










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkCaldasia
versión impresa ISSN 0366-5232versión On-line ISSN 2357-3759
Caldasia v.32 n.2 Bogotá jul./dic. 2010
El impacto del huracán Beta en los bosques de la isla Providencia, Colombia, Suroccidente caribeño
JORGE RUIZ
MARÍA C. FANDIÑO
Escuela de Ciencias Sociales; Convenio Uptc-IGAC. Universidad Pedagógica y Tecnológica de Colombia (Uptc). Bogotá, D.C. Colombia. jorge.ruiz@uptc.edu.co
Patrimonio Natural Fondo para la Biodiversidad y Áreas Protegidas, Programa Incentivos a la Conservación. Bogotá, D.C. Colombia. mfandino@patrimonionatural.org.co
ABSTRACT
One of the consequences of global warming in the Caribbean is an increase in the frequency and intensity of hurricanes. Little is known on the impact of this natural phenomenon on forests, particularly for dry tropical forests. Understanding this impact in terms of structure and species richness is important for forest management. Slow moving Hurricane Beta, a category 1, struck Old Providence island, Colombia, in October 29, 2005. Before Beta woody vegetation was characterized by 88 2 x 50 m plots (0.01 ha) established throughout the island following the protocol by Gentry (1982); 59 plots were studied five years earlier and 29 plots four to five months earlier. The impact of hurricane Beta was assessed within 11 plots located in the Dry Tropical Forests of Old Providence, six months after the hurricane. The These plots were measured in species composition, diameter at breast height (DBH), and height were measured within these plots. There was a considerable reduction in the number of individuals, stems, height, basal areas, and there was no significant difference between DBH. Height damage was positively associated with increasing DHB class. Furthermore, based on the results of species richness, even after controlling for the different number of individuals, through rarefaction, there was no major difference before and after Beta.
Key words. Caribbean, Colombia, dry tropical forest, hurricanes, Old Providence.
RESUMEN
Una de las consecuencias del calentamiento global en el Caribe es el aumento de la frecuencia e intensidad de los huracanes. Poco se sabe del impacto de este fenómeno natural en los bosques, particularmente para el bosque seco tropical (BsT). Entender este impacto en términos de estructura y riqueza de especies es importante para la gestión del bosque. El huracán Beta, categoría 1 y con lento movimiento, azota a Providencia el 29 de octubre de 2005. Antes de Beta, se contaba con una caracterización de la vegetación leñosa en 88 cuadrantes de 2 x 50 m (0.01 ha) establecidos por toda la isla siguiendo el protocolo de Gentry (1982); 59 cuadrantes estudiados cinco años antes y 29 cuadrantes cuatro o cinco meses antes del paso de Beta. El impacto del huracán es medido en once cuadrantes seis meses más tarde del paso del huracán. Estos cuadrantes se censaron en composición de especies, diámetro a la altura del pecho (DAP) y altura. El estudio revela una reducción considerable en el número de individuos, de fustes, área basal y altura; sin diferencia significativa en DAP. El daño en la altura se encuentra positivamente asociado al incremento en la clase de DAP. Adicionalmente, con base en los resultados de riqueza de especies, aun controlando por el distinto número de individuos, a través de la rarefacción, no hay una diferencia mayor antes y después de Beta.
Palabras clave. Bosque seco tropical, Caribe, Colombia, huracanes, Providencia.
Recibido: 08/06/2009
Aceptado: 01/11/2010
INTRODUCTION
Neotropical woody flora is being lost before it is understood. The Gentry (1982) blue print is one of the most commonly used protocols in the study of neotropical forests (Phillips & Raven, 1996) and researchers usually aim for 10 2 x 50 m (0.01 ha) of these plots for a total area of 0.1 ha. There are great advantages of this protocol; Phillips et al. (2002) claim its efficiency for floristic assessment in neotropical forests. Furthermore, given scarce resources, no wonder, the total number of 0.1-ha inventories exceeds 650 (Phillips & Miller, 2002). This database is of great research value, particularly with the current scenario of rapid global climate change, if one is able to resample.
One of the consequences of global warming in the Caribbean is an increase in the frequency and intensity of hurricanes (Emanuel 2005, Tompkins 2005, Knutson & Tuleya 2004 ). Little is known on the impact of this natural phenomenon on forests, particularly for Dry Tropical Forests (DtF). Dry tropical forests of the Caribbean islands are among the most threatened tropical ecosystems both as a result of human and natural disturbances (Imbert & Potercup 2008). While hurricanes are an important natural disturbance (Boose et al. 1994), nevertheless, it is difficult to assess hurricane damage on forests due to the lack of previous ecological data, as well as differences and timing of the studies (Tanner et al. 1991). Lugo (2000) argues that the hurricanes' impact on forests are 1) sudden and massive tree mortality 2) delayed patterns of tree mortality, 3) alternative methods of forest regeneration, 4) opportunities for a change in successional direction, 5) high species turnover and opportunities for species change in forests; diversity of age classes 6) faster biomass and nutrient turnover 7) species substitutions and changes in turnover time of biomass and nutrients, 8) lower aboveground biomass in mature vegetation, 9) carbon sinks 10) selective pressure on organisms, 11) and convergence of community structure and organization. The purpose of this paper is to assess the impact of hurricane Beta on tree mortality, sprouting ability, above ground biomass and species richness.
2005 was the year with the highest upper sea-surface temperature on record, and was unprecedented in hurricane activity (Shein, 2006). For example Katrina, Rita and Wilma, were all three category five hurricanes. This hurricane season will be remembered for breaking records in the Atlantic hurricane activity that completely exhausted the alphabet, thus, subsequent storms are named by the National Hurricane Center (NHC) following the Greek alphabet. Caviedes (1991) argues that hurricanes in the Caribbean are more common in El Niño years, yet based on Oceanic Niño Index (ONI) 2005 was not one of them.
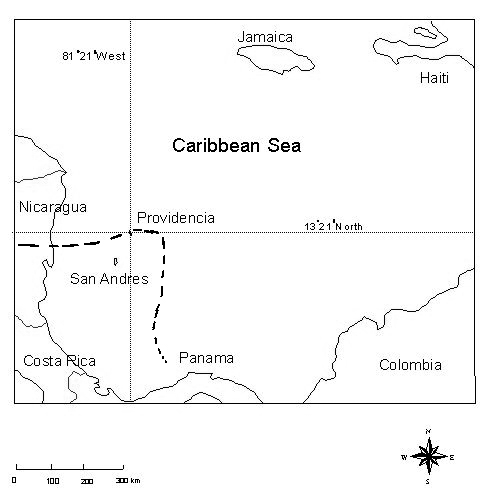
Slow moving hurricane Beta at 6 km/hr, a category one hurricane on the Saffir-Simpson Hurricane Scale, struck Old Providence Island (Providencia), on October 29, 2005 (National Hurricane Center, 2005). This was the thirteenth and final major hurricane of the season. Even if the category of this hurricane was not high, it is worth underlying that the island was battered for hours with 140 km/hr winds. When it struck the Miskito coast of Nicaragua, it had escalated to a category three hurricane (Figure 1). Most of the damage occurred to the west and south of the island. Hurricane Beta followed a path with noticeably less hurricane frequency than its northerly counterparts. In fact, the island was last struck by a hurricane, Hattie, 44 years before Beta, on October, 1961.
According to some authors, taller and larger trees are more severely damaged by hurricanes that their smaller counterparts (Oswalt & Oswalt 2008, Van Bloem et al. 2005, 2006, Reilly 1991, Dittus, 1985). However, after Katrina, a category five hurricane, Chapman and colleagues (2008) found that damage did not vary according to tree size. In the present study it was hypothesized that the hurricane had a significant effect on species richness, basal area, tree density, number of stems and tree height before and after the hurricane.
SITE DESCRIPTION
Old Providence, or Providencia, is a very small and rugged Colombian island, 22 km2 in area, about 7.5 x 4.5 km at its maximum dimensions, located in the southwestern Caribbean (13?21´North and 81?21´West, Figure 1). The island is situated 230 km east of Nicaragua and 714 km northwest of Cartagena, Colombia. The island is an extinct volcano of andesite and basalt (Dunn & Saxe 1950, Proctor 1950, Kerr 1978) and some coral limestone (Parsons 1956), with the highest summit reaching over 365 meters. Today the archipelago is comprised of Providencia and Santa Catalina, the latter is 1 km2 and are both united by a bridge, 190 m in length.
Alwyn H. Gentry from the Missouri Botanical Garden visited the island in 1993 and collected some specimens deposited at the Missouri Botanical Garden Herbarium (MO). In Gonzalez et al. (1995) prologue, citing Gentry´s field notes, the vegetation of Providencia is described as a subset of those species shared by the Yucatan Peninsula and the Caribbean, but with greater affinity with the latter. The woody vegetation of the DtF of Providencia is in good condition since the relative abundance curve follows a log-normal model, suggesting this forest hosts a large, mature and varied community (Ruiz & Fandiño 2009). The archipelago hosts the Old Providence and McBean Lagoon National Park, mostly in a shallow maritime zone. UNESCO declared the archipelago, together with San Andrés, a Man and the Biosphere reserve in November 2000.
The island contains, based on a 2005 QuickBird satellite image, approximately 46 ha of mangroves, and approximately 1,100 ha of forests. Overall, ca. 50% of the island is forested. The dominant ecosystem is dry tropical forest.
The simplest definition of what constitutes a dry tropical forest refers to forests located in the tropical regions that are affected by several months of draught, including moths with absolute draught (Mooney et al. 1996). According to the Colombian Meteorological Institute`s (IDEAM) meteorological station located at the airport, for 1973-2006, the mean annual precipitation for the archipelago was 1,681 mm and the mean annual temperature was 27.6º C, both show a historical positive trend. The archipelago experiences an annual dry period of five months, January through May, four of which precipitation is below 100 mm while 90% of the precipitation occurs between July and December.
MATERIALS AND METHODS
In April, 2006, eleven 2 x 50 m (0.01 ha) plots were established, following the Gentry (1982) protocol, as close to the original site as possible, nine originally established between December 2000 and January 2001, and two in August, 2005. The plots chosen were located in the west and south of the island, which was most severely impacted by the hurricane and for secondary forest 17 years and older, including old growth forest. Young forests, i.e. under 17 years of age were not chosen. Therefore it was hypothesized that younger trees were less likely to be damaged.
Previous to the hurricane, for each of the 11 plots, differentially corrected GPS coordinates were recorded at the beginning and end of 50 m tape. After Beta, the aim was to go back as close as possible where the initial plot was measured. For two plots assessed five years before the hurricane, all trees >2.5 cm DBH, were individually tagged before the hurricane thus permitting its exact relocation; the other nine plots are not at the exact same location but at the same property, given a navigation error and the fact that in Old Providence most owners have micro properties ca. 1 ha. Navigation to the selected plots was done using as 12 channel GPS receiver (MAGELLAN), initially using a map with a plastic cover in the Universal Transverse Mercator (UTM) coordinate system and the World Geodetic Survey 1984 datum (WGS 84), but once close to the plot, the "Inquire cursor" tool in ERDAS IMAGINE 8.7 on a laptop computer was used to ease navigation. The map, GPS receiver and software used the same datum and coordinate system. Once at the original site, an eight channel GPS receiver was located at both ends of the tape, with 15 second logging intervals. While the rover receiver was running, a base station was taking readings at the northwest of the island with identical logging interval in order to perform once more differential correction of the GPS readings and thus estimate the accuracy of the site sampled after Beta.
Following Gentry (1982) each plot was a rectangle measuring 2 x 50 m, set by laying a 50 meter fiberglass tape on the ground. All live woody vegetation >2.5 cm diameter at breast height (DBH) at 1.37 meters (Four feet) was censused. Following Condit et al. (1996) and Gentry (1982) some trees such as cotton tree (Ceiba pentandra, Bombacaceae), were measured at the nearest lower point where the stem was cylindrical, or above the buttress on large trunks. Following Gentry (1982) if a plant overlapped the quadrant boundary, the location of its diameter midpoint determined whether or not it was included. Once the two points of a plot were established, all woody vegetation that was alive, 1 m to each side of the 50 m tape and if the tree or stem had >2.54 cm or one inch DBH were included. For each tree, DHB, height and common name was recorded. For marginal trees in DBH, an aluminum piece with a 2.54 diameter hole was used. If the stem fitted in the hole, the stem was not included. DBH was measured using a LUFKIN Diameter tape that provided diameter when circumference was measured. The same procedure was applied for multiple-stemmed trees, i.e., each stem contributed to DBH and height but only once as an individual. Height was calculated using a 4 m pole and if higher estimated visually.
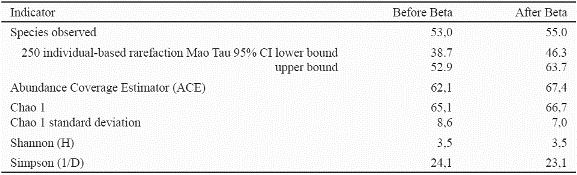
For each tree its common name was recorded. About 30 specimens were collected, with fruit or flower in order to reduce the number of species to be determined based on the unfertile previously collected specimens to be determined. Furthermore, considering that the same species can have more than one common name, all collected samples and common names were compared to the previously collected ones by Ruiz and Fandiño (2006), Morales (2005) and Ruiz and colleagues (2005a), which rest at the FSB Herbarium of the Humboldt Institute of Biological Research in Villa de Leyva, Boyacá, Colombia. More detail of the materials and methods used is available from Ruiz & Fandiño (2009, 2007, 2006) as well as Ruiz et al. (2005 a, b). Data on structure were analyzed in SPSS v. 12.0, specifically the paired T-Test statistic (2-tailed). Species diversity was measured in EstimateS for WINDOWS v. 7.5 (Colwell, 2009), specifically, two nonparametric estimators of species richness the Abundance-based Coverage Estimator (ACE) and Chao1. Chao 1 is based on the number of singletons (species with one individual) and doubletons (species with two individuals), while ACE is based on the number of species found with ten or fewer individuals (Chazdon et al. 1998); the rational is that rare species are more useful when estimating overall richness (Magurran 2004). Furthermore, in EstimateS 7.5. the individual based Mao Tau expected number of species and their 95% confidence intervals were calculated. Furthermore, in the same software, the Shannon (H) index was computed, as a measure that reflects the species richness element of diversity, and the Simpson Index that expresses the degree of dominance (evenness) in the data; the latter is reported as 1/D considering that as D increases, diversity decreases (Magurran, 2004).
RESULTS
The 22 points of the differentially corrected GPS files at each extreme of 50 m tape for the 11 plots after Beta, in comparison to before beta exhibited a mean of 7.36 m, with a minimum of 0.64 m a maximum of 23.3 m and a standard deviation of 6.58. For the eleven plots, 544 stems were found before the hurricane compared to 390 after the hurricane (Table 1). The number of individuals was reduced from 344 to 250, while the mean number of stems per individual dropped from 1.58 to 1.53. Both, the number of individuals as well as the number of stems show a highly statistical significance, in the paired T-Test, before and after the hurricane. There is no significant change in DBH before and after the hurricane after controlling for DBH class, at the 5% probability (Table 1). However, tree basal area was reduced by 16.6 % and this result was highly statistically significant. Moreover, mean stem height between both dates is substantially different, reduced by one-fifth from 6.86 m to 5.40 m before and after the hurricane respectively. This result is significant at the 1% probability level. As expected, the standard deviation in stem height before the hurricane was higher than after the hurricane. When controlling for DBH class, stems >10 cm DHB were most affected in comparison to smaller trees. Thicker trees experienced a 34.6% reduction in height, intermediate thickness trees 5 to = 10 cm DHB suffered a reduction of 11.95% while the >2.5 to < 5 cm DHB 10.9%. Species mostly affected by a reduction in height were canopy species such as Hog plum (Spondias mombin L.) or cotton tree (Ceiba pentandra (L.) Gaertn.).

In spite of the work done augmenting the number of fertile specimens, 21 species remained to be determined; three before Beta and 18 after beta. Nevertheless, this did not affect species richness which are based on morphospecies. There is no major difference in species richness between both dates (Table 2). This is clearly seen in the overlapping 95% confidence intervals of rarefaction at 250 individuals as well as the overlapping confidence intervals of Chao 1. Nevertheless, ACE increased from 62.1 to 67.4. It is concluded that species richness appears not to be altered due to hurricane disturbance.
DISCUSSION
A careful GPS work enabled the establishment of paired plots, as surrogates of permanent ones. The use of differential correction and careful navigation bestows data to be gathered before and after the hurricane not at the exact location, plus or minus seven meters, but definitely the same property. This procedure could be used in other plots that followed the Gentry (1982) protocol were impact assessment on forest structure and composition is needed, particularly in a scenario of rapid climate change. The fact that some of the plots were established four years before the hurricane undoubtedly introduced some noise in this research but not to the extent of compromising the results, considering that dry tropical forests has a slower growth rate than its moister counterparts.
The number of individuals, stems, stem height and basal area declined significantly after the hurricane. The drop in basal area is probably due to the reduction in the number of individuals as well as stems. Height damage increased with DHB class, with older trees showing the greatest damage in comparison to their smaller counterparts. Our results agree with those reported by Van Bloem et al. (2005) and Oswalt & Oswalt (2008) that documented increasing damage with escalating DHB and height and challenge the results by Chapman et al. (2008) that found no differential damage according to tree size. Other studies such as Tovilla & Orijuela (2004) have found greatest damage on intermediate size trees. Likewise the species richness and diversity indicators proved that was no major difference before and after the hurricane. Our results agree with those by Imbert & Portecup (2008), hurricanes appear to affect dry tropical forest structure but not species richness.
Hurricanes have been, are and will continue to be an important natural disturbance in Caribbean forests. While it is sad to see trees more than one hundred years flat on the ground with no evidence of sprouting, other species, while on the ground are alive have exploded in the number of new stems. It is recommended that nothing should be done with respect to the large amount of trees on the trails of the island, and one should permit nature take its course. This fact will undoubtedly prove to be an obstacle to cattle grazing in this very vulnerable island; considering that cattle need to be moved from ranch to ranch for water and grass, particularly in the dry season. In other words, in the long run, the hurricane could mean a positive disturbance for the perpetuation of this forest, indeed Ganzhorn (1995) claims that windfalls and excessive rains are an important factor for sustaining forest biodiversity. Nevertheless, the reduction in stem height has proven a threat to the endemic iguana (Iguana iguana rhinolopha) since it is far easier now for locals to hunt it.
ACKNOWLEDGEMENTS
This study is an official publication of the Caldas Research Group, registered in COLCIENCIAS under COL0021613. The study was possible thanks to the research permit 006 from 2002 and 598 from 2006; for the jurisdiction of CORALINA, Providencia`s Environmental Authority that, thanks to Elizabeth Taylor CORALINA´S CEO, kindly provided logistical support. We thank the late Asmond Duffis as well as Hawthorne Newball and Regino Robinson for sharing their knowledge of woody plants. Financial support was received for the plots previous to the hurricane from COLCIENCIAS, grant number 1243-13-16640, Universidad de La Salle grant number PINV-013-05, from Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, in Bogotá, Colombia as well from the Department of Natural Resources Management and the Research Foundation of the University of Connecticut, Storrs, CT, USA. We are grateful to the Talero family and Juan Manuel Orozco for their hospitality. Finally, we acknowledge the comments and corrections on an earlier draft by Dr. Ángela Parrado-Roselli from Universidad Distrital in Bogotá, Colombia; as well as comments and corrections provided to us from three anonymous reviewers from Caldasia.
LITERATURE CITED [ Links ]
2. CAVIEDES, C.N. 1991. Five Hundred Years of Hurricanes in the Caribbean: Their Relationship with Global Climatic Variabilities. GeoJournal 23:301-310. [ Links ]
3. CHAPMAN, E.L., J.Q. CHAMBERS, K.F. RIBBECK, D.B. BAKER, M.A. TOBLER, H. HONGCHENG ZENG & D.A. WHITE. 2008. Hurricane Katrina impacts on forest trees of Louisianas Pearl River basin. Forest Ecology and Management 256: 883–889. [ Links ]
4. COLWELL, R K. 2009. EstimateS: Statistical estimation of species richness and shared species from samples. Version 7.5 Users Guide and application. (http://purl.oclc.org/estimates) accessed 17 October, 2010. [ Links ]
5. CONDIT, R., S.P. HUBBELL, J.V. LAFRANKIE, R. SUKUMAR, N. MANOKARAN, R.B. FOSTER & P. S. ASHTON. 1996. Species-area and species-individual relationships for tropical trees: a comparison of three 50-ha plots. Journal of Ecology 84:549-562. [ Links ]
6. DITTUS, W.P.J. 1985. The influence of cyclones on the dry evergreen forest of Sri Lanka. Biotropica 17:1-14. [ Links ]
7. DUNN, E.R. & L.H. SAXE. 1950. Results of the Catherwood-Chaplin West Indies expedition, 1948. Part V. Amphibians & reptiles of San Andrés and Providencia. Proceedings of the Academy of Natural Sciences of Philadelphia CII 141-164. [ Links ]
8. EMANUEL, K. 2005. Increasing destructiveness of tropical cyclones over the past 30 years. Nature 436:686–688. [ Links ]
9. GANZHORN, J.U. 1995. Cyclones over Madagascar: fate or fortune? Ambio 24:124-125. [ Links ]
10. GENTRY, A.H. 1982. Patterns of Neotropical plant species diversity. Evolutionary Biology 15:1-84. [ Links ]
11. GONZÁLEZ, F., J.N. DÍAZ & P. LOWY. 1995. An illustrated flora of San Andrés and Providencia. Bogotá: SENA. [ Links ]
12. IMBERT, D. & J. PORTECOP. 2008. Hurricane disturbance and forest resilience: Assessing structural vs. functional changes in a Caribbean dry forest. Forest Ecology and Management 255(8-9): 3494–3501. [ Links ]
13. KERR, J. 1978. The volcanic and tectonic history of La Providencia Island , Colombia. Unpublished Masters Thesis Dissertation. Rutgers University, New Brunswick, NJ. [ Links ]
14. KNUTSON, T.R. & R. E. TULEYA. 2004. Impact of CO2-induced warming on simulated hurricane intensity and precipitation: sensitivity to the choice of climate model and convective parameterization. Journal of Climate 17:3477–3495. [ Links ]
15. LUGO, A.E. 2000. Effects and outcomes of Caribbean hurricanes in a climate change scenario. The Science of the Total Environment 26: 243-251. [ Links ]
16. MAGURRAN, A.E. 2004. Measuring biological diversity. Blackwell Publishing. Malden. [ Links ]
17. MOONEY, H.A., S.H. BULLOCK & E. MEDINA. 1996. Introduction. Pp 1-6. In: S.H. Bullock, H.A. Mooney & E. Medina (eds.) Seasonally dry tropical forests. Cambridge University Press, Cambridge. [ Links ]
18. MORALES, D.A. 2005. Estudio de la flora del bosque seco caribeño de la isla de Providencia (Colombia) en el área del Peak y análisis de su afinidad geográfica. Bogotá, D. C: Pontificia Universidad Javeriana, Facultad de Estudios Ambientales y Rurales, Departamento de Ecología, Unpublished B. Sc. Dissertation. [ Links ]
19. National Hurricane Center. 2005. (http://www.nhc.noaa.gov/archive/2005/pub/al262005.public.011.shtml) accessed April 17, 2006. [ Links ]
20. OSWALT, S. & C.M. OSWALT. 2008. Relationships between common forest metrics and realized impacts of Hurricane Katrina on forest resources in Mississippi. Forest Ecology and Management 255: 1692-1700. [ Links ]
21. PARSONS, J.J. 1956. Sanadres and Providencia, English speaking islands s in the Western Caribbean. University of California Publications in Geography 12:1-84. [ Links ]
22. PHILLIPS, O.L., R. VASQUEZ, P. NUÑEZ, A.L. MONTEAGUDO, M.E. CHUSPE, W. GALIANO, A. PEÑA, M. TIMANA M. YLI-HALLA & S. ROSE. 2003. Efficient plot-based floristic assessment of tropical forests. Journal of Tropical Ecology 19:629-645. [ Links ]
23. PHILLIPS, O.L. & J. MILLER. 2002. Global patterns of forest diversity:the dataset of Alwyn H. Gentry. Monographs in Systematic Botany 89. Missouri Botanical Garden, St. Louis, Missouri. 319 pp. [ Links ]
24. PHILLIPS, O.L. & P.H. RAVEN. 1996. A strategy for sampling Neotropical forests. In A. C. Gibson (Ed.). Neotropical biodiversity and conservation, pp. 141–165. Los Angeles, California: Occassional Publications of the Milred E. Mathias Botanical Garden. [ Links ]
25. PROCTOR, G.R. 1950. Results of the Catherwood-Chaplin West Indies expedition, 1948. Part I. Plants of Cayo Largo (Cuba), San Andrés and Providencia. Proceedings of the Academy of Natural Sciences of Philadelphia CII 27-42. [ Links ]
26. REILLY, A. 1991. The effects of Hurricane Hugo in three tropical forests in the U. S. Virgin Islands. Biotropica 23: 414-419. [ Links ]
27. RUIZ, J. & M.C. FANDIÑO. 2009 Estado del bosque seco tropical e importancia relativa de su flora leñosa, islas de la Vieja Providencia y Santa Catalina, Colombia, Caribe suroccidental. Revista Academia Colombiana de Ciencias Exactas, Físicas y Naturales 33(126): 5-15. [ Links ]
28. RUIZ, J. & M.C. FANDIÑO. 2007. Plantas leñosas del bosque seco tropical de la Isla de Providencia, Colombia, Caribe occidental. Biota Colombiana 8:87-98. [ Links ]
29. RUIZ, J. & M.C. FANDIÑO. 2006. Estructura y estimación no paramétrica de la riqueza total de especies del bosque seco tropical a través de una cronosecuencia teledetectada de más de 60 años en la Isla de Providencia, Colombia. Bogotá: Informe Final Proyecto1243-13-16640 presentado a COLCIENCIAS. Proyecto financiado por COLCIENCIAS, la Universidad de La Salle & Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. [ Links ]
30. RUIZ, J., M.C. FANDIÑO & R.L. CHAZDON. 2005a. Vegetation Structure, Composition, & Species Richness Across a 56-year Chronosequence of Dry Tropical Forest on Providencia Island, Colombia. Biotropica 37: 520-530. [ Links ]
31. RUIZ, J., M.C. FANDIÑO, T.H. MEYER & I. M. ORTEGA. 2005b. Cambio de la cobertura boscosa en Providencia. Revista de Investigación, Universidad de La Salle 5:141-153. [ Links ]
32. SHEIN, K.A. (Ed.), 2006. State of the Climate in 2005. Special supplement to the Bulletin of the American Meteorological Society 87(6): S1-S102. [ Links ]
33. TANNER, E.V.J., V. KAPOS & J.R. HEALEY. 1991. Hurricane effects on forest ecosystems in the Caribbean. Biotropica 23(4):513-521. [ Links ]
34. TOMPKINS, E.L. 2005. Planning for climate change in small isl & s: Insights from national hurricane preparedness in the Cayman Islands. Global Environmental Change 15:139-149. [ Links ]
35. TOVILLA, C. & D. E. ORIJUELA. 2004. Impacto del huracán Rosa sobre los bosques de manglar de la costa norte de Nayarit, México. Madera y Bosques 2004:63-74. [ Links ]
36. VAN BLOEM, S.J., P.G. MURPHY, A.E. LUGO, R. OSTERTAG, M.R. COSTA, I. RUIZ, S. MOLINA & S.M. CANALS. 2005. The influence of hurricane winds on Caribbean dry forest structure & nutrient pools. Biotropica 37:571-583. [ Links ]
37. VAN BLOEM, S.J., A.E. LUGO & P.G. MURPHY. 2006. Structural response of Caribbean dry forests to hurricane winds: a case study from Guánica Forest, Puerto Rico. Journal of Biogeography 33:517-523. [ Links ]

