron dentición definitiva y con las epífisis de las falanges del ala con osificación incompleta; los adultos presentaron dentición definitiva (generalmente con algún grado de desgaste) y falanges completamente osificadas.
Con el fin de confirmar la identificación de las especies se recolectó al menos un espécimen de referencia por cada especie detectada y todos los individuos cuya identificación fue dudosa. Los especímenes se conservaron como piel de estudio con cráneo limpio y cuerpo en líquido, o espécimen completo en líquido, fijado en formol 10% y luego transferido a etanol 70%. Los especímenes se depositaron en la colección de mamíferos del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia (ICN), con los números de catálogo desde el ICN16971 al 17003.
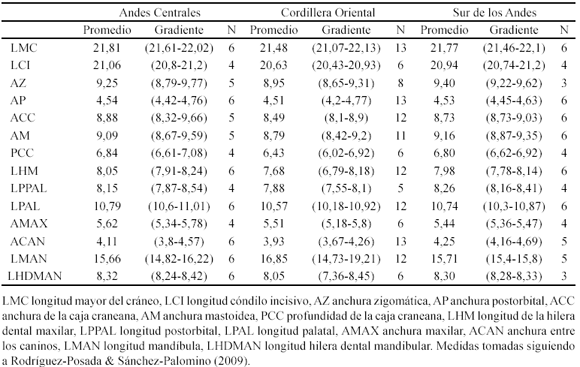
Las medidas craneanas se tomaron siguiendo a Rodríguez-Posada & Sánchez-Palomino (2009). Los metacarpos se midieron con el ala plegada, desde la superficie distal de los carpos hasta el extremo distal del metacarpo. Las falanges se midieron entre sus extremos distal y proximal.
La taxonomía se presenta de acuerdo con Simmons (2005), a excepción de los cambios más recientes que se tratan en los comentarios de cada especie. Para la identificación definitiva se utilizó literatura especializada citada en los comentarios de cada especie y comparaciones con especímenes de otras localidades depositados en las colecciones de mamíferos del Instituto de Ciencias Naturales (ICN), Museo de Historia Natural Ginebra (MHNG) y la colección de mamíferos del Museo de Historia Natural de la Universidad del Cauca (MHNUC).
Finalmente, se presenta una lista comentada con información sobre las especies registradas durante el muestreo, incluyendo el número de catálogo de los especímenes recolectados, número de individuos capturados, sexo, categoría de edad y estado reproductivo, sitios donde fueron capturados y tipo de vegetación circundante. Adicionalmente, se presentan comentarios sobre la identificación, la taxonomía y la distribución de cada especie, incluyendo literatura pertinente, comparaciones con especímenes de otras localidades y una lista del material examinado de otras localidades.
RESULTADOS
Se capturaron 35 murciélagos correspondientes a ocho especies, seis de la familia Phyllostomidae y dos de la familia Vespertilionidae (Tabla 1).
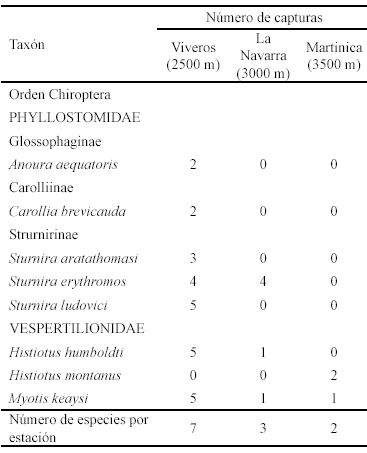
Éxito del muestreo. La estación Viveros presentó el mayor número de capturas (26) y de especies registradas (7). En las estaciones La Navarra y Martinica el número de capturas fue bajo (6 y 3), al igual que el número de especies registradas (3 y 2).
LISTA COMENTADA
FAMILIA PHYLLOSTOMIDAE
Subfamilia GLOSSOPHAGINAE
Anoura aequatoris (Lönnberg 1921)
Se capturaron dos individuos (ICN 16971, 16972) en la estación Viveros (2500 m). Los individuos correspondieron a una hembra no reproductiva y un macho con testículos abdominales. Uno en una reforestación mixta de aliso y eucalipto con vegetación natural en sotobosque y otro en un bosque subandino en regeneración.
Comentarios. En Colombia se han registrado las ocho especies del género Anoura: A. geoffroyi, A. latidens, A. caudifer, A. cultrata, A. luismanueli, A. cadenai, A. aequatoris y A. fistulata (Alberico et al. 2000, Mantilla-Meluk & Baker 2006, 2008). Los individuos capturados en Río Blanco fueron identificados ambiguamente como A. caudifer, A. luismanueli o A. aequatoris, según los criterios de Tamsitt & Nargosen (1982), Molinari (1994) y Mantilla-Meluk & Baker (2006). Al comparar con especímenes de otras localidades en los Andes por encima de 1000 m, provenientes de las regiones de diferenciación geográfica de estos tres taxones en Colombia (Mantilla-Meluk & Baker 2006. 1- A. aequatoris, poblaciones al sur de los 4º N sobre los Andes; 2- A. caudifer, poblaciones del centro y norte de las Cordilleras Occidental y Central; 3- A. luismanueli, Cordillera Oriental), sé encontró sobreposición en las medidas morfométricas (Tabla 2) y que todos los especímenes comparten una combinación de caracteres morfológicos con los que se pueden identificar como A. aequatoris o A. luismanueli (Molinari 1994, Mantilla-Meluk & Baker 2006): uropatagio semicircular con una orla de pelos a lo largo del margen libre (la longitud y densidad de los pelos de la orla varió individualmente y no geográficamente), calcáneo pequeño (menor a 4mm), coloración dorsal varió entre negro y pardo rojizo, con dos bandas, la distal oscura y corta (1/3-1/5), y la basal blanca de mayor longitud (2/3 a 4/5).
La hilera dental maxilar recta (excepto tres especímenes de Nariño que la presentaron ligeramente convexa a la altura de los premolares, ICN 13633-13636), el paladar ornamentado con surcos transversales y la caja craneana ligeramente cuadrangular. Finalmente, no se encontró algún carácter continuo o discreto que permitiera diferenciar A. luismanueli y A. aequatoris, y las observaciones morfológicas no apoyan la propuesta de diferenciación geográfica de Mantilla-Meluk & Baker (2006). Teniendo en cuenta las características morfológicas y la distribución geográfica, los especímenes de Río Blanco fueron identificados como A. aequatoris y se sugiere que poblaciones de otras localidades al norte de las Cordilleras Central y Oriental deben ser asignados a este taxón. Sin embargo, las relaciones y los límites entre estos taxones no son claros y es necesario realizar una revisión exhaustiva, utilizando conjuntos de caracteres morfológicos adicionales y análisis moleculares, incluyendo series de especímenes de todo el gradiente de distribución de A. caudifer en el sentido dado por Tamsitt & Valdivieso (1966) y Koopman (1994).
Otros especímenes examinados (32): COLOMBIA. Antioquia: Jardín, vereda La Linda, quebrada La Linda, 2200 m (ICN 16455). Antioquia: Medellín, Parque Regional Aruí, sitio La Aguada, 2500 m (ICN 18045, 18047, 18048). Caldas: Manizales, quebrada Guayabal, Jardín de Mariposas, Recinto del Pensamiento, Maltería, 2180 m (ICN 16729). Cauca (MHNG 1908.007). Cauca: El Tambo, Parque Nacional Natural Munchique (MHNG 1907.061). Nariño: Barbacoas, corregimiento de Altaquer, 1400 m (13633, 13636-13638). Nariño: Barbacoas, corregimiento Junín, 900 m (13634-13635). Santander: Charalá, Inspección de policía Virolín, 1790 m (ICN 6602-6608, 8123, 8124, 8981). Santander: Encino, vereda Rionegro, Reserva Biológica Cachalú, 1900 m (ICN 17520-17524). Santander: Suaita, vereda La Meseta, Inspección de policía San José de Suaita, quebrada La Espumosa, 1580 m (ICN 15295-15298). ECUADOR: Pastaza, Mera, 2700 m (MHNG 1682.80).
Subfamilia CAROLLIINAE
Carollia brevicauda (Schinz 1821)
Se capturaron dos hembras adultas no reproductivas (ICN 16973, 16974) en la estación Viveros (2500 m), en un bosque subandino en regeneración y en una reforestación mixta de aliso y eucalipto con sotobosque denso.
Comentarios. Actualmente el género Carollia está compuesto por nueve especies (Solari & Baker 2006), de las cuales se han registrado cinco en Colombia: C. perspicillata, C. brevicauda, C. castanea, C. colombiana y C. monohernandezi (Alberico et al. 2000, Cuartas et al. 2001, Muñoz et al. 2004). Los dos individuos capturados coinciden morfométrica y morfológicamente con las características que separan a C. brevicauda de las otras especies del género: externamente presentan pelaje largo y denso de color gris oscuro, que se extiende a lo largo de la superficie dorsal de los antebrazos, y pelaje dorsal con tres bandas, la basal oscura y ancha, la intermedia blanquecina muy notoria y la distal oscura con las puntas anteadas. A nivel cráneo-dental, el margen labial de la hilera dental maxilar es cóncavo, la cúspide estilar anterior del premolar posterior superior (parástilo del P4) es reducida y no tiene contacto con el premolar anterior superior (P3), los márgenes linguales de los premolares inferiores (p4 y p3) forman un ángulo convexo, los incisivos externos inferiores (i2) no son reducidos y son visibles en vista oclusal (Pine 1972, Cloutier & Thomas 1992, Koopman 1994, Cadena et al. 1998, Cuartas et al. 2001, Muñoz et al. 2004, Pacheco et al. 2004).
Subfamilia STENODERMATINAE
Sturnira aratathomasi Peterson & Tamsitt 1968
Se registraron dos hembras adultas no reproductivas y una lactante, los tres individuos (ICN 16988-16990) fueron capturados en la estación Viveros (2500m), en un bosque subandino en regeneración
Comentarios. Entre las tres especies de Sturnira registradas, S. aratathomasi es la más fácil de identificar debido a su mayor tamaño (longitud del antebrazo mayor a 54 mm), rostro e incisivos centrales superiores proyectados hacia delante y presencia de entocónido en el primer molar inferior (Peterson & Tamsitt 1968, Davis 1980, Soriano y Molinari 1984). Estrada-Villegas et al. (2007) revisaron la distribución de esta especie, incluyendo este registro en sus resultados.
Sturnira erythromos (Tschudi 1844)
Se registraron ocho individuos en la Reserva (ICN 16991-16998): una hembra gestante con embrión completamente desarrollado, una hembra lactante, dos hembras adultas no reproductivas y cuatro machos adultos con testículos abdominales. La especie fue registrada en las estaciones Viveros (2500 m) y La Navarra (3000 m); en bosques subandinos y andinos en regeneración, y en reforestaciones de aliso y eucalipto con sotobosque en diferentes estados de sucesión.
Comentarios. La identificación de los murciélagos del género Sturnira es complicada, algunos caracteres como la morfología de la superficie masticatoria de los incisivos y coloraciones de pelaje son ambiguos o subjetivos, algunas medidas morfológicas se sobreponen y el género no está bien comprendido taxonómicamente (Sánchez-Hernández et al. 2005). Para la identificación se siguió a Davis (1980), Pacheco & Patterson (1991) y Giannini & Barquez (2003), pero los individuos fueron identificados ambiguamente como S. erythromos o S. bogotensis. Todos los especímenes examinados presentaron los incisivos superiores centrales con el borde cortante entero, los inferiores levemente bilobulados, el paladar duro plano y el borde lingual del primer molar inferior (m1) sin entocónido. Linares (1987), diferenció S. bogotensis con incisivos superiores bilobulados, mientras que en S. erythromos el borde cortante sería entero, sin embargo este carácter es afectado por el desgaste. Las medidas de antebrazo del material examinado (40-43.8 mm, n= 6), se sobreponen con las propuestas por Giannini & Barquez (2003) para las dos especies (S. erythromos 38-42 mm y S. bogotensis 43-45.3 mm). Conteras (1999), utilizó el grado de desarrollo de la hendidura trigónica y del paraconido en m1 para diferenciar las dos especies, los especímenes examinados presentaron la hendidura trigónica bien desarrollada hasta la base del cono en m1 y el paracónido bien desarrollado (Fig. 1), correspondiendo a S. erythromos, a diferencia de S. bogotensis que no presenta hendidura trigónica en m1, observándose un muro reducido entre metacónido y paracónido (Contreras 1999).
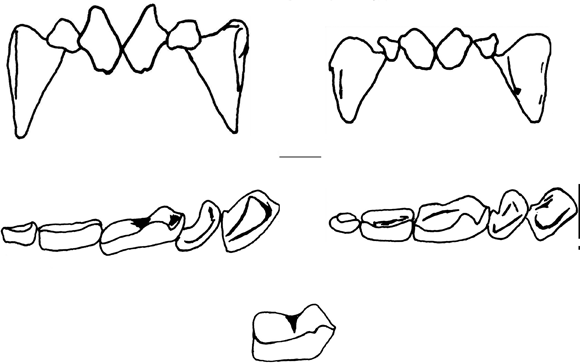
En la clave para el género Sturnira propuesta por Muñoz (2001), se encuentran dos errores que dificultan la correcta identificación de las especies del género en Colombia, el primero es que incluye a S. thomasi, especie que no se encuentra en el país (Contreras 1999, Alberico et al. 2000, Simmons 2005) y el segundo es que incluye a S. bogotensis entre las especies con -muesca lingual- en los molares, es decir con entocónido en el m1, agrupándola junto a S. aratathomasi, S. tildae, S. thomasi, S. luisi y S. lilium. Pero S. bogotensis no presenta entocónido en m1, carácter que comparte son S. erythromos, S. koopmahilli, S. magna, S. mordax, S. ludovici y S. sorianoi.
Otro material examinado (1): PERÚ. (MHNG 1021.59).
Sturnira ludovici Anthony 1924
Se capturaron cinco individuos (ICN 16999-17003), dos hembras gestantes con embrión desarrollado, dos hembras adultas no reproductivas y un macho con testículos abdominales. Todos los individuos fueron registrados en la estación Viveros (2500 m); en un bosque natural maduro con algún grado de intervención y en una reforestación de aliso y eucalipto con presencia de vegetación nativa.
Comentarios. Gardner (2007) trata S. ludovici como una subespecie de Sturnira oporaphilum, que correspondería a las poblaciones que habitan la parte norte de Suramérica; otros autores que han revisado Sturnira, consideraron S. ludovici como una especie válida (Davis, 1980; Pacheco & Patterson,1991; Contreras, 1999 y Giannini & Barquez (2003); Iudica (2000), basado en evidencia morfológica y molecular considera S. ludovici a nivel de especie, representada por especimenes de Ecuador y con los limites específicos por definir, sus resultados muestran a S. oporaphilum como el grupo hermano de S. ludivici y reconoce Sturnira hondurensis como especie válida restringida a Centroamérica. En vista que la asignación específica de las poblaciones de Colombia no es clara, se sigue a Simmons (2005) considerándolas como S. ludovici, siguiendo para su identificación a Davis (1980), Pacheco & Patterson (1991), Contreras (1999) y Giannini & Barquez (2003). Los especímenes identificados como S. ludovici se diferenciaron por la ausencia de entocónido, presencia de hendidura posterior y paracónido reducido en m1 y m2, incisivos superiores centrales rectangulares en contacto en la mitad de la corona (Fig. 1), e incisivos inferiores bilobulados. Otros caracteres que facilitan su identificación son: paladar duro cóncavo y m1 con la región trigónica más angosta que la talónica (Contreras 1999). La morfología de los incisivos centrales superiores y la estructura del borde lingual del m1 sirvieron para diferenciar a S. ludovici de S. erythromos en campo y laboratorio. La longitud del antebrazo (LA), no presentó sobreposición, siendo mayor en S. ludovici (LA 44-47.5 mm; n= 5), que en S. erythromos (LA 40-43.8 mm; n=8); sin embargo, en otras localidades de los Andes puede encontrarse sobreposición en esta medida.
FAMILIA VESPERTILIONIDAE
Histiotus humboldti Handley 1996
Se capturaron seis murciélagos de esta especie (ICN 16975-16979), incluyendo tres hembras adultas inactivas, dos preñadas y un macho adulto con testículos escrotales. La especie se registró en la estación Viveros (2500 m) y en la estación La Navarra (3000 m). Las capturas se hicieron en bosques subandinos y reforestaciones de aliso y eucalipto con sotobosque en diferentes estados de regeneración.
Comentarios. Se capturaron ocho individuos del género Histiotus que fueron identificados preliminarmente como H. montanus. Al revisar los especímenes con los cráneos extraídos limpios y compararlos con especímenes de otras localidades e imágenes del holótipo de Histiotus colombiae Thomas 1916 (BMNH 99.11.4.1), se confirmó que la muestra estaba compuesta por dos especies: H. montanus e H. humboldti. Aunque los especímenes de H. montanus de Ro Blanco fueron ligeramente más grandes que los de H. humboldti, no se encontró algún carácter externo que permitiera la identificación inequívoca de cada una de las especies. Handley (1996), menciona diferencias en las longitudes de la tibia y los elementos óseos de los dedos de las alas; sin embargo, en los especímenes de Río Blanco hubo sobreposición en estás medidas (Tabla 3).
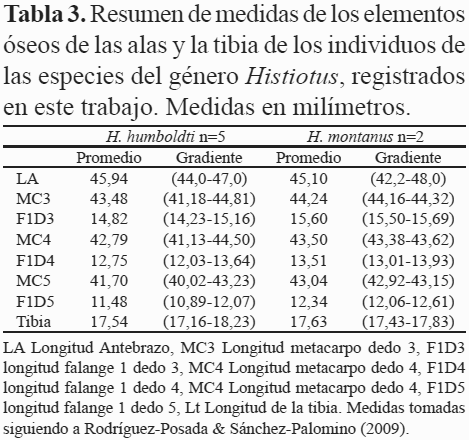
H. humboldti pudo diferenciarse de H. montanus por morfología craneana (Fig. 2), H. humboldti presenta el cráneo más pequeño que H. montanus.
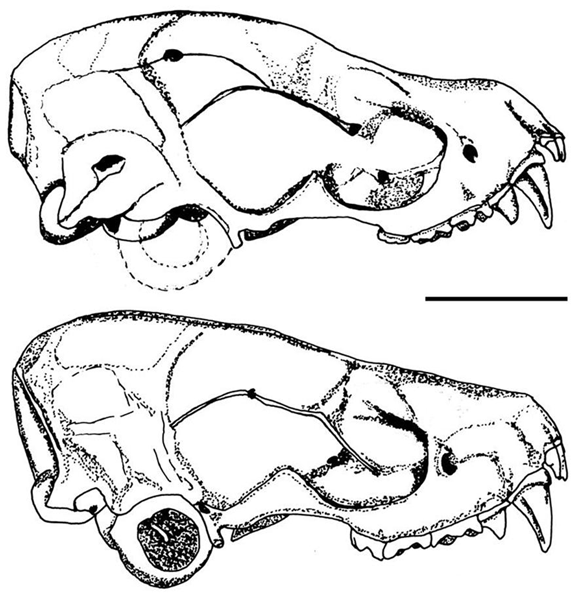
En vista lateral, H. humboldti presenta el perfil de la caja craneana horizontal seguido por una pendiente abrupta al nivel de los frontales que forma un ángulo con el rostro, nasales deprimidos, dando apariencia levemente respingada al rostro. H. montanus no presenta ángulo entre la caja craneana y el rostro, los nasales son abombados y el rostro no es respingado, dando al perfil del cráneo una apariencia de pendiente continua desde la región más posterior de la cresta sagital, hasta la punta de los nasales. La región supraorbital es abombada en ambas especies pero H. montanus la presenta mucho más abombada que H. humboldti. La región yugal del arco zigomático es ensanchada en H. montanus y delgada en H. humboldti. H. montanus presenta cresta sagital desarrollada continuando hacia las crestas lambdoideas formando una placa triangular engrosada en la región dorsocaudal del cráneo, en contraste H. humboldti no presenta cresta sagital desarrollada ni placa ósea. La dentición es más robusta en Histiotus montanus que en H. humboldti. Debido a la confusión en la identificación, se recomienda revisar la taxonomía y la distribución de las especies del género Histiotus del norte de los Andes, todos los especímenes revisados se encontraban erróneamente identificados como H. montanus y algunos de estos han sido registrados previamente en la literatura como tal (ICN 15160, Castaño et al. 2003; MHNUC 1058, 1395, Rivas-Pava et al. 2007). Muñoz (2001), omite la presencia de esta especie en Colombia. H. humboldti ha sido categorizada globalmente por la Unión Internacional para la Conservación de la Naturaleza (UICN), como especie con datos deficientes y endémica para Venezuela (Hutson et al. 2001).
Otro material examinado (5): COLOMBIA. Boyacá: Ráquira, Iglesia de Ráquira, 2150 m (MHNG 1910.074). Caldas: Manizales, vereda Las Palomas, Reserva Hidrográfica Río Blanco, Fundación Ecológica Gabriel Arango Restrepo, 2500 m (ICN 15160,). Risaralda: Santuario, vereda Campamento (ICN 11865), Cauca: Popayán, El Hogar, Bocatoma del acueducto, 1880 m (MHNUC 1395). Cauca: Totoró, Novirao, Resguardo indígena Novirao, Hacienda Novirao, 1745 m (MHNUC 1058).
Histiotus montanus (Philippi & Landbeck 1861).
Se registraron dos hembras adultas no reproductivas (ICN 16980, 16981). Las capturas se realizaron en la estación Martinica (3500 m) en el interior de un bosque altoandino en regeneración.
Comentarios. La apariencia general de los murciélagos de esta especie, es similar a H. humboldti, los individuos de H. montanus son proporcionalmente de mayor tamaño, aunque las medidas externas se sobreponen. La mayoría de las diferencias se presentan a nivel craneano (ver comentarios sobre la identificación de H. humboldti), por lo cual es necesario recolectar especímenes de referencia para confirmar su identificación. Los individuos registrados tienen el cráneo menos robusto y medidas inferiores a las observadas en el holótipo de H. montanus colombiae y en un espécimen (MHNG 1909.038) proveniente de una localidad cercana a la localidad tipo de H. montanus colombiae. Tradicionalmente se ha aceptado que esta especie se distribuye a lo largo de los Andes, desde el sur de Argentina y Chile, hasta Colombia y el occidente de Venezuela (Koopman 1994). Algunas subespecies que daban sentido a esta distribución son consideradas especies válidas en la actualidad (e.g., H. macrotus, H. laephotis, H. magellanicus; Simmons 2005), por lo tanto, la idea sobre su supuesta distribución continua de norte a sur no debe ser correcta (Barquez et al. 1999). Al revisar más especímenes a lo largo de toda la distribución de la especie, posiblemente H. montanus colombiae resulte valida a nivel específico. Es necesario clarificar la taxonomía y la distribución de las especies de Histiotus en Colombia.
Otro material examinado (1): COLOMBIA. Cundinamarca: Chipaque, Iglesia de Chipaque, 2400 m (MHNG 1909.038).
Myotis keaysi J. A. Allen 1914
Se registraron siete individuos (ICN 16982-16987) incluyendo dos hembras lactantes, dos machos adultos con testículos abdominales y un macho con testículos abdominales pero con los cauda epidídimo entre el uropatagio ubicados a los costados de la cola. La especie fue capturada en las tres estaciones muestreadas (2500 m, 3000 m y 3500 m), en interior de bosque subandino en regeneración, reforestaciones de aliso y eucalipto, y al interior de un bosque altoandino.
Comentarios. Las medidas morfológicas externas y características del pelaje como distribución cobre los patagios, largo, textura y coloración fueron insuficientes para identificar definitivamente los Myotis capturados en Río Blanco, siendo asignados ambiguamente como M. keaysi, M. nigricans o M. oxyotus, las tres especies de presencia probable en la zona de estudio (Alberico et al. 2000), y similares superficialmente cuando se encuentran en simpatría en la región Andina (LaVal 1973). Finalmente y siguiendo la única revisión disponible de Myotis del Neotrópico (LaVal 1973) y otros trabajos donde se revisan algunas especies de Myotis (Wilson & LaVal 1974, Koopman 1994, Simmons & Voss 1998, López-González et al. 2001, Hernandez-Mesa et al. 2005), los especímenes fueron identificados como M. keaysi, diferenciándose de las otras especies del género por presentar las crestas sagital y lambdoideas visibles y bien desarrolladas, constricción postorbital (POC) angosta en relación a la anchura entre los márgenes externos de los caninos (C-C), con un promedio de la relación POC/CC de 1.07 mm (n = 5), los tres premolares superiores incluidos en la hilera dental. Todos los individuos presentaron las extremidades y patágios con abundante pelo, extendiéndose dorsalmente sobre las extremidades inferiores sobre el fémur, ocasionalmente más allá de la rodilla. La coloración del pelaje varió individualmente entre pardo ferruginoso y pardo opaco.
DISCUSIÓN
El inventario tuvo una escala de tiempo breve y debido al método de muestreo la representatividad del ensamblaje puede ser incompleta y sesgada, los resultados no muestran completamente la riqueza y la abundancia de las especies, por lo que no es prudente hacer inferencias sobre la estructura del ensamblaje. El principal sesgo del muestreo fue el uso exclusivo de redes de niebla, método más efectivo para capturar filostómidos. Sin embargo, se registraron tres especies de murciélagos insectívoros de la familia Vespertilionidae, que potencialmente pueden detectar y evadir las redes. Se recomienda para estudios futuros utilizar un conjunto de métodos complementarios, como búsqueda de refugios, trampas harpa, etc., con el fin de registrar otras especies que utilizan los estratos superiores del bosque o que suelen evadir las redes gracias a sistemas de ecolocalización sofisticados (Kalko 1998). En el caso particular de este estudio, pudieron ser omitidos murciélagos de la familia Molossidae (e.g., Tadarida brasiliensis) cuya presencia es probable en el área de estudio.
Los ensamblajes de murciélagos responden a un patrón de disminución progresiva de la riqueza de especies a medida que aumenta la altitud (Graham 1990, Patterson et al. 1998); en Río Blanco, la estación de muestreo a menor altitud presentó la mayor riqueza de especies (7) y la de mayor altitud presentó la menor riqueza (2). La riqueza de especies fue similar a la encontrada en otros estudios a altitudes similares en la Cordillera Central (c.f. Muñoz 1990, Alfonso & Cadena 1994), sin embargo la composición varió y no se registraron especies de los géneros Artibeus, Dermanura, Enchistenes y Desmodus. La presencia de especies frugívoras debe estar influenciada por la cobertura vegetal de la Reserva, ya que los recursos alimentarios más abundantes son plantas típicas de procesos de regeneración (piperáceas, melastomatáceas, cecropiáceas y solanáceas) que favorecen la presencia de especies de Sturnira y Carollia que se alimentan de estas plantas. Las especies de frugívoros más específicos (e.g., Dermanura spp.), estarían restringidos a las zonas con vegetación nativa y a las temporadas de fructificación de las especies que consumen (e.g., Ficus spp.). La presencia de Desmodus rotundus ha sido sugerida previamente por registros indirectos (Sánchez et al. 2004).
En la estación Viveros, fueron registradas tres especies de Sturnira en simpatría y en la estación La Navarra solamente una. Este patrón de reemplazo o sucesión altitudinal se ha reportado en otras localidades, donde las especies de este género presentan diferencias en su distribución geográfica y muestran distribuciones altitudinales distintas con un área reducida de existencia simpátrica, lo que se expresa en un patrón de variación morfológica que sugiere un proceso de especiación parapátrica, desplazamiento de caracteres y exclusión competitiva (Jarrín 2005). H. humboldti se registró en las estaciones Viveros y Martinica e H. montanus se registró en la estación La Navarra; morfológicamente las dos especies presentan una gran similitud en apariencia externa y a su vez, una gran divergencia en morfología craneana. Se sugiere que H. montanus esta distribuida en zonas por encima de 3500 m, mientras que H. humboldti estaría distribuida a alturas menores en bosques andinos y subandinos entre los 1700 y 3000 m, en la franja altitudinal o ecológica de simpatría puede que la diferenciación externa sea más evidente. M. keaysi fue registrada en las tres estaciones y presenta un gradiente de distribución altitudinal amplio, los registros previos de M. nigricans y M. keaysi (Sánchez et al. 2004), fueron revisados y corresponden a M. keaysi, por lo cual esta especie no se encuentra en la reserva en simpatría con ningún congenérico, presentando una gran variación individual en morfología externa, al punto de confundirse con otras especies del género. Respecto a C. brevicauda y A. aequatoris, sólo se registraron en la estación más baja y teniendo en cuenta sus gradientes altitudinales conocidos, no debe encontrarse a mayores altitudes. En la Reserva se ha registrado Anoura geoffroyi (Sánchez et al. 2004), especie que tiene un gradiente de distribución altitudinal amplio y que puede estar presente hasta los 3500 m (Alberico et al. 2000), por lo cual, en Anoura también se puede presentar el patrón de reemplazamiento o sucesión altitudinal con divergencia morfológica sugerido para Histiotus y Sturnira.
La confusión en la identificación de especies similares morfológicamente y el desconocimiento de la magnitud de la variación intraespecífica, así como la falta de guías locales que faciliten la identificación de las especies en ecosistemas colombianos, dificultan la evaluación del estado de conservación de las especies, como es el caso entre M. keaysi y M. nigricans (Hernández-Mesa et al. 2005). Entre las especies registradas en este estudio se detectó que puede haber confusiones entre especies que son muy diferentes superficialmente en localidades diferentes, pero muy similares en simpatría, como M. nigricans caucensis, M. keaysi y M. oxyotus (LaVal 1973), así como la dificultad en la diferenciación entre especies como H. humboldti e H. montanus o S. erythromos y S. bogotensis. Es posible que algunos registros de estas especies estén confundidos y que el estado real de sus poblaciones, historia natural y distribución sea parcialmente desconocida, lo que lleva a que su estado de conservación y asignación de categoría de amenaza permanezcan inciertas. Es necesario asegurarse de la identidad taxonómica de las especies registradas en los estudios, en particular en ecosistemas tan vulnerables y desconocidos como los Andes septentrionales, ya que esto permite hacer comparaciones y evaluar realmente el estado de las poblaciones que componen su biota.
Finalmente, se recomienda a los investigadores ser más estrictos en la identificación de las especies en los estudios sobre murciélagos, así como recolectar series de especímenes y depositarlos en colecciones científicas, con el fin de conocer la variabilidad de las especies y la diferenciación de sus congenéricos más similares. Es necesario adelantar estudios para esclarecer la taxonomía y la distribución de las especies de murciélagos de Colombia, ya que la desactualización taxonómica distorsiona la utilidad de la información. Los registros errados pueden afectar la interpretación de los resultados, sobreestimando o subestimando la diversidad y dando lugar a interpretaciones distorsionadas de los resultados y sus conclusiones. La certeza taxonómica puede develar patrones básicos para interpretar procesos importantes dentro de los ecosistemas y su estado de conservación, de lo contrario puede implicar consecuencias inmensurables cuando la información errada es utilizada y los errores replicados.
AGRADECIMIENTOS
La fase de campo de este trabajo se realizó gracias a los aportes de la Fundación Gabriel Arango Restrepo y del laboratorio de mamíferos del Instituto de Ciencia Naturales de la Universidad Nacional de Colombia. Hugo López-Arevalo (ICN), Yaneth del Socorro Muñoz-Saba (ICN), Pilar Rivas Pava (MHNUC), Oscar Murillo (UV) y Manuel Ruedi (MHNG), permitieron la consulta de los especímenes en las colecciones a su cargo. Louise Tomsett (BMNH) facilitó imágenes de alta resolución del Holotipo de Histiotus colombiae. Paul Velazco, Martha Rocha, Alberto Cadena, Marcela Gómez-Laverde y Jorge Contreras, suministraron literatura y equipos indispensables para el desarrollo del trabajo. La ciudad de Ginebra (Suiza), financió la visita del autor al MHNG donde examinó material crítico para este trabajo. Algunas conclusiones aquí expuestas surgieron de discusiones casuales y muy interesantes con Hugo Mantilla-Meluk y Héctor Ramírez. Ángela Camargo Sanabria, Danny Zulay Urrego, Ángela Patricia Cuervo y Carolina Castellanos aportaron valiosos comentarios editoriales al manuscrito. A las familias que prestaron su hospitalidad en la fase campo, en especial a don Airan Arango y su familia. Finalmente, un agradecimiento especial a Martha María Carmona, sin su excelente compañía y ánimo no hubiera sido tan exitoso el trabajo en campo.
LITERATURA CITADA
1. ALBERICO, M., A. CADENA, J. HERNANDEZ-CAMACHO & Y. MUÑOZ-SABA. 2000. Mamíferos (Synapsida: Theria) de Colombia. Biota Colombiana 1:43-75. [ Links ]
2. ALFONSO-B, A. & A. CADENA. 1994. Composición y estructura trófica de la comunidad de murciélagos del Parque Regional Natural Ucumarí. Págs.361-373 en: J. O. Rangel (ed.), Ucumarí: Un caso típico de la diversidad biótica Andina. Carder, Universidad Nacional de Colombia. Pereira. [ Links ]
3. ARMENTERAS, D., F. GAST & H. VILLAREAL. 2003. Andean forest fragmentation and the representativeness of protected areas in the eastern Andes, Colombia. Biological Conservation 113:245-156. [ Links ]
4. BARQUEZ, R.M., M.A. MARES & J.K. BRAUN. 1999. The bats of Argentina. Special Publications, Museum of Texas Tech University 42:1-275. [ Links ]
5. BEJARANO-BONILLA, D.A., A. YATE-RIVAS & M.H. BERNAL-BAUTISTA. 2007. Diversidad y distribución de la fauna quiróptera en un transecto altitudinal en el departamento del Tolima, Colombia. Caldasia 29:297-308. [ Links ]
6. CADENA, A., R.P. ANDERSON & P. RIVAS-PAVA. 1998. Colombian mammals from the chocoan slopes of Nariño. Occasional Papers, Museum of Texas Tech University 108:1-15. [ Links ]
7. CASTAÑO, J.H., Y. MUÑOZ-SABA, J.E. BOTERO & J.H. VÉLEZ. 2003. Mamíferos del departamento de Caldas-Colombia. Biota Colombiana 4:247-259. [ Links ]
8. CASTAÑO, J.C., J.E. BOTERO, S. VELÁSQUEZ & J.D. CORRALES. 2004. Murciélagos en agrosistemas cafeteros de Colombia. Chiroptera Neotropical 10:196-199. [ Links ]
9. CLOUTIER, D. & D.W. THOMAS. 1992. Carollia perspicillata. Mammalian Species 417:1-9. [ Links ]
10. CONTRERAS, M. 1999. Género Sturnira (Chiroptera: Phyllostomidae) de Colombia. Tesis de Maestría, Instituto de Ciencias Naturales, Universidad de Colombia. Bogotá, D.C. [ Links ]
11. CUARTAS-CALLE, C.A. & J.A. MUÑOZ-ARANGO. 2003. Lista de los mamíferos (Mammalia: Theria) del departamento de Antioquia, Colombia. Biota Colombiana 4:65-78. [ Links ]
12. CUARTAS, C.A., J.A. MUÑOZ & M. GONZÁLEZ. 2001. Una nueva especie de Carollia Gray, 1838 (Chiroptera: Phyllostomidae) de Colombia. Actualidades Biológicas 23:63-73. [ Links ]
13. DAVIS, W.B. 1980. New Sturnira (Chiroptera: Phyllostomidae) from Central and South America, with key to currently recognized species. Occasional Papers, Museum of Texas Tech University 70:1-5. [ Links ]
14. ESTRADA-VILLEGAS, S., C.A. PEDRAZA, J PÉREZ-TORRES & P. STEVENSON. 2007. Nuevo reporte sobre la dieta de Sturnira aratathomasi y algunas notas sobre su distribución en Colombia. Chiroptera Neotropical 13:298-304. [ Links ]
15. GARDNER, A.L. 2007. Tribe Sturnirini. Págs.363-376 en. A. L. Gardner (ed.), Mammals of South America, Volume 1, Marsupials, Xenarthrans, Shrews and Bats. The University of Chicago Press. Chicago. [ Links ]
16. GRAHAM, G.L. 1990. Bats versus Birds: comparisons among Peruvian volant vertebrate faunas along an elevational gradient. Journal of Biogeography 17:657-668. [ Links ]
17. GIANNINI, N. & R. BARQUEZ. 2003. Sturnira erythromos. Mammalian Species 729:1-5. [ Links ]
18. HANDLEY, C.O. Jr. 1996. New species of mammals northern South America: Bats of the genera Histiotus Gervais and Lasiurus Gray (Chiroptera: Vespertilionidae). Proceedings of the Biological Society of Washington 109:1-9. [ Links ]
19. HERNÁDEZ-MESA, B., Y. DOMÍNGUEZ-CASTELLANOS & J. ORTEGA. 2005. Myotis keaysi. Mammalian Species 785:1-3. [ Links ]
20. HUTSON, A.M., S.P. MICKLEBURGH & P.A. RACEY (comp.). 2001. Microchiropteran bats: global status survey and conservation action plan. IUCN/SSC Chiroptera Specialist Group. IUCN, Gland & Cambridge. [ Links ]
21. IUDICA, C. 2000. Systematic revision of the Neotropical fruit bats of the genus Sturnira: a molecular and morphological approach. Tesis de Doctor de Filosofía, Universidad de Florida, Gainesville. [ Links ]
22. KALKO, E.K.V. 1998. Organisation and diversity of tropical bat communities through space and time. Zoology 101:281-297. [ Links ]
23. KATTAN, G.H., P. FRANCO, V. ROJAS & G. MORALES. 2004. Biological diversification in a complex region: A spatial analysis of faunistic diversity and biogeography of the Andes of Colombia. Journal of Biogeography 31:1829-1839. [ Links ]
24. KOOPMAN, K.F. 1994. Chiroptera: Systematics. Handbook of zoology volume 8 pt 60, Mammalia:1-217. [ Links ]
25. KUNZ, T.H. & A. KURTA. 1988. Capture methods and holding devices. Págs.1-28 en: Kunz T. H. (ed.), Ecological and behavioral methods for the study of bats. Smithsonian Institution Press. Washington D. C. [ Links ]
26. JARRÍN-V., P. 2005. Forma, tamaño y estructura: Aspectos ecológicos, consecuencias evolutivas y ejemplos en los murciélagos ecuatorianos. Revista de la Pontificia Universidad Católica del Ecuador 76:125-146. [ Links ]
27. LAVAL, R.K. 1973. A revision of the neotropical bats of the genus Myotis. Contributions in Science, Los Angeles County Museum 15:1-52. [ Links ]
28. LINARES, O.J. 1987. Murciélagos de Venezuela. Cuadernos Lagoven. Caracas. [ Links ]
29. LÓPEZ-GONZÁLEZ, C., S.J. PRESLEY, R.D. OWEN & M.R WILLIG. 2001. Taxonomic status of Myotis (Chiroptera: Vespertilionidae) in Paraguay. Journal of Mammalogy 82:138-160. [ Links ]
30. MANTILLA-MELUK, H. & R.J. BAKER. 2006. Systematic of Small Anoura (Chiroptera: Phyllostomidae) from Colombia, with Description of a New Species. Occasional Papers, Museum of Texas Tech University 261:1-17. [ Links ]
31. MANTILLA-MELUK, H. & R. J. BAKER. 2008. Mammalia, Chiroptera, Phyllostomidae, Anoura fistulata: Distribution extension. Check List 4:427-430. [ Links ]
32. MOLINARI, J. 1994. A new species of Anoura (Mammalia Chiroptera Phyllostomidae) from the Andes of northern South America. Tropical Zoology 7:73-86. [ Links ]
33. MUÑOZ, J. 1990. Diversidad y hábitos alimenticios de murciélagos en transectos altitudinales a través de la Cordillera Central de los Andes en Colombia. Studies on Neotropical Fauna and Environment 25:1-17. [ Links ]
34. MUÑOZ, J. 2001. Los Murciélagos de Colombia. Sistemática, distribución, descripción, historia natural y ecología. Editorial Universidad de Antioquia, Medellín. [ Links ]
35. MUÑOZ, J., C.A. CUARTAS-CALLE & M. GONZÁLES. 2004. Se describe una nueva especie de murciélago del género Carollia Gray, 1838 (Chiroptera: Phyllostomidae) de Colombia. Actualidades Biológicas 26:80-90. [ Links ]
36. MYERS, N., R.A. MITTERMEIER, CG. MITTERMEIER, G.A.B. DA FONSECA & J. KENT. 2000. Biodiversity hotspots for conservation priorities. Nature 403:853-858. [ Links ]
37. NUMA, C., J.R. VERDÚ & P. SÁNCHEZ-PALOMINO. 2005. Phyllostomid bat diversity in a variegated coffee landscape. Biological Conservation 122:151-158. [ Links ]
38. PACHECO, V., S. SOLARI & P. VELAZCO. 2004. A new species of Carollia (Chiroptera: Phyllostomidae) from the Andes of Peru and Bolivia. Occasional Papers, Museum of Texas Tech University 236:1-15. [ Links ]
39. PACHECO, V. & B.D. PATTERSON. 1991. Phylogenetic relationships of the new world bat genus Sturnira (Chiroptera: Phyllostomidae). Bulletin of the American Museum of Natural History 206:101-121. [ Links ]
40. PATTERSON, B. D., D.F. STOTZ, S. SOLARI, J.W. FITZPATRICK & V. PACHECO. 1998. Contrasting patterns of elevational zonation for birds and mammals in the Andes of southeastern Peru. Journal of Biogeography 24:593-607. [ Links ]
41. PETERSON, R. L. & J. R. TAMSITT. 1968. A new species of bat of the genus Sturnira (Family Phyllostomatidae) from northwestern South America. Life Sciences Occasional Papers, Royal Ontario Museum 12:1-8. [ Links ]
42. PINE, R.H. 1972. The bats oh the genus Carollia. The Texas Agricultural Experiment Station Technical Monograph 8:1-125. [ Links ]
43. RIVAS-PAVA, P., H.E. RAMÍREZ, Z.I. ÁLVAREZ & B.L. NIÑO-VALENCIA. 2007. Catálogo de los mamíferos presentes en las colecciones de referencia y exhibición del Museo de Historia Natural de la Universidad del Cauca. Taller Editorial, Universidad del Cauca, Popayán. [ Links ]
44. RODRÍGUEZ-POSADA, M.E. & P. SÁNCHEZ-PALOMINO. 2009. Taxonomía del género Phyllostomus (Chiroptera: Phyllostomidae) en Colombia. Mastozoología Neotropical 16:153-168. [ Links ]
45. SÁNCHEZ, F., P. SÁNCHEZ-PALOMINO, & A. CADENA.2004. Inventario de mamíferos en un bosque de los Andes Centrales de Colombia. Caldasia 26:291-309. [ Links ]
46. SÁNCHEZ, F., P. SÁNCHEZ-PALOMINO, & A. CADENA. 2008. Species richness and indices of abundance of medium-sized mammals in Andean forest and reforestations with Andean leader: A preliminary analysis. Caldasia 30:197-208. [ Links ]
47. SÁNCHEZ-HERNÁNDEZ, C., M.L. ROMERO-ALAMARAZ & G.D. SCHNELL. 2005. A new species of Sturnira (Chiroptera: Phyllostomidae) from northern South America. Journal of Mammalogy 86:866-872. [ Links ]
48. SIMMONS, N.B. 2005. Order Chiroptera. Págs.312-529 en: D. E. Wilson & D. M. Reeder (eds.), Mammal species of the world: a Taxonomic and geographic reference. Smithsonian Institution Press. Washington D.C. [ Links ]
49. SIMMONS, N. & R.S. VOSS. 1998. The mammals of Paracou, French Guiana: A Neotropical lowland rainforest fauna. Part 1. Bats. Bulletin of the American Museum of Natural History 237:1-219. [ Links ]
50. SOLARI, S. & R.J. BAKER. 2006. Mitochondrial DNA sequence, karyotypic, and morphological variation in the Carollia castanea species complex (Chiroptera: Phyllostomidae) with description of a new species. Occasional Papers, Museum of Texas Tech University 254:1-16. [ Links ]
51. SORIANO, P. . & J. MOLINARI. 1987. Sturnira aratathomasi. Mammalian Species 284:1-4. [ Links ]
52. STILES, F.G. & L. ROSELLI. 1998. Inventario de las aves de un bosque altoandino: comparación de dos métodos. Caldasia 20:29-43. [ Links ]
53. TAMSITT, J. R. & D. NAGORSEN. 1982. Anoura cultrata. Mammalian Species 179:1-5. [ Links ]
54. TAMSITT, J. R. & D. VALDIVIESO. 1966. Taxonomic comments on Anoura caudifer, Artibeus lituratus and Molossus molossus. Journal of Mammalogy 47:230-238. [ Links ]
55. THOMAS, O. 1916. Notes on bats of the genus Histiotus. Annals and Magazine of Natural History series 8, 17:272-276. [ Links ]
56. TIRIRA, D. 1998. Memorias del Seminario - Taller: Biología, sistemática y conservación de los Mamíferos del Ecuador. Museo de Zoología. Centro de Biodiversidad y Ambiente. Pontificia Universidad Católica del Ecuador. Publicación especial I. Quito. [ Links ]
57. VOSS, R.S. & L.H. EMMONS. 1996. Mammalian diversity in Neotropical Lowland Rainforest: a preliminary assessment. Bulletin of the American Museum of Natural History 230:1-115. [ Links ]
58. WILSON, D.E., F.R. COLE, J.D. NICHOLS, R. RUDRAN & M.S. FOSTEr (eds.). 1996. Measuring and monitoring biological diversity: Standard methods for mammals. Smithsonian Institution Press, Washington D.C. [ Links ]
59. WILSON, D.E. & R.K. LAVAL. 1974. Myotis nigricans. Mammalian Species 39:1-3. [ Links ]

